
Complete Mitochondrial Genomes of Five Racerunners (Lacertidae: Eremias) and Comparison with Other Lacertids: Insights into the Structure and Evolution of the Control Region



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Library Construction and High-Throughput Sequencing
2.3. Sequence Assembly, Annotation, and Analysis
2.4. Phylogenetic Analysis
3. Results
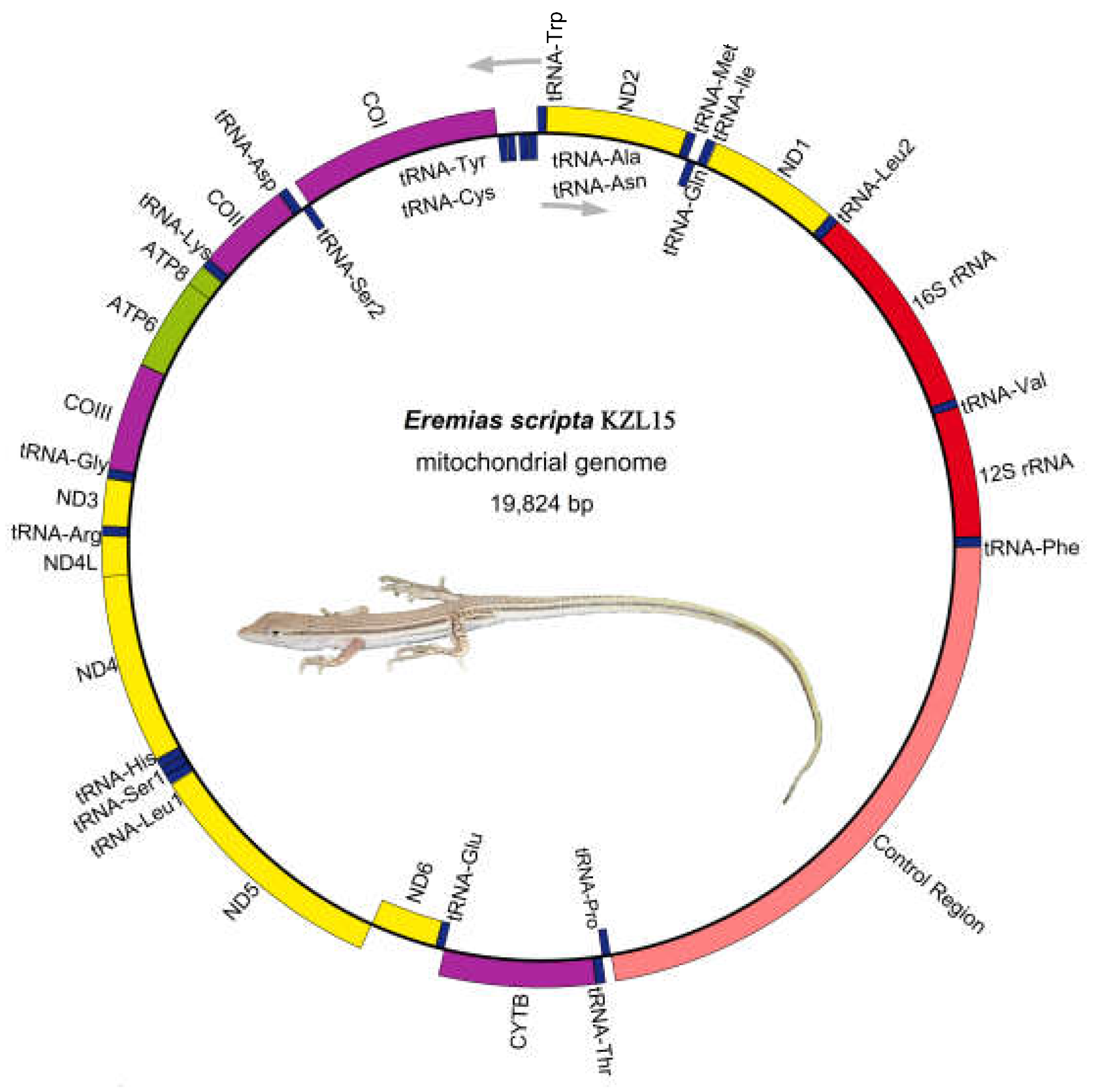
3.1. Genome Organization and Base Composition
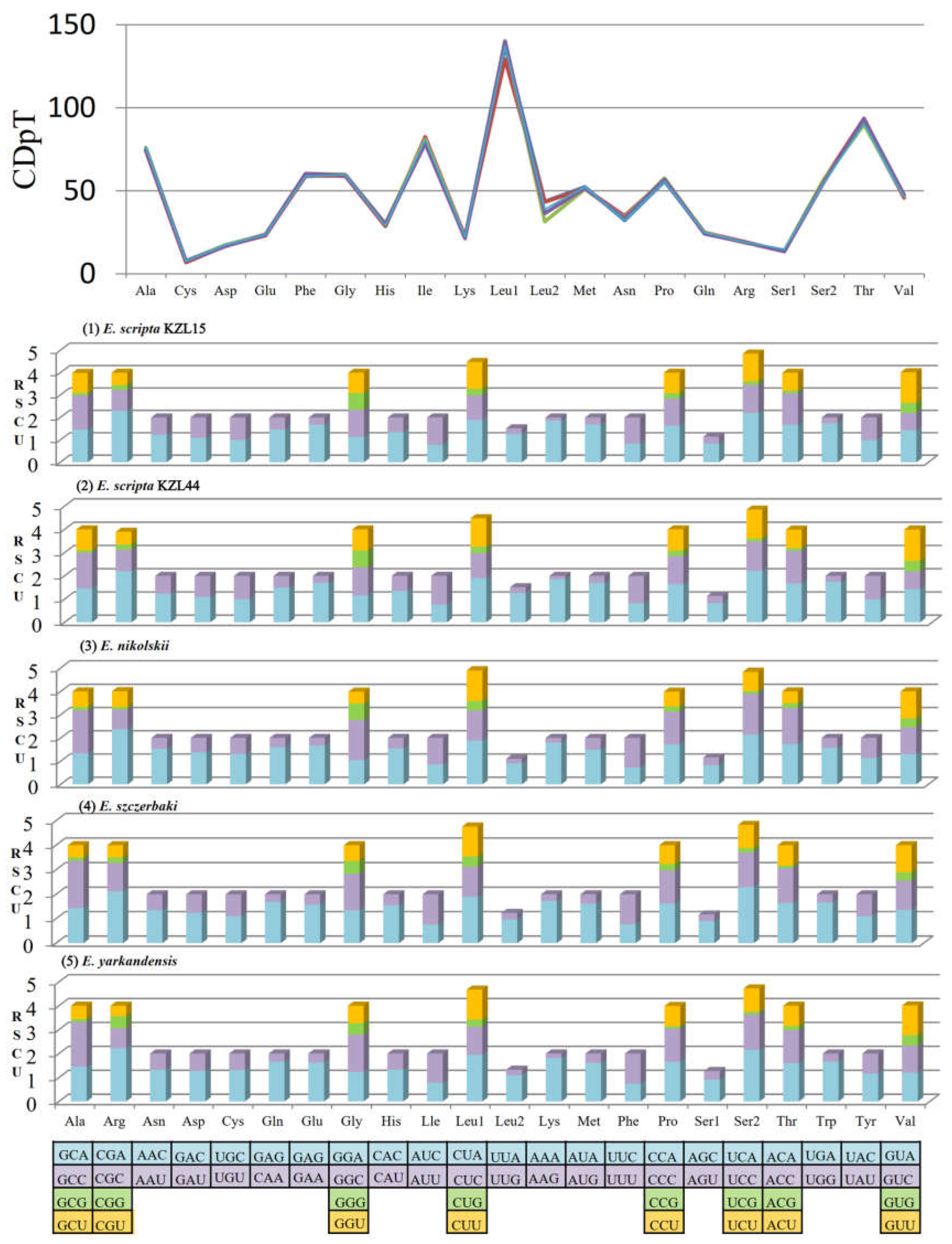
3.2. PCGs and Codon Usage
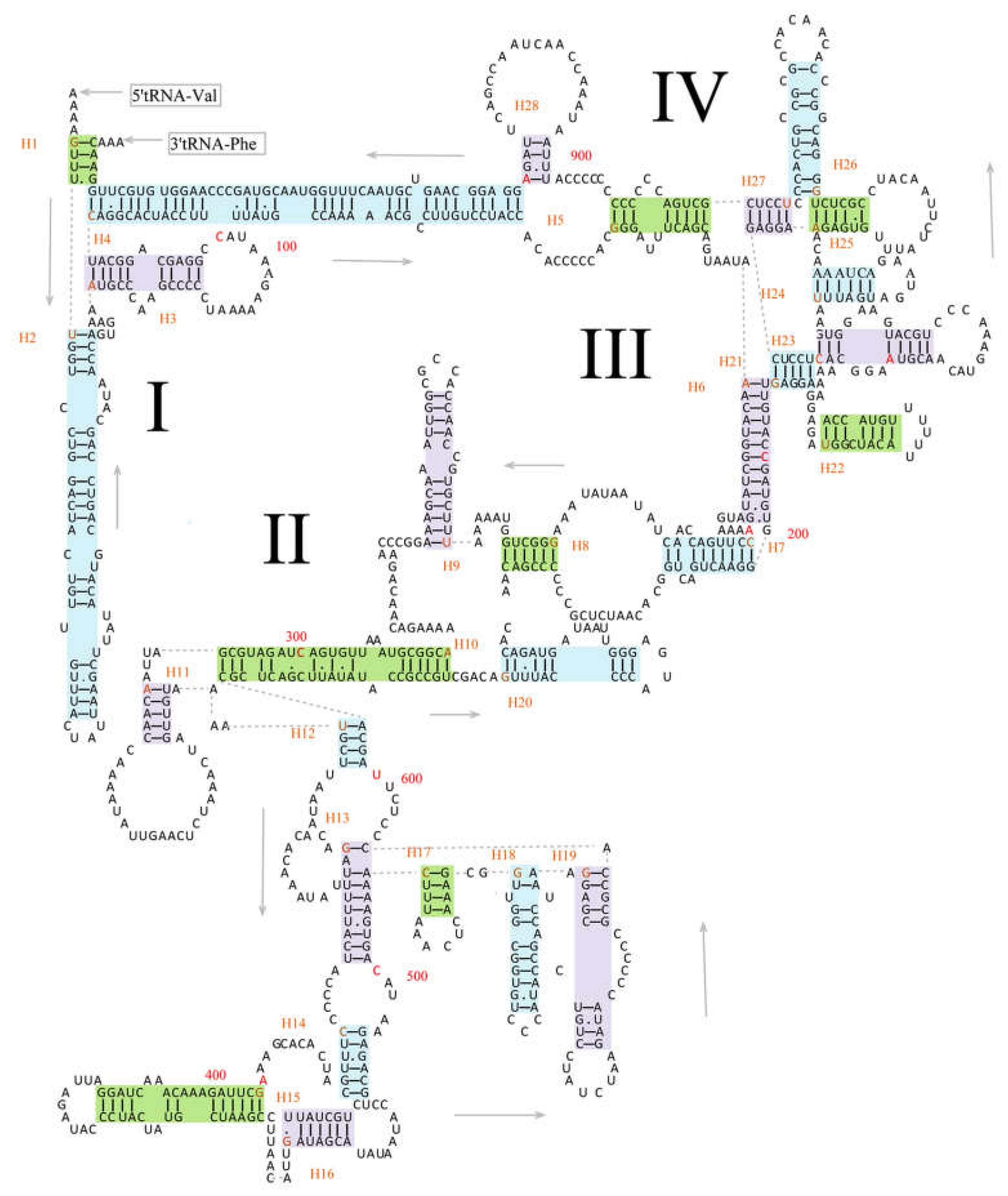
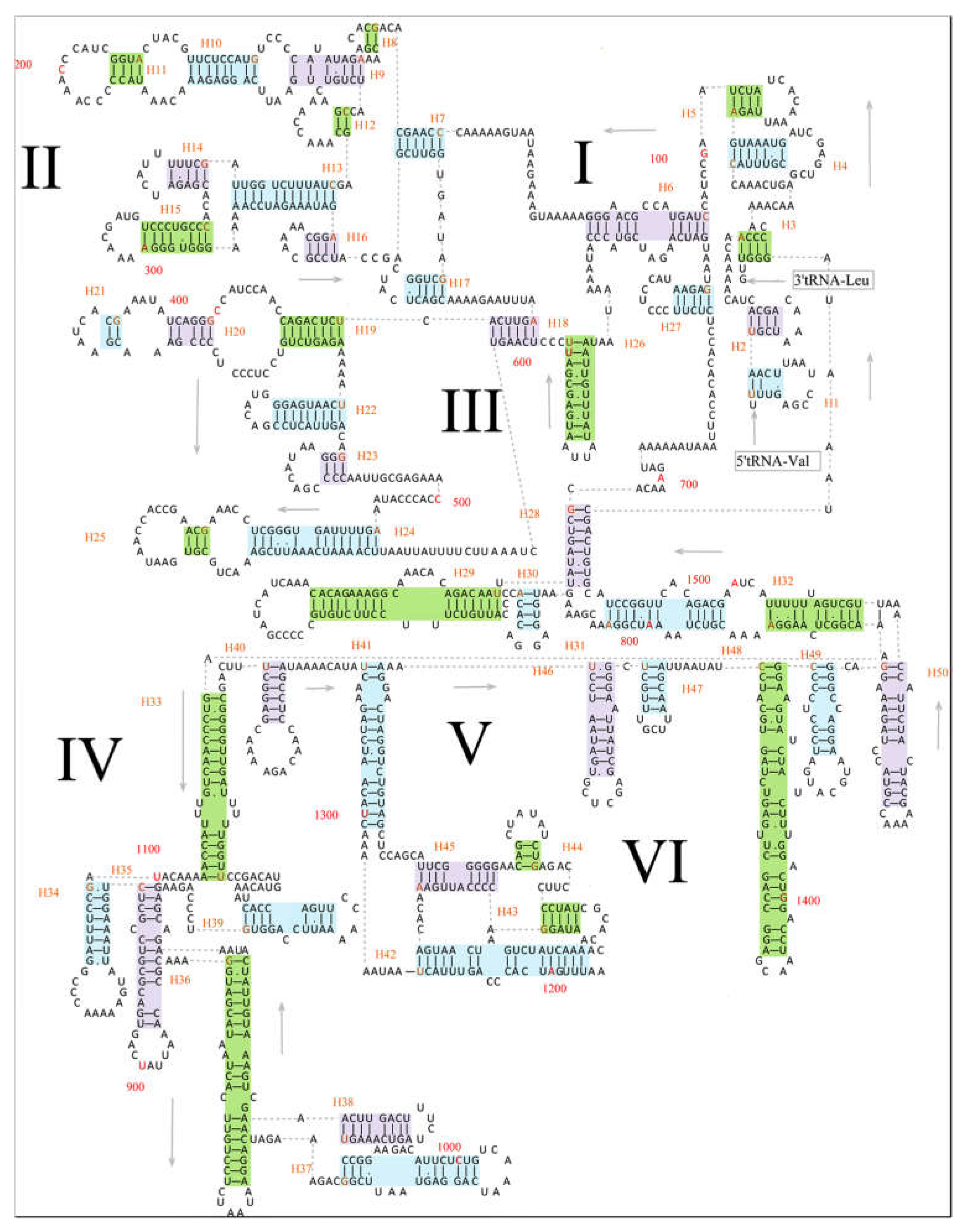
3.3. Transfer RNAs and Ribosomal RNAs
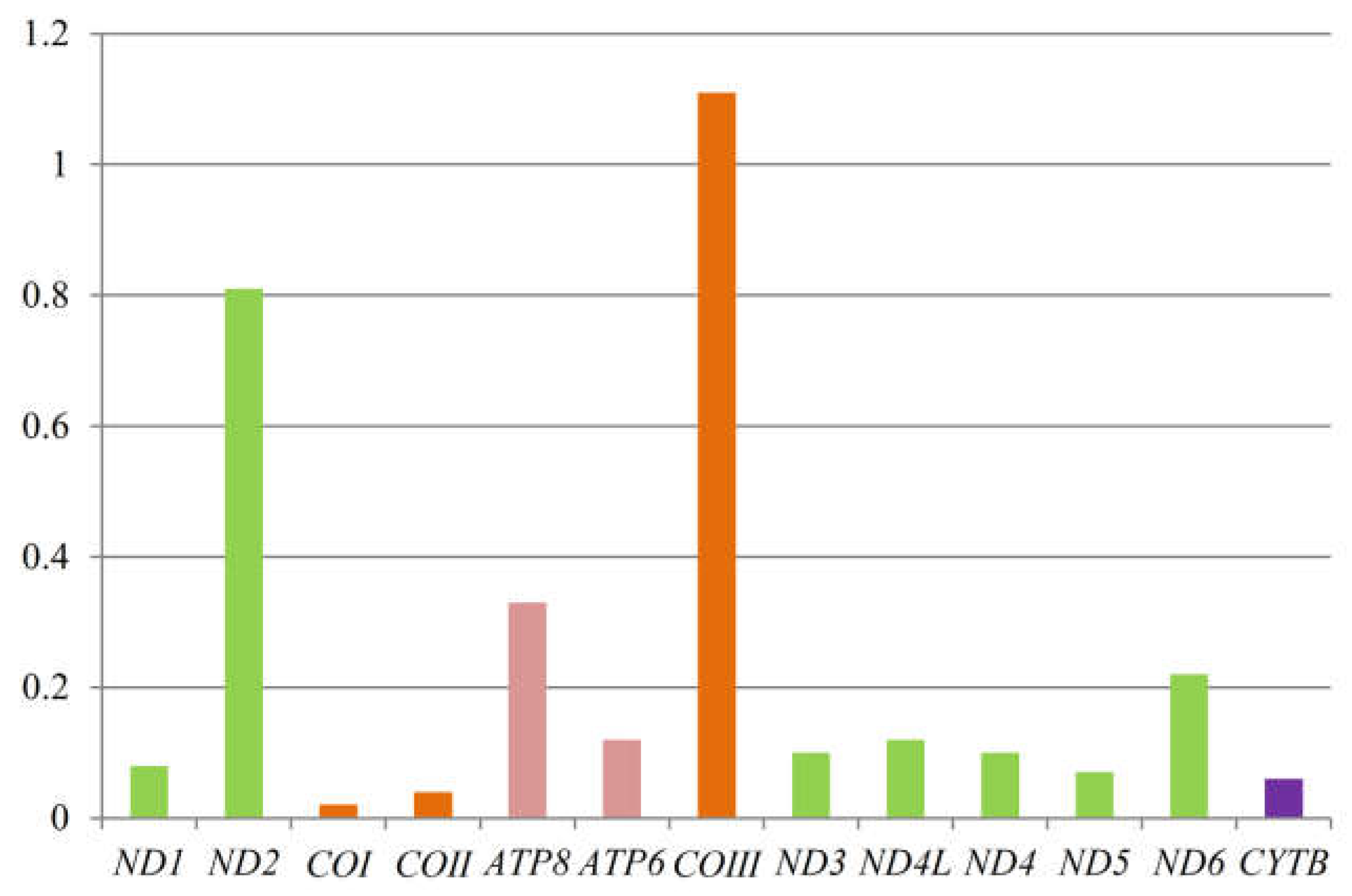
3.4. Non-Synonymous and Synonymous Substitutions
3.5. Structure of Control Region
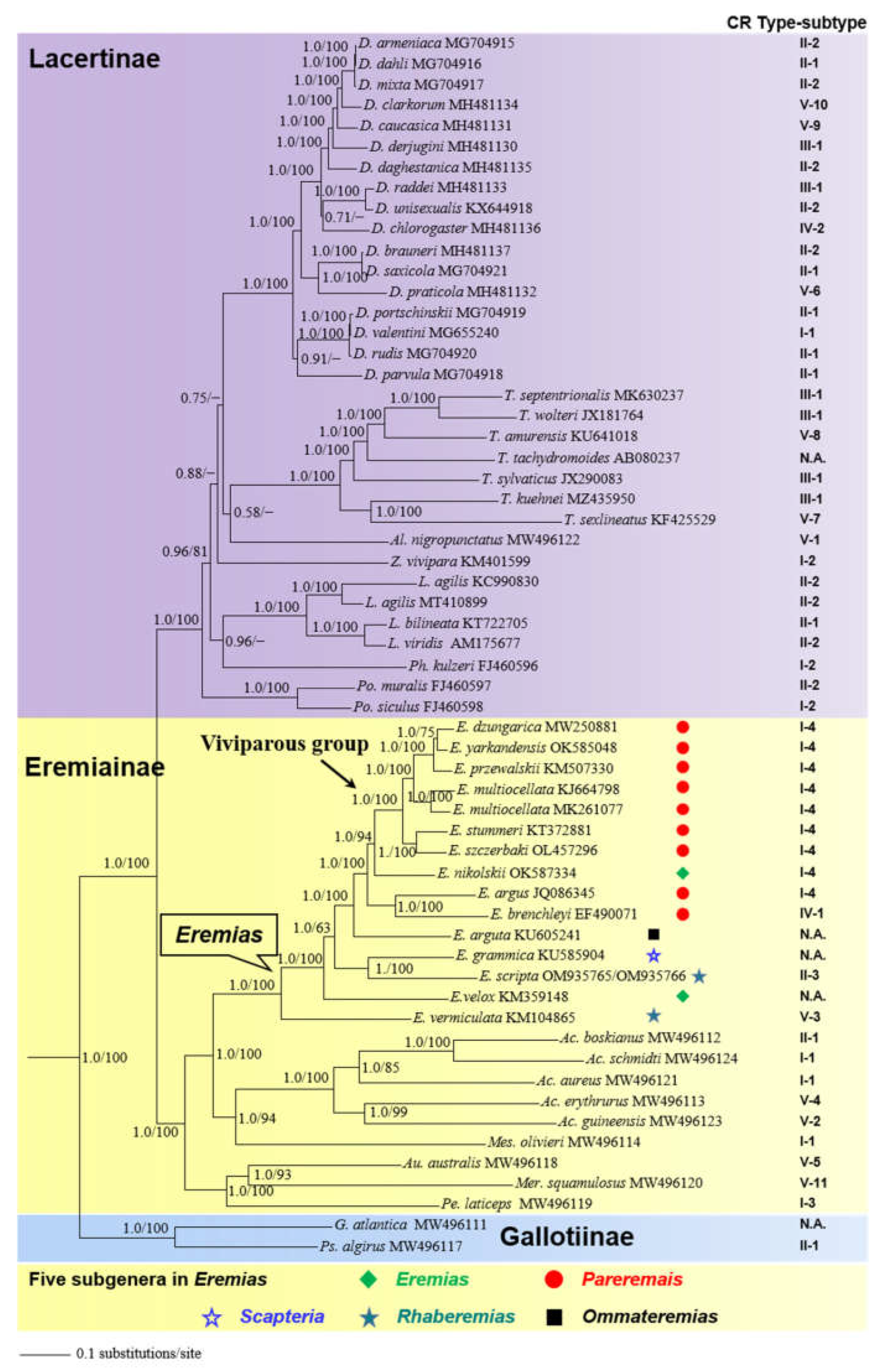
3.6. Phylogenetic Analysis
4. Discussion
4.1. Secondary Structures of rRNA Are Useful for Phylogenetic Inference
4.2. Selection Pressure on PCGs
4.3. Phylogenetic Implications
4.4. Structure and Evolution of Control Region
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.L.; Coulson, A.R.; Drouin, J.I.; Eperon, C.; Nierlich, D.P.; Roe, B.A.; Sanger, F.; et al. Sequence and organization of the human mitochondrial genome. Nature 1981, 290, 457–465. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, D.R. Animal mitochondrial DNA: Structure and evolution. Int. Rev. Cytol. 1992, 141, 173–216. [Google Scholar] [PubMed]
- Brehm, A.; Harris, D.J.; Alves, C.D.; Jesus, J.D.; Thomarat, F.D.; Vicente, L.D. Structure and evolution of the mitochondrial DNA complete control region in the lizard Lacerta dugesii (Lacertidae, Sauria). J. Mol. Evol. 2003, 56, 46–53. [Google Scholar] [CrossRef]
- Hoelzel, A.R.; Lopez, J.V.; Dover, G.A.; O’Brien, S.J. Rapid evolution of a heteroplasmic repetitive sequence in the mitochondrial DNA control region of carnivores. J. Mol. Evol. 1994, 39, 191–199. [Google Scholar]
- Wu, N.; Liu, J.; Wang, S.; Guo, X. Comparative analysis of mitochondrial genomes in two subspecies of the sunwatcher toad-headed agama (Phrynocephalus helioscopus): Prevalent intraspecific gene rearrangements in Phrynocephalus. Genes 2022, 13, 203. [Google Scholar] [CrossRef] [PubMed]
- Sbisà, E.; Tanzariello, F.; Reyes, A.; Pesole, G.; Saccone, C. Mammalian mitochondrial D-loop region structural analysis: Identification of new conserved sequences and their functional and evolutionary implications. Gene 1997, 205, 125–140. [Google Scholar] [CrossRef]
- Harris, D.J.; Arnold, E.N.; Thomas, R.H. Relationships of lacertid lizards (Reptilia: Lacertidae) estimated from mitochondrial DNA sequences and morphology. Proc. Biol. Sci. B 1998, 265, 1939–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, W.; Pavlicev, M. The phylogeny of the family Lacertidae (Reptilia) based on nuclear DNA sequences: Convergent adaptations to arid habitats within the subfamily Eremiainae. Mol. Biol. Evol. 2007, 44, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Pavlicev, M.; Mayer, W. Fast radiation of the subfamily Lacertinae (Reptilia: Lacertidae): History or methodical artefact? Mol. Phylogenet. Evol. 2009, 52, 727–734. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 22 March 2022).
- Arnold, E.N.; Arribas, O.; Carranza, S. Systematics of the Palaearctic and Oriental lizard tribe Lacertini (Squamata: Lacertidae: Lacertinae), with descriptions of eight new genera. Zootaxa 2007, 1430, 1–86. [Google Scholar] [CrossRef]
- Hipsley, C.A.; Himmelmann, L.; Metzler, D.; Müller, J. Integration of Bayesian molecular clock methods and fossil-based soft bounds reveals early Cenozoic origin of African lacertid lizards. BMC Evol. Biol. 2009, 9, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, J.; Harris, D.J.; Carranza, S.; Salvi, D. Evaluating the phylogenetic signal limit from mitogenomes, slow evolving nuclear genes, and the concatenation approach. New insights into the Lacertini radiation using fast evolving nuclear genes and species trees. Mol. Phylogenet. Evol. 2016, 100, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Kirchhof, S.; Lyra, M.L.; Rodríguez, A.; Ineich, I.; Müller, J.; Rödel, M.O.; Trape, J.F.; Vences, M.; Boissinot, M. Mitogenome analyses elucidate the evolutionary relationships of a probable Eocene wet tropics relic in the xerophilic lizard genus Acanthodactylus. Sci. Rep. 2021, 11, 4858. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Dai, X.; Dali, C.; Papenfuss, T.J.; Ananjeva, N.B.; Melnikov, D.A.; Wang, Y. Phylogeny and divergence times of some racerunner lizards (Lacertidae: Eremias) inferred from mitochondrial 16S rRNA gene segments. Mol. Phylogenet. Evol. 2011, 61, 400–412. [Google Scholar] [CrossRef]
- Khan, M.A.; Jablonski, D.; Nadeem, M.S.; Masoor, R.; Kehlmaier, C.; Spitzweg, C.; Fritz, U. Molecular phylogeny of Eremias spp. from Pakistan contributes to a better understanding of the diversity of racerunners. J. Zool. Syst. Evol. Res. 2021, 59, 466–483. [Google Scholar] [CrossRef]
- Liu, J.L.; Dujsebayeva, T.N.; Chirikova, M.A.; Gong, X.; Li, D.J.; Guo, X.G. Does the Dzungarian racerunner (Eremias dzungarica Orlova, Poyarkov, Chirikova, Nazarov, Munkhbaatar, Munkhbayar &Terbish, 2017) occur in China? Species delimitation and identification with DNA barcoding and morphometric analyses. Zool. Res. 2021, 42, 287–293. [Google Scholar]
- Wang, S.; Liu, J.L.; Zhang, B.; Guo, X.G. The complete mitochondrial genome of Eremias dzungarica (Reptilia, Squamata, Lacertidae) from the Junggar Basin in Northwest China. Mitochondrial DNA B 2021, 6, 2012–2014. [Google Scholar] [CrossRef]
- Brown, R.P. Large subunit mitochondrial rRNA secondary structures and site-specific rate variation in two lizard lineages. J. Mol. Evol. 2005, 60, 45–56. [Google Scholar] [CrossRef]
- Böhme, M.U.; Fritzsch, G.; Tippmann, A.; Schlegel, M.; Berendonk, T.U. The complete mitochondrial genome of the green lizard Lacerta viridis viridis (Reptilia: Lacertidae) and its phylogenetic position within squamate reptiles. Gene 2007, 394, 69–77. [Google Scholar] [CrossRef]
- Podnar, M.; Pinsker, W.; Mayer, W. Complete mitochondrial genomes of three lizard species and the systematic position of the Lacertidae (Squamata). J. Zool. Syst. Evol. Res. 2009, 47, 35–41. [Google Scholar] [CrossRef]
- Jauss, R.-T.; Solf, N.; Kolora, S.R.R.; Schaffer, S.; Wolf, R.; Henle, K.; Fritz, U.; Schlegel, M. Mitogenome evolution in the Lacerta viridis complex (Lacertidae, Squamata) reveals phylogeny of diverging clades. Syst. Biodivers. 2021, 19, 682–692. [Google Scholar] [CrossRef]
- Stoneking, M.; Hedgecock, D.; Higuchi, R.G.; Vigilant, L.; Erlich, H.A. Population variation of human mtDNA control region sequences detected by enzymatic amplification and sequence-specific oligonucleotide probes. Am. J. Hum. Genet. 1991, 48, 370–382. [Google Scholar] [PubMed]
- Aquadro, C.F.; Greenberg, B.D. Human mitochondrial DNA variation and evolution: Analysis of nucleotide sequences from seven individuals. Genetics 1983, 103, 287–312. [Google Scholar] [CrossRef]
- Kasamatsu, H.; Robberson, D.L.; Vinograd, J. A novel closed-circular mitochondrial DNA with properties of a replicating intermediate. Proc. Natl. Acad. Sci. USA 1971, 68, 2252–2257. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.J.; Conroy, J.; Howell, W.H.; Kocher, T.D. Structure and evolution of teleost mitochondrial control regions. J. Mol. Evol. 1995, 41, 54–66. [Google Scholar] [CrossRef]
- Randi, E.; Lucchini, V. Organization and evolution of the mitochondrial DNA control region in the avian genus Alectoris. Mol. Evol. 1998, 47, 449–462. [Google Scholar] [CrossRef]
- Liu, H.; Yang, G.; Wei, F.W.; Li, M.; Hu, J.C. Sequence variability of the mitochondrial DNA control region and population genetic structure of sika deers (Cervus nippon) in China. Acta Zool. Sinica 2003, 49, 53–60, (In Chinese with English abstract). [Google Scholar]
- Matson, C.W.; Baker, R.J. DNA sequence variation in the mitochondrial control region of red-backed voles (Clethrionomys). Mol. Biol. Evol. 2001, 18, 1494–1501. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Liu, J.; Chen, D.; Guo, X. Complete mitochondrial genome of a blue-tailed skink Plestiodon capito (Reptilia, Squamata, Scincidae) and comparison with other Scincidae lizards. Genetica 2020, 148, 229–241. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one fastq preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Fastp. Available online: https://github.com/OpenGene/fastp (accessed on 22 March 2022).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SPAdes-v3.10.1. Available online: http://cab.spbu.ru/software/spades/ (accessed on 22 March 2022).
- Web BankIt. Available online: http://www.ncbi.nlm.nih.go-v/BankIt/index.html (accessed on 22 March 2022).
- Web BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 22 March 2022).
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- MITOS Web Server. Available online: http://mitos.bioinf.uni-leipzig.de/index.py (accessed on 22 March 2022).
- tRNAscan-SE Web Server. Available online: http://lowelab.ucsc.edu/tRNAscan-SE/ (accessed on 22 March 2022).
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Web RNAfold. Available online: http://rna.tbi.univie.ac.at//cgi-bin/RNAWebSuite/RNAfold.cgi (accessed on 22 March 2022).
- Comparative RNA Web (CRW) Site. Available online: http://www.rna.ccbb.utexas.edu/ (accessed on 22 March 2022).
- Hickson, R.E.; Simon, C.; Cooper, A.; Spicer, G.S.; Sullivan, J.; Penny, D. Conserved sequence motifs, alignment, and secondary structure for the third domain of animal 12S rRNA. Mol. Biol. Evol. 1996, 17, 530–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannone, J.J.; Subramanian, S.; Schnare, M.N.; Collett, J.R.; D’Souza, L.M.; Du, Y.; Feng, B.; Lin, N.; Madabusi, L.V.; Müller, K.M. The Comparative RNA Web (CRW) Site: An online database of comparative sequence and structure information for ribosomal, intron, and other RNAs. BMC Bioinform. 2002, 3, 2. [Google Scholar]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [Green Version]
- OGDRAW. Available online: https://chlorobox.mpimp-golm.mpg.de/OGDraw.html (accessed on 10 April 2022).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Tandem Repeats Finder Web Server. Available online: http://tandem.bu.edu/trf/trf.html (accessed on 22 March 2022).
- Du, Y.; Qiu, Q.B.; Tong, Q.L.; Lin, L.H. The complete mitochondrial genome of Eremias przewalskii (Squamata: Lacertidae). Mitochondrial DNA A 2016, 27, 1918–1919. [Google Scholar]
- Zhou, T.; Li, D.; Dujsebayeva, T.N.; Liu, J.; Guo, X. Complete mitochondrial genome of Stummer’s Racerunner (Eremias stummeri) from Kazakhstan. Mitochondrial DNA A 2016, 27, 4340–4341. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.L.; Yao, Y.T.; Lin, L.H.; Ji, X. The complete mitochondrial genome of Eremias vermiculata (Squamata: Lacertidae). Mitochondrial DNA 2016, 27, 1447–1448. [Google Scholar] [CrossRef] [PubMed]
- Rui, J.L.; Wang, Y.T.; Nie, L.W. The complete mitochondrial DNA genome of Eremias brenchleyi (Reptilia: Lacertidae) and its phylogeny position within Squamata reptiles. Amphib-Reptil. 2009, 30, 25–35. [Google Scholar] [CrossRef]
- Li, D.; Song, S. Complete mitochondrial genome of Eremias multiocellata. 2015; unpublished work. [Google Scholar]
- Su, X.; Liu, J.; Chen, D.; Guo, X. Next-generation sequencing yields a nearly complete mitochondrial genome of the Multiocellated Racerunner (Eremias multiocellata) in Northwest China. Mitochondrial DNA B 2019, 4, 1430–1431. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Yoo, D.U.; Hwang, U.W. Complete mitochondrial genome of Mongolia racerunner (Eremias argus). 2016; unpublished work. [Google Scholar]
- Yu, X.; Lin, L. The partial mitochondrial genome of Eremias arguta. 2021; unpublished work. [Google Scholar]
- Zhou, Z.S.; Li, H.; Tong, Q.L.; Lin, L.H.; Ji, X. The nearly complete mitochondrial genome of the rapid racerunner Eremias velox (Squamata: Lacertidae). Mitochondrial DNA A 2016, 27, 1781–1782. [Google Scholar]
- Murtskhvaladze, M.; Tarkhnishvili, D.; Anderson, C.L.; Kotorashvili, A. Phylogeny of caucasian rock lizards (Darevskia) and other true lizards based on mitogenome analysis: Optimisation of the algorithms and gene selection. PLoS ONE 2020, 15, e0233680. [Google Scholar] [CrossRef]
- Tao, C.R. The Complete Mitogenome of Lacerta agilis and the Phylogenetic Analysis of Squamata. Master’s Dissertation, Guangxi Normal University, Guilin, China, 2014. (In Chinese with English abstract). [Google Scholar]
- Margaryan, A. The partial mitochondrial genome of Lacerta agilis. 2020; unpublished work. [Google Scholar]
- Kolora, S.R.; Faria, R.; Weigert, A.; Schaffer, S.; Grimm, A.; Henle, K.; Sahyoun, A.H.; Stadler, P.F.; Nowick, K.; Bleidorn, C. The complete mitochondrial genome of Lacerta bilineata and comparison with its closely related congener L. viridis. Mitochondrial DNA A 2017, 28, 116–118. [Google Scholar] [CrossRef]
- Ma, W.W.; Liu, H.; Zhao, W.G.; Liu, P. The complete mitochondrial genome of Takydromus amurensis (Squamata: Lacertidae). Mitochondrial DNA B 2016, 1, 214–215. [Google Scholar] [CrossRef]
- Wu, L.X.; Luo, K.N.; Ding, G.H. Complete mitochondrial genome of Takydromus kuehnei. 2021; unpublished work. [Google Scholar]
- Hu, J.G.; Peng, L.F.; Tang, X.S.; Huang, S. The complete mitochondrial genome of Takydromus septentrionalis (Reptilia: Lacertidae). Mitochondrial DNA B 2019, 4, 2193–2194. [Google Scholar] [CrossRef] [Green Version]
- Qin, P.S.; Zeng, D.L.; Hou, L.X.; Yang, X.W.; Qin, X.M. Complete mitochondrial genome of Takydromus sexlineatus (Squamata, Lacertidae). Mitochondrial DNA 2015, 26, 465–466. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.S.; Chen, J.M.; Huang, S. Mitochondrial genome of the Chung-an ground lizard Takydromus sylvaticus (Reptilia: Lacertidae). Mitochondrial DNA 2014, 25, 319–320. [Google Scholar] [CrossRef]
- Kumazawa, Y. Mitochondrial genomes from major lizard families suggest their phylogenetic relationships and ancient radiations. Gene 2007, 388, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.N.; Ji, X. The complete mitochondrial genome of Takydromus wolteri (Squamata: Lacertidae). Mitochondrial DNA 2013, 24, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhu, D.; Zhao, W.G.; Ji, X. The complete mitochondrial genome of the common lizard Zootoca vivipara (Squamata: Lacertidae). Mitochondrial DNA 2016, 27, 1944–1945. [Google Scholar]
- Macey, J.R.; Papenfuss, T.J.; Kuehl, J.V.; Fourcade, H.M.; Boore, J.L. Phylogenetic relationships among amphisbaenian reptiles based on complete mitochondrial genome sequences. Mol. Phylogenet. Evol. 2004, 33, 22–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunes, T.O.; Lyra, M.L.; Maldonado, J.A.; Pellegrino, K.C.M.; Rodrigues, M.T.; Fujita, M.K. The first mitochondrial genome of a South America parthenogenetic lizard (Squamata: Gymnophthalmidae). Mitochondrial DNA B 2021, 6, 2393–2395. [Google Scholar] [CrossRef]
- GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 22 March 2022).
- Wiens, J.J.; Hutter, C.R.; Mulcahy, D.G.; Noonan, B.P.; Townsend, T.M.; Sites, J.W., Jr.; Reeder, T.W. Resolving the phylogeny of lizards and snakes (Squamata) with extensive sampling of genes and species. Biol. Lett. 2012, 8, 1043–1046. [Google Scholar] [CrossRef] [Green Version]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenet. Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Li, W.X.; Jakovlić, I.; Zou, H.; Zhang, J.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. Mrbayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erixon, P.; Svennblad, B.; Britton, T.; Oxelman, B. Reliability of Bayesian posterior probabilities and bootstrap frequencies in phylogenetics. Syst. Biol. 2003, 52, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Rannala, B. Frequentist properties of Bayesian posterior probabilities of phylogenetic trees under simple and complex substitution models. Syst. Biol. 2004, 53, 904–913. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; Haeseler, A.V. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- FigTree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 22 March 2022).
- Yu, P.; Zhou, L.; Yang, W.T.; Miao, L.J.; Li, Z.; Zhang, X.J.; Wang, Y.; Gui, J.F. Comparative mitogenome analyses uncover mitogenome features and phylogenetic implications of the subfamily Cobitinae. BMC Genomics 2021, 22, 50. [Google Scholar] [CrossRef]
- Topal, M.D.; Fresco, J.R. Complementary base pairing and the origin of substitution mutations. Nature 1976, 263, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Castellana, S.; Vicario, S.; Saccone, C. Evolutionary patterns of the mitochondrial genome in Metazoa: Exploring the role of mutation and selection in mitochondrial protein coding genes. Genome Biol. Evol. 2011, 3, 1067–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southern, S.O.; Southern, P.J.; Dizon, A.E. Molecular characterization of a cloned dolphin mitochondrial genome. J. Mol. Evol. 1988, 28, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.G.; Gadaleta, G.; Pepe, G.; Saccone, C.; Sbisà, E. Structural conservation and variation in the D-loop-containing region of vertebrate mitochondrial DNA. J. Mol. Biol. 1986, 192, 503–511. [Google Scholar] [CrossRef]
- Orlova, V.F.; Poyarkov, N.A., Jr.; Chirikova, M.A.; Nazarov, R.A.; Munkhbaatar, M.; Munkhbayar, K.; Terbish, M. MtDNA differentiation and taxonomy of Central Asian racerunners of Eremias multiocellata-E. przewalskii species complex (Squamata, Lacertidae). Zootaxa 2017, 4282, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Dixon, M.T.; Hillis, D.M. Ribosomal RNA secondary structure: Compensatory mutations and implications for phylogenetic analysis. Mol. Biol. Evol. 1993, 10, 256–267. [Google Scholar]
- Springer, M.S.; Hollar, L.J.; Burk, A. Compensatory substitutions and the evolution of the mitochondrial 12S rRNA gene in mammals. Mol. Biol. Evol. 1995, 12, 1138–1150. [Google Scholar]
- Kjer, K.M. Use of rRNA secondary structure in phylogenetic studies to identify homologous positions: An example of alignment and data presentation from the frogs. Mol. Phylogenet. Evol. 1995, 4, 314–330. [Google Scholar] [CrossRef]
- Hudelot, C.; Gowri-Shankar, V.; Jow, H.; Rattray, M.; Higgs, P.G. RNA-based phylogenetic methods: Application to mammalian mitochondrial RNA sequences. Mol. Phylogenet. Evol. 2003, 28, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Schöniger, M.; von Haeseler, A. A stochastic model for the evolution of autocorrelated DNA sequences. Mol. Phylogenet. Evol. 1994, 3, 240–247. [Google Scholar] [CrossRef]
- Telford, M.J.; Wise, M.J.; Gowri-Shankar, V. Consideration of RNA secondary structure significantly improves likelihood-based estimates of phylogeny: Examples from the Bilateria. Mol. Biol. Evol. 2005, 22, 1129–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jow, H.; Hudelot, C.; Rattray, M.; Higgs, P.G. Bayesian phylogenetics using an RNA substitution model applied to early mammalian evolution. Mol. Biol. Evol. 2002, 19, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, X.; Kong, X.; Zhao, K.; He, S.; Mayden, R.L. Variation patterns of the mitochondrial 16S rRNA gene with secondary structure constraints and their application to phylogeny of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2008, 47, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. Preponderance of synonymous changes as evidence for the neutral theory of molecular evolution. Nature 1977, 267, 275–276. [Google Scholar] [CrossRef]
- Yang, Z.; Bielawski, J.P. Statistical methods for detecting molecular adaptation. Trends Ecol. Evol. 2000, 15, 496–503. [Google Scholar] [CrossRef]
- Magnus, W.J.; Rute, R.D.F.; Louis, B.; Michael, M.H. Comparative analysis of complete mitochondrial genomes suggests that relaxed purifying selection is driving high nonsynonymous evolutionary rate of the NADH2 gene in whitefish (Coregonus ssp.). Mol. Phylogenet. Evol. 2016, 95, 161–170. [Google Scholar]
- Melville, R.V. Opinion 1318 (Opinion correcting the ruling given in Opinion 92) Lacerta velox Pallas, 1771 is the type species of Eremias Wiegmann, 1834. Bull. Zooll. Nomencl. 1985, 42, 171–172. [Google Scholar]
- Brown, W.M.; George, M., Jr.; Wilson, A.C. Rapid evolution of animal mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1979, 76, 1967–1971. [Google Scholar] [CrossRef] [Green Version]
- Doda, J.N.; Wright, C.T.; Clayton, D.A. Elongation of displacement loop strands in human and mouse mitochondrial DNA is arrested near specific template sequences. Proc. Natl. Acad. Sci. USA 1981, 78, 6116–6120. [Google Scholar] [CrossRef] [Green Version]
- Saccone, C.; Pesole, G.; Sbisà, E. The main regulatory region of mammalian mitochondrial DNA: Structure-function model and evolutionary pattern. J. Mol. Evol. 1991, 33, 83–91. [Google Scholar] [CrossRef]
- Gao, S.; Tian, X.; Chang, H.; Sun, Y.; Wu, Z.; Cheng, Z.; Dong, P.; Zhao, Q.; Ruan, J.; Bu, W. Two novel lncRNAs discovered in human mitochondrial DNA using PacBio full-length transcriptome data. Mitochondrion 2018, 38, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Xu, X.; Jin, X.; Yin, H.; Luo, J.; Liu, G.; Zhao, Q.; Chen, Z.; Bu, W.; Gao, S. Using high-resolution annotation of insect mitochondrial DNA to decipher tandem repeats in the control region. RNA Biol. 2019, 16, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ji, H.; Jin, X.; Cheng, Z.; Yao, X.; Liu, Y.; Zhao, Q.; Zhang, T.; Ruan, J.; Bu, W.; et al. Using pan RNA-seq analysis to reveal the ubiquitous existence of 5′ and 3′ end small RNAs. Front. Genet. 2019, 10, 105. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher Number | Collection Date | Collection Site |
|---|---|---|---|
| E. scripta | KZL15 | July 2014 | Kazakhstan; 45.91764° N, 79.96855° E |
| E. scripta | KZL44 | July 2014 | Kazakhstan; 45.91764° N, 79.96855° E |
| E. nikolskii | Guo4717 | August 2014 | Kyrgyzstan; 41.38750° N, 73.93999° E |
| E. szczerbaki | Guo4719 | August 2014 | Kyrgyzstan; 41.48393° N, 75.97943° E |
| E. yarkandensis | Guo4722 | August 2014 | Kyrgyzstan; 39.64809° N, 73.86512° E |
| Taxon | Family | Subfamily | Accession Number | Length (bp) | Reference |
|---|---|---|---|---|---|
| Eremias | |||||
| E. scripta KZL15 | Lacertidae | Eremiainae | OM935765 | 19,824 | This study |
| E. scripta KZL44 | Lacertidae | Eremiainae | OM935766 | 19,831 | This study |
| E. nikolskii | Lacertidae | Eremiainae | OK587334 | 20,840 | This study |
| E. szczerbaki | Lacertidae | Eremiainae | OL457296 | 19,650 | This study |
| E. yarkandensis | Lacertidae | Eremiainae | OK585048 | 18,743 | This study |
| E. dzungarica | Lacertidae | Eremiainae | MW250881 | 19,899 | [19] |
| E. przewalskii | Lacertidae | Eremiainae | KM507330 | 18,225 | [52] |
| E. stummeri | Lacertidae | Eremiainae | KT372881 | 19,602 | [53] |
| E. vermiculata | Lacertidae | Eremiainae | KM104865 | 19,914 | [54] |
| E. brenchleyi | Lacertidae | Eremiainae | EF490071 | 19,542 | [55] |
| E. multiocellata | Lacertidae | Eremiainae | KJ664798 | 18,996 | [56] |
| E. multiocellata | Lacertidae | Eremiainae | MK261077 | 17,333 | [57] |
| E. argus | Lacertidae | Eremiainae | JQ086345 | 18,521 | [58] |
| E. arguta | Lacertidae | Eremiainae | KU605241 | 15,192 | [59] |
| E. grammica | Lacertidae | Eremiainae | KU585904 | 15,338 | [59] |
| E. velox | Lacertidae | Eremiainae | KM359148 | 18,033 | [60] |
| Acanthodactylus | |||||
| Ac. aureus | Lacertidae | Eremiainae | MW496121 | 15,756 | [15] |
| Ac. boskianus | Lacertidae | Eremiainae | MW496112 | 17,143 | [15] |
| Ac. erythrurus | Lacertidae | Eremiainae | MW496113 | 16,827 | [15] |
| Ac. guineensis | Lacertidae | Eremiainae | MW496123 | 16,963 | [15] |
| Ac. schmidti | Lacertidae | Eremiainae | MW496124 | 16,943 | [15] |
| Australolacerta | |||||
| Au. australis | Lacertidae | Eremiainae | MW496118 | 17,019 | [15] |
| Meroles | |||||
| Mer. squamulosus | Lacertidae | Eremiainae | MW496120 | 16,860 | [15] |
| Mesalina | |||||
| Mes. olivieri | Lacertidae | Eremiainae | MW496114 | 16,899 | [15] |
| Pedioplanis | |||||
| Pe. laticeps | Lacertidae | Eremiainae | MW496119 | 17,046 | [15] |
| Algyroides | |||||
| Al. nigropunctatus | Lacertidae | Lacertinae | MW496122 | 15,844 | [15] |
| Darevskia | |||||
| D. armeniaca | Lacertidae | Lacertinae | MG704915 | 17,521 | [61] |
| D. brauneri | Lacertidae | Lacertinae | MH481137 | 16,976 | [61] |
| D. caucasica | Lacertidae | Lacertinae | MH481131 | 16,343 | [61] |
| D. chlorogaster | Lacertidae | Lacertinae | MH481136 | 17,479 | [61] |
| D. clarkorum | Lacertidae | Lacertinae | MH481134 | 16,301 | [61] |
| D. dahli | Lacertidae | Lacertinae | MG704916 | 17,528 | [61] |
| D. daghestanica | Lacertidae | Lacertinae | MH481135 | 17,189 | [61] |
| D. derjugini | Lacertidae | Lacertinae | MH481130 | 16,960 | [61] |
| D. mixta | Lacertidae | Lacertinae | MG704917 | 17,532 | [61] |
| D. parvula | Lacertidae | Lacertinae | MG704918 | 17,510 | [61] |
| D. portschinskii | Lacertidae | Lacertinae | MG704919 | 17,529 | [61] |
| D. praticola | Lacertidae | Lacertinae | MH481132 | 16,418 | [61] |
| D. raddei | Lacertidae | Lacertinae | MH481133 | 20,478 | [61] |
| D. rudis | Lacertidae | Lacertinae | MG704920 | 17,534 | [61] |
| D. saxicola | Lacertidae | Lacertinae | MG704921 | 17,524 | [61] |
| D. unisexualis | Lacertidae | Lacertinae | KX644918 | 21,433 | [61] |
| D. valentini | Lacertidae | Lacertinae | MG655240 | 17,393 | [61] |
| Lacerta | |||||
| L. agilis | Lacertidae | Lacertinae | KC990830 | 17,090 | [62] |
| L. agilis | Lacertidae | Lacertinae | MT410899 | 17,069 | [63] |
| L. bilineata | Lacertidae | Lacertinae | KT722705 | 17,154 | [64] |
| L. viridis viridis | Lacertidae | Lacertinae | AM176577 | 17,156 | [21] |
| Phoenicolacerta | |||||
| Ph. kulzeri | Lacertidae | Lacertinae | FJ460596 | 17,199 | [22] |
| Podarcis | |||||
| Po. muralis | Lacertidae | Lacertinae | FJ460597 | 17,311 | [22] |
| Po. siculus | Lacertidae | Lacertinae | FJ460598 | 17,297 | [22] |
| Takydromus | |||||
| T. amurensis | Lacertidae | Lacertinae | KU641018 | 17,333 | [65] |
| T. kuehnei | Lacertidae | Lacertinae | MZ435950 | 17,224 | [66] |
| T. septentrionalis | Lacertidae | Lacertinae | MK630237 | 18,304 | [67] |
| T. sexlineatus | Lacertidae | Lacertinae | KF425529 | 18,943 | [68] |
| T. sylvaticus | Lacertidae | Lacertinae | JX290083 | 17,838 | [69] |
| T. tachydromoides | Lacertidae | Lacertinae | AB080237 | 18,245 | [70] |
| T. wolteri | Lacertidae | Lacertinae | JX181764 | 18,236 | [71] |
| Zootoca | |||||
| Z. vivipara | Lacertidae | Lacertinae | KM401599 | 17,046 | [72] |
| Psammodromus | |||||
| Ps. algirus | Lacertidae | Gallotiinae | MW496117 | 17,118 | [15] |
| Outgroup | |||||
| Amphisbaena | |||||
| Am. schmidti | Amphisbaenidae | − | AY605475 | 17,423 | [73] |
| Bipes | |||||
| B. biporus | Bipedidae | − | AY605481 | 16,430 | [73] |
| Loxopholis | |||||
| Lo. percainatum | Gymnophthalmidae | − | MW864329 | 15,875 | [74] |
| Type | Number of Species | TAS1 | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L. dugesii | A | C | T | A | T | T | A | T | G | T | A | T | A | T | A | G | T | G | C | A | T | T | A | A | |
| 1 | 27 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 2 | 11 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | A | · | · | · | · | · | · |
| 3 | 1 | · | · | · | · | · | · | · | C | · | G | · | · | · | · | · | · | C | · | · | · | · | · | · | C |
| 4 | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | T | · | · | A | · | · | · | · | · | · |
| 5 | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | C | · | · | A | · | · | · | · | · | · |
| 6 | 1 | · | · | A | T | · | · | · | · | · | · | · | · | · | A | T | · | · | A | · | · | · | · | · | · |
| 7 | 1 | · | · | A | · | · | · | · | · | · | · | · | · | · | · | · | T | · | · | · | · | · | · | C | A |
| 8 | 1 | · | · | A | T | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | G |
| 9 | 1 | · | · | · | · | C | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| Type | Number of Species | CSB1 | |||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L. dugesii | C | T | A | T | A | T | G | G | T | A | T | T | A | T | T | G | T | C | T | T | A | A | T | G | C | T | T | G | G | T | A | G | A | C | A | T | A | T | |
| 1 | 12 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 2 | 13 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | C | · | T | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 3 | 2 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | T | C | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 4 | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | T | C | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 5 | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | T | C | T | · | G | · | · | · | · | · | · | · | · | C | · | · | · | · | · | · | · | · | · |
| 6 | 1 | · | · | · | · | T | · | · | · | · | · | · | · | · | C | · | T | C | A | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 7 | 1 | · | · | · | · | · | · | · | · | · | · | · | · | G | · | · | C | · | T | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 8 | 1 | · | · | · | · | · | · | · | · | · | G | · | · | · | · | · | · | C | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 9 | 2 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | G | A | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 10 | 2 | · | · | · | · | · | · | · | · | · | · | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 11 | 2 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 12 | 3 | · | · | · | · | T | · | · | · | · | · | · | · | · | · | · | · | · | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 13 | 1 | · | · | · | · | · | · | · | · | · | G | · | · | · | · | · | · | A | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 14 | 1 | · | · | · | · | · | · | · | · | G | G | C | · | · | · | · | · | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 15 | 1 | · | · | · | · | T | · | · | · | · | G | · | · | · | · | · | · | · | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 16 | 1 | · | · | · | · | T | · | · | · | · | · | · | · | · | · | · | T | A | · | · | G | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 17 | 1 | · | · | · | · | · | A | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | A |
| 18 | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | A | · | · | · | · | · | · | · | · | · | · | A | · | · | · | · | · | · | · | · | · |
| Type | Number of Species | CSB2 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L. dugesii | C | A | A | A | C | C | C | C | C | C | T | A | C | C | C | C | C | C | |
| 1 | 41 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 2 | 1 | · | · | · | · | T | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 3 | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | T | · | · | · |
| 4 | 1 | T | · | · | · | · | T | · | · | T | · | · | · | · | · | · | · | · | T |
| 5 | 1 | · | · | · | · | T | · | T | · | T | · | · | · | · | · | T | · | · | · |
| 6 | 1 | · | · | · | · | · | T | · | · | · | · | · | G | · | · | · | · | · | T |
| Type | Number of Species | TAS2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| L. dugesii | C | A | T | A | C | A | T | T | A | A | |
| 1 | 1 | · | · | · | · | · | · | · | · | · | · |
| 2 | 13 | T | · | · | G | · | · | · | · | · | · |
| 3 | 5 | T | · | · | · | · | · | · | · | · | · |
| 4 | 5 | T | · | · | · | · | · | · | · | · | T |
| 5 | 2 | T | · | · | · | T | · | · | · | · | · |
| 6 | 1 | T | · | · | · | A | · | · | · | · | · |
| 7 | 1 | T | · | · | · | · | · | · | · | · | C |
| 8 | 1 | T | · | · | · | · | · | · | A | · | T |
| 9 | 1 | T | · | · | C | · | · | · | · | · | · |
| 10 | 3 | A | · | · | · | · | · | · | · | · | T |
| 11 | 1 | A | G | · | · | · | · | · | · | · | · |
| 12 | 1 | A | · | · | · | · | · | · | · | · | · |
| 13 | 1 | · | · | · | · | · | · | · | · | · | T |
| 14 | 2 | · | · | · | G | · | · | · | · | · | · |
| 15 | 1 | · | · | · | G | T | · | · | · | · | · |
| 16 | 1 | · | G | · | · | · | · | · | · | · | · |
| 17 | 1 | · | · | · | · | T | · | · | · | T | · |
| 18 | 2 | · | · | · | · | T | · | · | · | · | · |
| 19 | 1 | · | · | · | · | · | T | · | · | · | · |
| 20 | 1 | · | · | · | · | T | · | · | T | · | · |
| 21 | 1 | · | · | · | · | A | C | · | · | G | · |
| 22 | 1 | · | · | · | C | · | G | A | · | · | · |
| 23 | 1 | · | · | · | A | · | · | T | · | · | · |
| 24 | 1 | · | · | · | · | · | · | · | A | · | T |
| 25 | 1 | · | · | · | · | · | · | · | · | C | · |
| 26 | 1 | · | · | · | · | · | A | · | · | · | · |
| Type | Number of Species | CSB3 | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L. dugesii | T | C | G | C | C | A | A | A | C | C | C | C | T | A | A | A | A | C | G | A | |
| 1 | 36 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · |
| 2 | 5 | · | · | · | · | · | · | · | · | · | · | · | · | A | · | · | · | · | · | · | · |
| 3 | 1 | · | · | · | T | · | · | · | · | · | · | A | · | · | · | · | · | · | · | · | · |
| 4 | 1 | · | · | · | · | · | · | · | · | T | · | · | · | · | · | · | · | · | · | · | · |
| 5 | 1 | C | · | · | · | · | · | · | · | T | T | · | · | C | G | · | · | · | · | C | · |
| 6 | 1 | · | · | · | · | · | · | · | · | · | · | T | · | G | · | · | · | · | · | A | · |
| 7 | 1 | · | · | · | · | · | · | · | · | T | · | · | · | · | · | G | · | · | · | A | G |
| 8 | 1 | A | G | G | · | · | · | A | T | T | · | · | · | · | · | · | T | · | T | · | · |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, L.; Guo, X. Complete Mitochondrial Genomes of Five Racerunners (Lacertidae: Eremias) and Comparison with Other Lacertids: Insights into the Structure and Evolution of the Control Region. Genes 2022, 13, 726. https://doi.org/10.3390/genes13050726
Tian L, Guo X. Complete Mitochondrial Genomes of Five Racerunners (Lacertidae: Eremias) and Comparison with Other Lacertids: Insights into the Structure and Evolution of the Control Region. Genes. 2022; 13(5):726. https://doi.org/10.3390/genes13050726
Chicago/Turabian StyleTian, Lili, and Xianguang Guo. 2022. "Complete Mitochondrial Genomes of Five Racerunners (Lacertidae: Eremias) and Comparison with Other Lacertids: Insights into the Structure and Evolution of the Control Region" Genes 13, no. 5: 726. https://doi.org/10.3390/genes13050726