
Ultrastructural Changes in Mitochondria in Patients with Dilated Cardiomyopathy and Parvovirus B19 Detected in Heart Tissue without Myocarditis

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Endomyocardial Biopsy
2.3. Quantitative Real-Time PCR (qPCR)
2.4. Transmission Electron Microscopy (TEM)
2.5. Statistical Methods
3. Results
3.1. Study Population
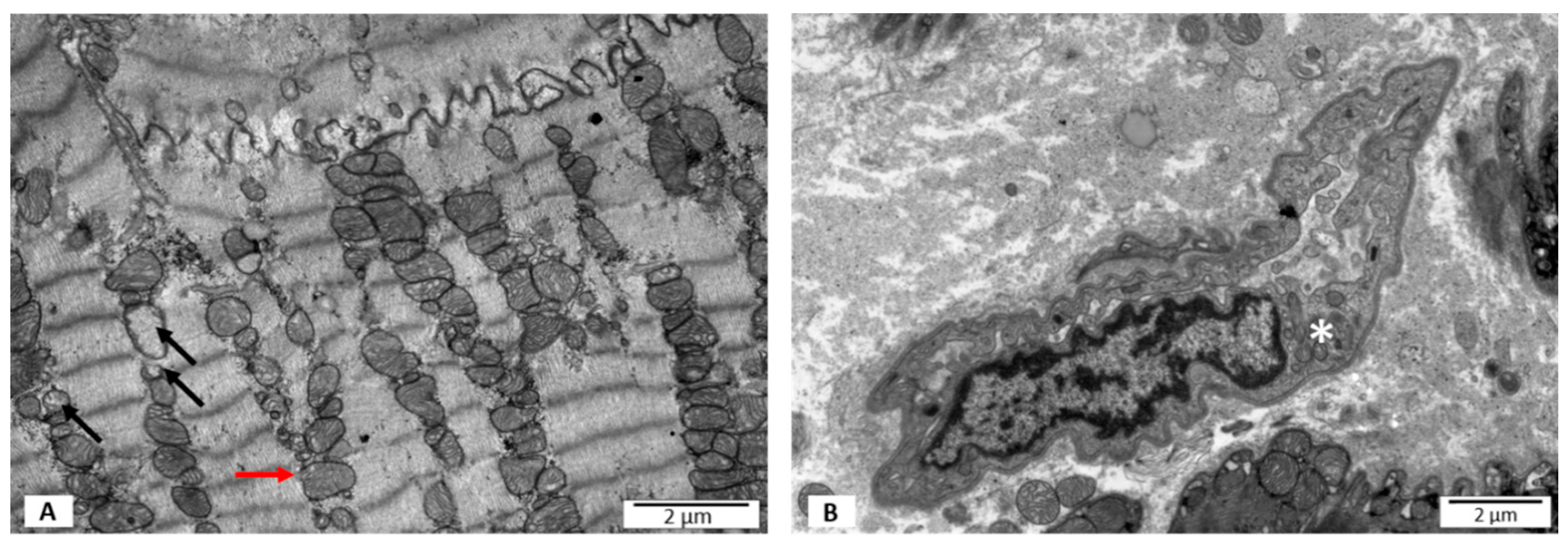
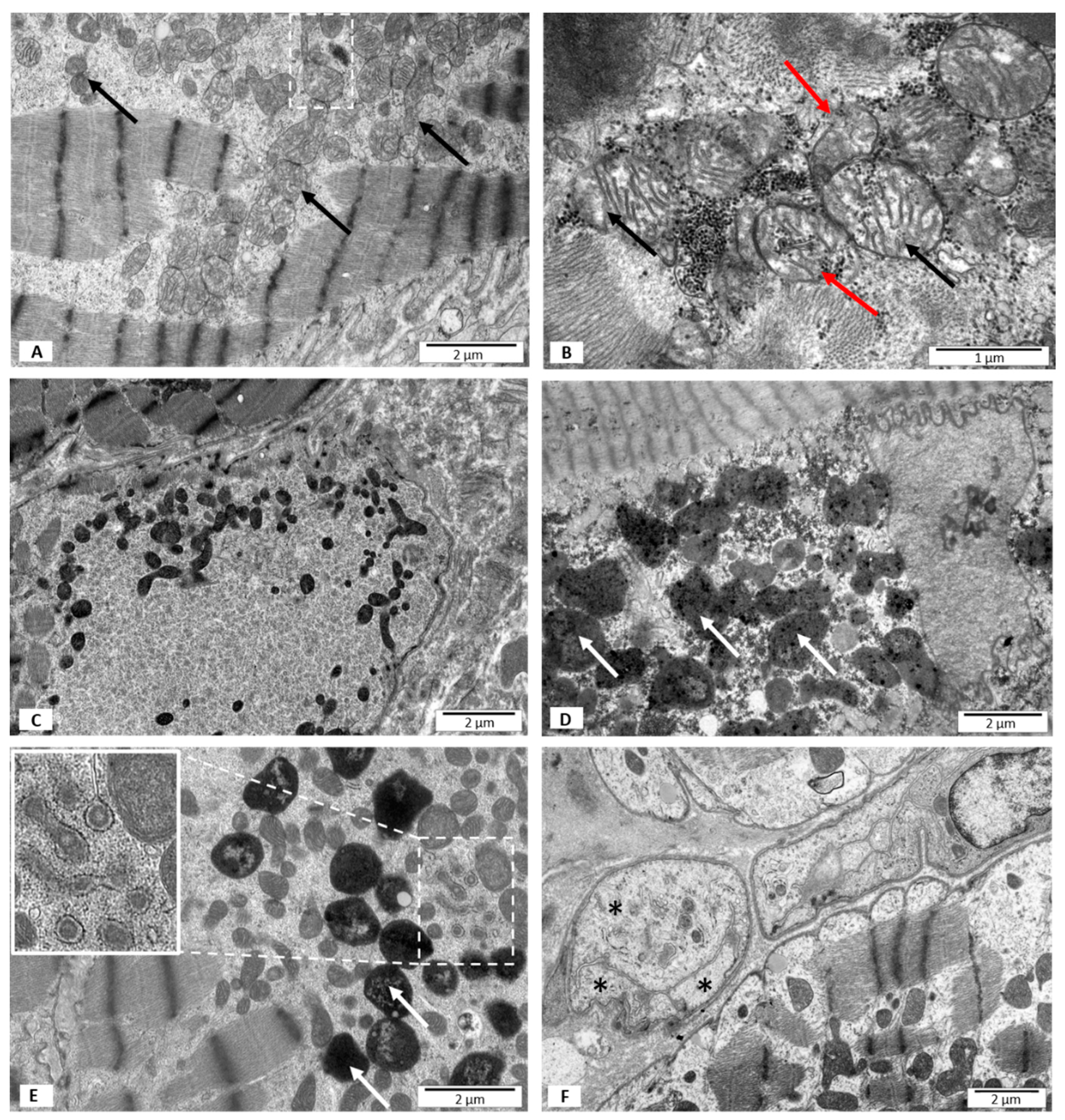
3.2. Electron Microscopy Studies
3.2.1. Group I—LVEF ≥ 50%, without PB19V DNA
3.2.2. Group II—LVEF < 50%, without PB19V DNA
3.2.3. Group III—LVEF < 50%, with B19V DNA
4. Discussion
5. Conclusions
6. Study Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kindermann, I.; Barth, C.; Mahfoud, F.; Ukena, C.; Lenski, M.; Yilmaz, A.; Klingel, K.; Kandolf, R.; Sechtem, U.; Cooper, L.T.; et al. Update on myocarditis. J. Am. Coll. Cardiol. 2012, 59, 779–792. [Google Scholar] [CrossRef]
- Felker, G.M.; Hu, W.; Hare, J.M.; Hruban, R.H.; Baughman, K.L.; Kasper, E.K. The Spectrum of Dilated Cardiomyopathy: The Johns Hopkins Experience with 1278 Patients. Medicine 1999, 78, 270–283. [Google Scholar] [CrossRef]
- Kühl, U.; Pauschinger, M.; Noutsias, M.; Seeberg, B.; Bock, T.; Lassner, D.; Poller, W.; Kandolf, R.; Schultheiss, H.-P. High prevalence of viral genomes and multiple viral infections in the myocardium of adults with “idiopathic” left ventricular dysfunction. Circulation 2005, 111, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Klein, R.M.; Jiang, H.; Niederacher, D.; Adams, O.; Du, M.; Horlitz, M.; Schley, P.; Marx, R.; Lankisch, M.R.; Brehm, M.U.; et al. Frequency and quantity of the parvovirus B19 genome in endomyocardial biopsies from patients with suspected myocarditis or idiopathic left ventricular dysfunction. Z. Kardiol. 2004, 93, 300–309. [Google Scholar] [CrossRef]
- Singh, S.P.; Amar, S.; Gehlot, P.; Patra, S.K.; Kanwar, N.; Kanwal, A. Mitochondrial Modulations, Autophagy Pathways Shifts in Viral Infections: Consequences of COVID-19. Int. J. Mol. Sci. 2021, 22, 8180. [Google Scholar] [CrossRef]
- Twig, G.; Elorza, A.; Molina, A.J.; Mohamed, H.; Wikstrom, J.D.; Walzer, G.; Stiles, L.; Haigh, S.E.; Katz, S.; Las, G.; et al. Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J. 2008, 27, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Lehman, J.J.; Barger, P.M.; Kovacs, A.; Saffitz, J.E.; Medeiros, D.M.; Kelly, D.P. Peroxisome proliferator–activated receptor γ coactivator-1 promotes cardiac mitochondrial biogenesis. J. Clin. Investig. 2000, 106, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Pillai, V.B.; Bindu, S.; Sharp, W.; Fang, Y.H.; Kim, G.; Gupta, M.; Samant, S.; Gupta, M.P. Sirt3 protects mitochondrial DNA damage and blocks the development of doxorubicin-induced cardiomyopathy in mice. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H962–H972. [Google Scholar] [CrossRef] [Green Version]
- Goh, K.Y.; Qu, J.; Hong, H.; Liu, T.; Dell’Italia, L.J.; Wu, Y.; O’Rourke, B.; Zhou, L. Impaired mitochondrial network excitability in failing guinea-pig cardiomyocytes. Cardiovasc. Res. 2016, 109, 79–89. [Google Scholar] [CrossRef]
- Lemasters, J.J. Variants of mitochondrial autophagy: Types 1 and 2 mitophagy and micromitophagy (Type 3). Redox Biol. 2014, 2, 749–754. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, T.P.; Bjorklund, M. Mitochondrial Function and Cell Size: An Allometric Relationship. Trends Cell Biol. 2017, 27, 393–402. [Google Scholar] [CrossRef]
- Siasos, G.; Tsigkou, V.; Kosmopoulos, M.; Theodosiadis, D.; Simantiris, S.; Tagkou, N.M.; Tsimpiktsioglou, A.; Stampouloglou, P.K.; Oikonomou, E.; Mourouzis, K.; et al. Mitochondria and cardiovascular diseases-from pathophysiology to treatment. Ann. Transl. Med. 2018, 6, 256. [Google Scholar] [CrossRef]
- Aretz, H.T. Myocarditis. The Dallas criteria. Hum. Pathol. 1987, 18, 619–624. [Google Scholar] [CrossRef]
- Caforio, A.L.; Pankuweit, S.; Arbustini, E.; Basso, C.; Gimeno-Blanes, J.; Felix, S.B.; Fu, M.; Heliö, T.; Heymans, S.; Jahns, R.; et al. Current state of knowledge on aetiology, diagnosis, management, and therapy of myocarditis: A position statement of the European Society of Cardiology Working Group on Myocardial and Pericardial Diseases. Eur. Heart J. 2013, 34, 2636–2648. [Google Scholar] [CrossRef]
- Pawlak, A.; Wiligorska, N.; Wiligorska, D.; Frontczak-Baniewicz, M.; Przybylski, M.; Krzyzewski, R.; Ziemba, A.; Gil, R.J. Viral Heart Disease and Acute Coronary Syndromes—Often or Rare Coexistence? Curr. Pharm. Des. 2018, 24, 532–540. [Google Scholar] [CrossRef]
- Czech, M.; Opolski, G.; Zdrojewski, T.; Dubiel, J.S.; Wizner, B.; Bolisęga, D.; Fedyk-Łukasik, M.; Grodzicki, T. The costs of heart failure in Poland from the public payer’s perspective. Polish programme assessing diagnostic procedures, treatment and costs in patients with heart failure in randomly selected outpatient clinics and hospitals at different levels of care: POLKARD. Kardiol. pol. 2013, 71, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Kuethe, F.; Lindner, J.; Matschke, K.; Wenzel, J.J.; Norja, P.; Ploetze, K.; Schaal, S.; Kamvissi, V.; Bornstein, S.R.; Schwanebeck, U.; et al. Prevalence of parvovirus B19 and human bocavirus DNA in the heart of patients with no evidence of dilated cardiomiopathy or myocarditis. Clin. Infect. Dis. 2009, 49, 1660–1666. [Google Scholar] [CrossRef] [Green Version]
- Klingel, K.; Sauter, M.; Bock, C.T.; Szalay, G.; Schnorr, J.J.; Kandolf, R. Molecular pathology of inflammatory cardiomyopathy. Med. Microbiol. Immunol. 2004, 193, 101–107. [Google Scholar] [CrossRef]
- Wei, J.; Gao, D.G.; Wang, H.; Yan, R.; Liu, Z.Q.; Yuan, Z.Y.; Liu, G.; Chen, M.X. Impairment of Myocardial Mitochondria in Viral Myocardial Disease and Its Reflective Window in Peripheral Cells. PLoS ONE 2014, 9, e116239. [Google Scholar] [CrossRef]
- Nykky, J.; Vuento, M.; Gilbert, L. Role of Mitochondria in Parvovirus Pathology. PLoS ONE 2014, 9, e86124. [Google Scholar] [CrossRef]
- Eisner, V.; Cupo, R.R.; Gao, E.; Csordás, G.; Slovinsky, W.S.; Paillard, M.; Cheng, L.; Ibetti, J.; Chen, S.R.; Chuprun, J.K. Mitochondrial fusion dynamics is robust in the heart and depends on calcium oscillations and contractile activity. Proc. Natl. Acad. Sci. USA 2017, 114, E859–E868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Sun, L.; Ji, S.; Zhao, T.; Zhang, W.; Xu, J.; Zhang, J.; Wang, Y.; Wang, X.; Franzini-Armstrong, C.; et al. Kissing and nanotunneling mediate intermitochondrial communication in the heart. Proc. Natl. Acad. Sci. USA 2013, 110, 2846–2851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavorato, M.; Iyer, V.R.; Dewight, W.; Cupo, R.R.; Debattisti, V.; Gomez, L.; De la Fuente, S.; Zhao, Y.T.; Valdivia, H.H.; Hajnóczky, G.; et al. Increased mitochondrial nanotunneling activity, induced by calcium imbalance, affects intermitochondrial matrix exchanges. Proc. Natl. Acad. Sci. USA 2017, 114, E849–E858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, A.; Tanaka, N.; Tamai, K.; Kyuuma, M.; Ishikawa, Y.; Sato, H.; Yoshimori, T.; Saito, S.; Sugamura, K. Survival of parvovirus B19-infected cells by cellular autophagy. Virology 2006, 349, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, S.E.; Sena, L.A.; Chandel, N.S. Mitochondria in the regulation of innate and adaptive immunity. Immunity 2015, 42, 406–417. [Google Scholar] [CrossRef] [Green Version]
- Banoth, B.; Cassel, S.L. Mitochondria in innate immune signaling. Transl. Res. 2018, 202, 52–68. [Google Scholar] [CrossRef]
- Xia, M.; Meng, G.; Li, M.; Wei, J. Mitophagy in viral infections. DNA Cell Biol. 2014, 33, 739–742. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Y.; Chen, M. Viral strategies for triggering and manipulating mitophagy. Autophagy 2018, 14, 1665–1673. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.H.; Kim, J.K.; Jo, E.K. Mitophagy and innate immunity in infection. Mol. Cells 2020, 43, 10–22. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, A. Virus-induced ER stress and the unfolded protein response. Front. Plant Sci. 2012, 3, 293. [Google Scholar] [CrossRef] [Green Version]
- Iwata, Y.; Koizumi, N. Plant transducers of the endoplasmic reticulum unfolded protein response. Trends Plant Sci. 2012, 17, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Binder, P.; Fang, Q.; Wang, Z.; Xiao, W.; Liu, W.; Wang, X. Endoplasmic reticulum stress in the heart: Insights into mechanisms and drug targets. Br. J. Pharmacol. 2018, 175, 1293–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bültmann, B.D.; Klingel, K.; Sotlar, K.; Bock, C.T.; Baba, H.A.; Sauter, M.; Kandolf, R. Fatal parvovirus B19-associated myocarditis clinically mimicking ischemic heart disease: An endothelial cell-mediated disease. Hum. Pathol. 2003, 34, 92–95. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Control Group (n = 7) | Patients with DCM without Virus (10) | Patients with DCM and Virus (10) | p |
|---|---|---|---|---|
| Male, n (%) | 7 (100) | 10 (100) | 10 (100) | 0.97 |
| Age, years | 28.0 ± 3.2 | 27.0 ± 6.3 | 28.7 ± 6.5 | 0.79 |
| NYHA Class, n (%) | ||||
| Class I | 2 (29) | 1(10) | 1 (10) | 0.14 |
| Class II | 5 (71) | 7 (70) | 6 (60) | 0.76 |
| Class III | 0 | 2 (20) | 3 (30) | 0.07 |
| Class IV | 0 | 0 | 0 | 0.98 |
| LVEF, (%) | 61.3 ± 2.3 | 23.3 ± 10.4 | 21.7 ± 6.1 | 0.01 * |
| LVEDD, (mm) | 47.7 ± 2.1 | 64.0 ± 7.2 | 62.3 ± 9.5 | 0.01 * |
| IVSDD, (mm) | 10.5 ± 0.7 | 9.3 ± 1.1 | 10.3 ± 1.1 | 0.08 |
| PWDD, (mm) | 10.0 ± 0.9 | 9.3 ± 1.1 | 10.0 ± 1.0 | 0.11 |
| NT-pro-BNP, (pg/dL) | 398.6 ± 123.2 | 609.3 ± 126.3 | 2281.6 ± 313.4 | 0.001 ** |
| CRP > 5 mg/L, n (%) | 13.9 ± 4.2 | 10.2 ± 3.9 | 33.8 ± 7.2 | 0.05 ** |
| Leukocytes > 10.000/uL, n (%) | 6.3 ± 1.4 | 7.4 ± 1.3 | 6.5 ± 1.8 | 0.12 |
| TroponinI > 0.07 ng/mL, n (%) | 19.6 ± 4.3 | 17.7 ± 3.5 | 23.3 ± 5.3 | 0.08 |
| Diseases, n (%) | ||||
| Hypertension | 0 | 0 | 0 | 0.98 |
| Hyper-lipidemia | 0 | 0 | 0 | 0.97 |
| Diabetes | 0 | 0 | 0 | 0.97 |
| Renal failure (MDRD < 40 mL/min/1.73 m2) | 0 | 0 | 0 | 0.97 |
| History of smoking | 1 (14) | 1 (10) | 2 (20) | 0.32 |
| Medication, n (%) | ||||
| ACE-I i/lub ARB | 7 (100) | 10 (100) | 10 (100) | 0.98 |
| Beta-blockers | 2 (29) | 7 (70) | 10 (100) | 0.05 * |
| Aldosteron antagonist | 0 | 3 (30) | 7 (70) | 0.08 |
| Loop diuretics | 0 | 7 (70) | 10 (100) | 0.04 |
| Statin | 0 | 7 (70) | 10 (100) | 0.04 |
| Digoxin | 0 | 0 | 33 | 0.07 |
| EMB results, n (%) | ||||
| Acute myocarditis | 0 | 0 | 0 | 0.98 |
| Borderline myocarditis | 0 | 0 | 0 | 0.98 |
| No myocarditis | 7 (100) | 10 (100) | 10 (100) | 0.98 |
| Detection of viral genome | ||||
| PB19 [no of copy] | 0 | 0 | 6670 | 0.001 ** |
| Group III Patients with LVEF ≥ 50%, PB19V Negative (Control) n = 7 | Group II Patients with DCM, LVEF < 50% PB19V Negative n = 10 | Group III Patients with DCM, LVEF < 50% PB19V Positive n = 10 | p | |
|---|---|---|---|---|
| Blurred structure of mitochondria [median (IQR)] | 1(1;1) | 1 (1;1) | 3 (2;3) | 0.0001 * |
| Increased electron density of mitochondrial matrix [median (IQR)] | 0 (0;0) | 0 (0;0) | 1 (1;1) | 0.01 * |
| Disrupted outer mitochondrial membrane [median (IQR)] | 1(1;1) | 1 (1;2) | 2 (1;2) | 0.001 ** |
| Loss of mitochondrial cristae [median (IQR)] | 1 (1;1) | 1 (1;2) | 1 (1;1) | 0.01 *** |
| Swelling of mitochondria [median (IQR)] | 0 (0;0) | 1 (1;1) | 2 (2;3) | 0.001 ** |
| Autophagy of mitochondria [median (IQR)] | 0 (0;1) | 1 (1;2) | 2 (2;3) | 0.0001 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pawlak, A.; Gewartowska, M.; Przybylski, M.; Kuffner, M.; Wiligórska, D.; Gil, R.; Król, Z.; Frontczak-Baniewicz, M. Ultrastructural Changes in Mitochondria in Patients with Dilated Cardiomyopathy and Parvovirus B19 Detected in Heart Tissue without Myocarditis. J. Pers. Med. 2022, 12, 177. https://doi.org/10.3390/jpm12020177
Pawlak A, Gewartowska M, Przybylski M, Kuffner M, Wiligórska D, Gil R, Król Z, Frontczak-Baniewicz M. Ultrastructural Changes in Mitochondria in Patients with Dilated Cardiomyopathy and Parvovirus B19 Detected in Heart Tissue without Myocarditis. Journal of Personalized Medicine. 2022; 12(2):177. https://doi.org/10.3390/jpm12020177
Chicago/Turabian StylePawlak, Agnieszka, Magdalena Gewartowska, Maciej Przybylski, Mateusz Kuffner, Diana Wiligórska, Robert Gil, Zbigniew Król, and Malgorzata Frontczak-Baniewicz. 2022. "Ultrastructural Changes in Mitochondria in Patients with Dilated Cardiomyopathy and Parvovirus B19 Detected in Heart Tissue without Myocarditis" Journal of Personalized Medicine 12, no. 2: 177. https://doi.org/10.3390/jpm12020177