
Cryptic Diversity in Paramecium multimicronucleatum Revealed with a Polyphasic Approach

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, Strain Choice, and Culture Maintenance
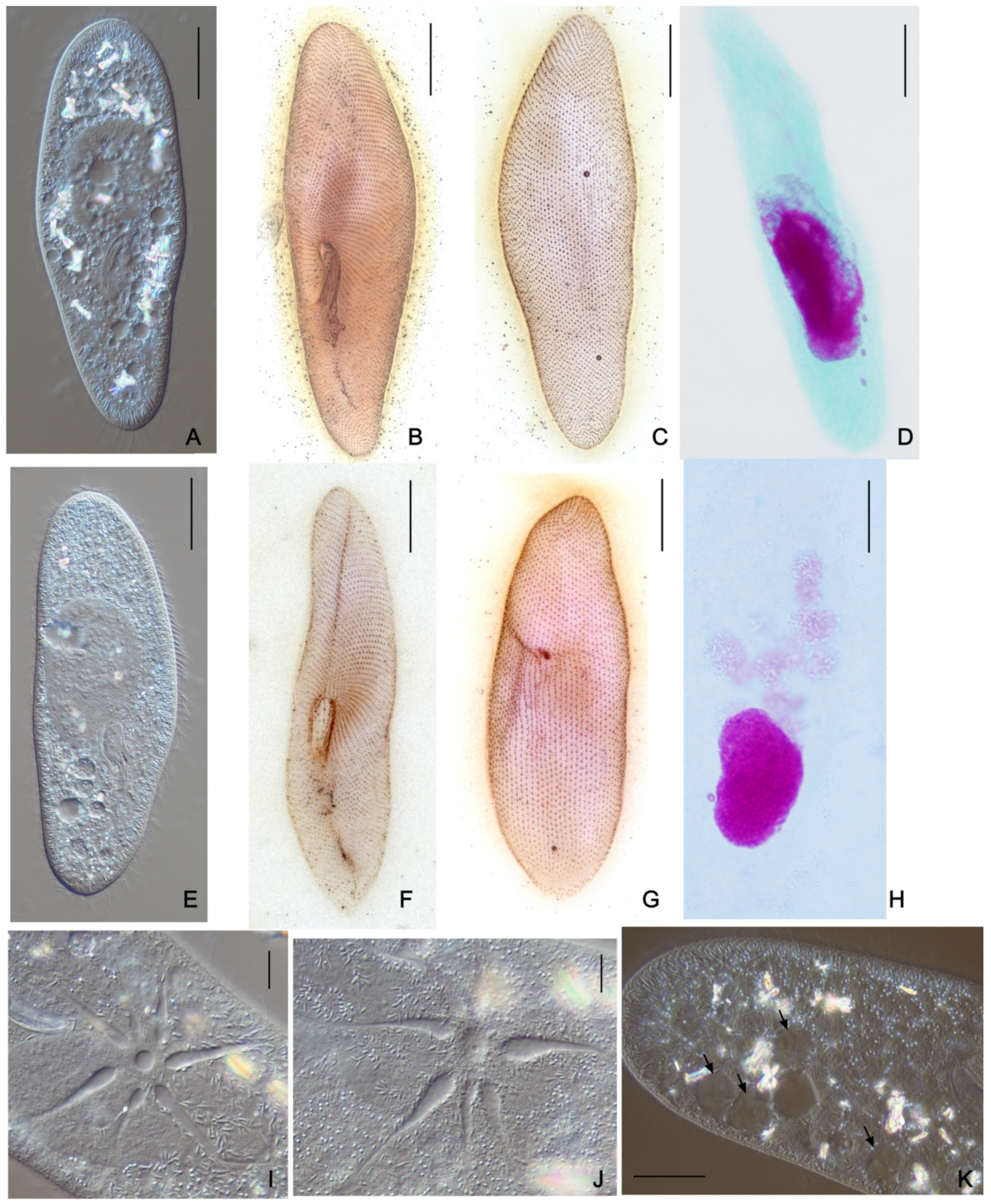
2.2. DIC Microscopy and Stainings
2.3. Molecular Identification of Paramecium Strains
2.4. Fluorescence In Situ Hybridization Assay
2.5. Molecular Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Index | Geographic Origin and Year of Collection | Biotope | Morphological and Physiological Characteristics | NCBI Accession Numbers | ||
|---|---|---|---|---|---|---|---|
| COI | 18S rDNA | ITS1-5.8S-ITS2 | |||||
| Paramecium multimicronucleatum | CyP5-3 | Cyprus, Paralimni, 2016 | ditch, 2‰ | CL: 130.7 ± 10.9 µm; CW: 30.9 ± 4.1 µm; ML: 55.7 ± 13.8 µm; MW: 20.2 ± 3 µm; NM: 1 or 3; DMIC: 2.09 ± 0.34 µm; vesicular MIC; MIC location: freely in cytoplasm; NCVC: 6–7; NCR: 74 ± 9 | OM401905 | OM200731 | |
| ID1-13 | India, Delhi, 2018 | pond | OM401906 | nd | nd | ||
| MSA-5 | Malta, San Anton Gardens, 2013 | stone bowl | OM401907 | OM200732 | |||
| Ns2-16 | Russia, Novosibirsk, 2002 | creek | OM401908 | OM200756 | [12] | ||
| Vv171-1 | Russia, Vladivostok, 2007 | pond | OM221497 | OM200757 | [12] | ||
| Thk-16 | Thailand, Phi Phi don, 2014 | creek | OM401909 | OM200733 | |||
| R51-6 | Mexico, Requena Lake, 2019 | lake | CL: 149.3 ± 11 µm; CW: 27.8 ± 4.3 µm; ML: 51.1 ± 8.1 µm; MW: 16 ± 2.6 µm; NM: 1–3; DMIC: 4.31 ± 0.3 µm; vesicular MIC; MIC location: near MAC; NCVC: 5–8; NCR: 73 ± 8; Intrastrain conjugation (selfing) observed in some strains | OM401910 | OM200734 | ||
| SMM80-11 | Mexico, San Miguel Almaya, 2019 | lake | OM401911 | nd | nd | ||
| ChP10-2 | Mexico City, Chapultepec, 2019 | lake in the city park | OM401912 | OM200735 | |||
| K4-2 | Mexico City, Cantera Oriente, 2019 | pond | [19] | OM200736 | |||
| L72-1 | Mexico, Lerma, 2019 | marsh | [19] | OM200737 | |||
| MB2-5 | Moldova, Bendery, 1996 | river | [12] | OM200758 | [12] | ||
| Or4-3 | Russia, Orenburg, 2015 | wastewater stream | OM221498 | OM200738 | |||
| OmN-1 | USA, Omaha, NE, 2018 | Missouri river | OM401913 | OM200739 | |||
| SK6-3 | Mexico, Sian Kaan, 2019 | ditch | OM401914 | OM200740 | |||
| LB-2 | Mexico, Bacalar, 2019 | freshwater lagoon | OM401915 | OM200741 | |||
| ChP3-4 | Mexico City, Chapultepec, 2019 | lake in the city park | CL: 180.3 ± 13.9 µm; CW: 36.2 ± 3.9 µm; ML: 61 ± 7.5 µm; MW: 19 ± 2.7 µm; NM: 2; DMIC: 3.49 ± 0.46 µm; vesicular MIC; MIC location: freely in cytoplasm; NCVC: 6–8; NCR: 72 ± 7 | OM401916 | nd | nd | |
| E59-1B | Mexico, Endoh Lake, 2019 | lake | OM401917 | nd | nd | ||
| IP2-1 | Italy, Pisa, 2016 | channel | OM401918 | nd | nd | ||
| ChP5-3 | Mexico City, Chapultepec, 2019 | lake in the city park | [19] | OM200742 | |||
| Paramecium fokini n. sp. | SMM81-1 | Mexico, San Miguel Almaya, 2019 | lake | CL: 155.6 ± 13.9 µm; CW: 27.6 ± 4,8 µm; ML: 54.6 ± 7.7 µm; MW: 16.4 ± 2.4 µm; NM: 1–3; DMIC: 4.1 ± 0.66 µm; vesicular MIC; MIC location: near MAC; NCVC: 6–8; NCR: 67 ± 8; Intrastrain conjugation (selfing) observed in some strains | [19] | OM200743 | |
| T42-1 | Mexico City, Tlahuac, 2019 | channel | [19] | OM200744 | |||
| AB9-8 | USA, Boston, 1994 | pond | OM401919 | OM200759 | [12] | ||
| CyL3-21 | Cyprus, Larnaka, Aliki region, 2010 | ditch, 1–2‰ | OM401920 | OM200745 | |||
| PP-2 | Russia, Pskov region, 2012 | ditch | OM401921 | OM200746 | |||
| PL4-1 | Portugal, Lisbon, 2019 | concrete basin | OM401922 | OM200747 | |||
| OP13 | Russia, Saint Petersburg, 1992 | city pond | OM401923 | OM200760 | [12] | ||
| FCB10-1 | France, Corsica, Bastia, 2015 | stream | OM221499 | OM200748 | |||
| Paramecium lynni n. sp. | ShKm41 | Russia, Kemerovo region, Shestakovo, 2008 | river | CL: 133.7 ± 13.1 µm; CW: 31.9 ± 5.1 µm; ML: 38.6 ± 6.2 µm; MW: 15.8 ± 2.8 µm; NM: 1–3; DMIC: 3.55 ± 0.46 µm; ‘fried egg’ MIC; MIC location: freely in cytoplasm; NCVC: 6–8; NCR: 64 ± 8 | OM401924 | OM200749 | |
| HSG3-10 | Russia, Saint Petersburg region, Peterhof, 2017 | ditch | OM401925 | OM200750 | |||
| SD11-9 | Russia, Saint Petersburg region, Sestroretsk, 2017 | pond | OM401926 | OM200751 | |||
| PO16-1 | Russia, Pskov region, Ostrov, 2019 | pond | OM401927 | OM200752 | |||
| SP-1 | Russia, Saint Petersburg region, Peterhof, 2019 | pond | OM401928 | OM200753 | |||
| Paramecium caudatum | Or4-4 | Russia, Orenburg, 2015 | wastewater stream | Cell size about 200 µm; single large compact MIC adjacent to MAC; 5–8 (more often 7) canals of CV; 1 pore per CV * | OM401929 | OM200754 | |
| K5-2 | Mexico City, Cantera Oriente, 2019 | pond | [19] | OM200755 | |||
3. Results
3.1. Identification of the New Species Distant from Paramecium multimicronucleatum
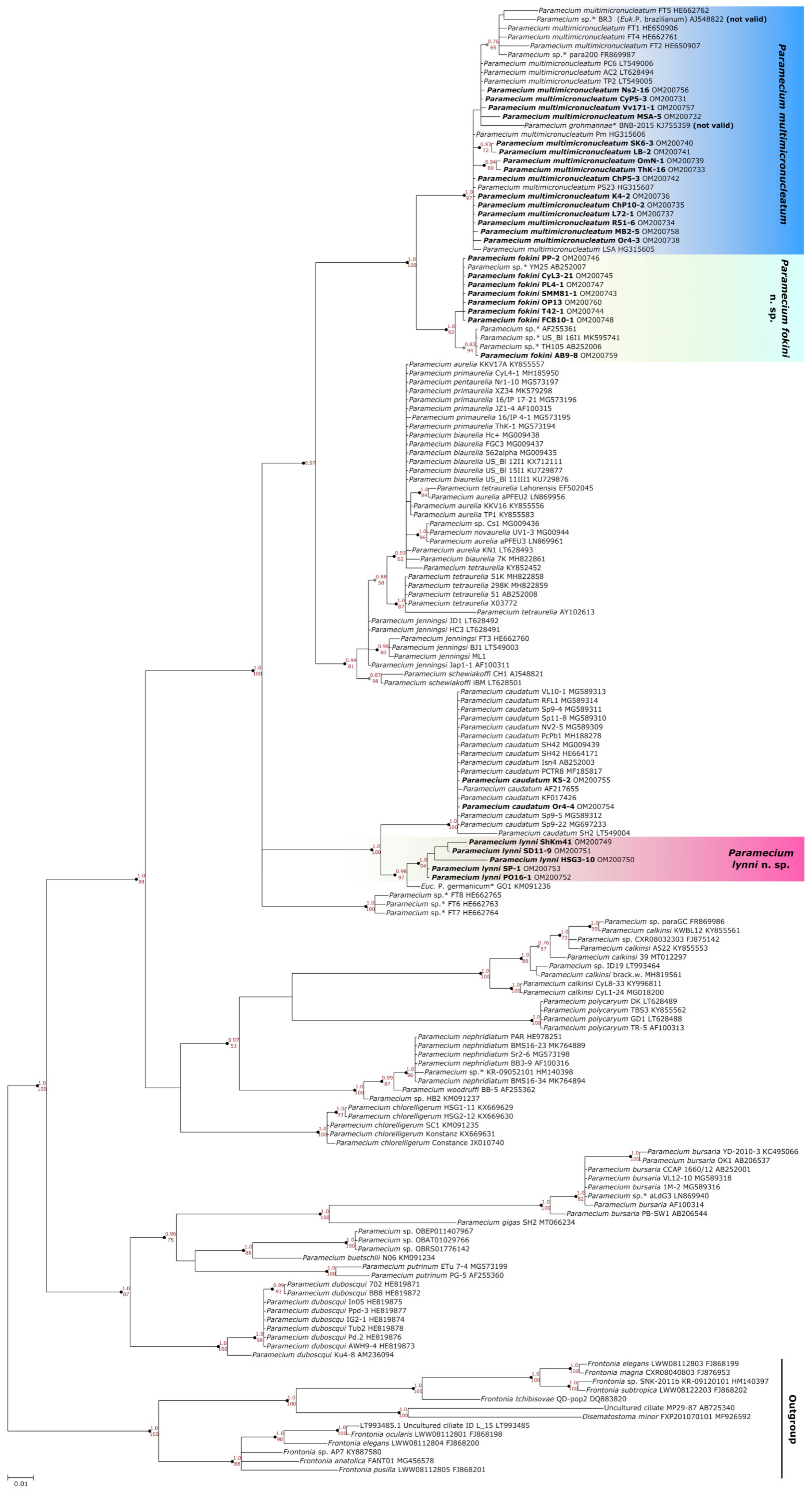
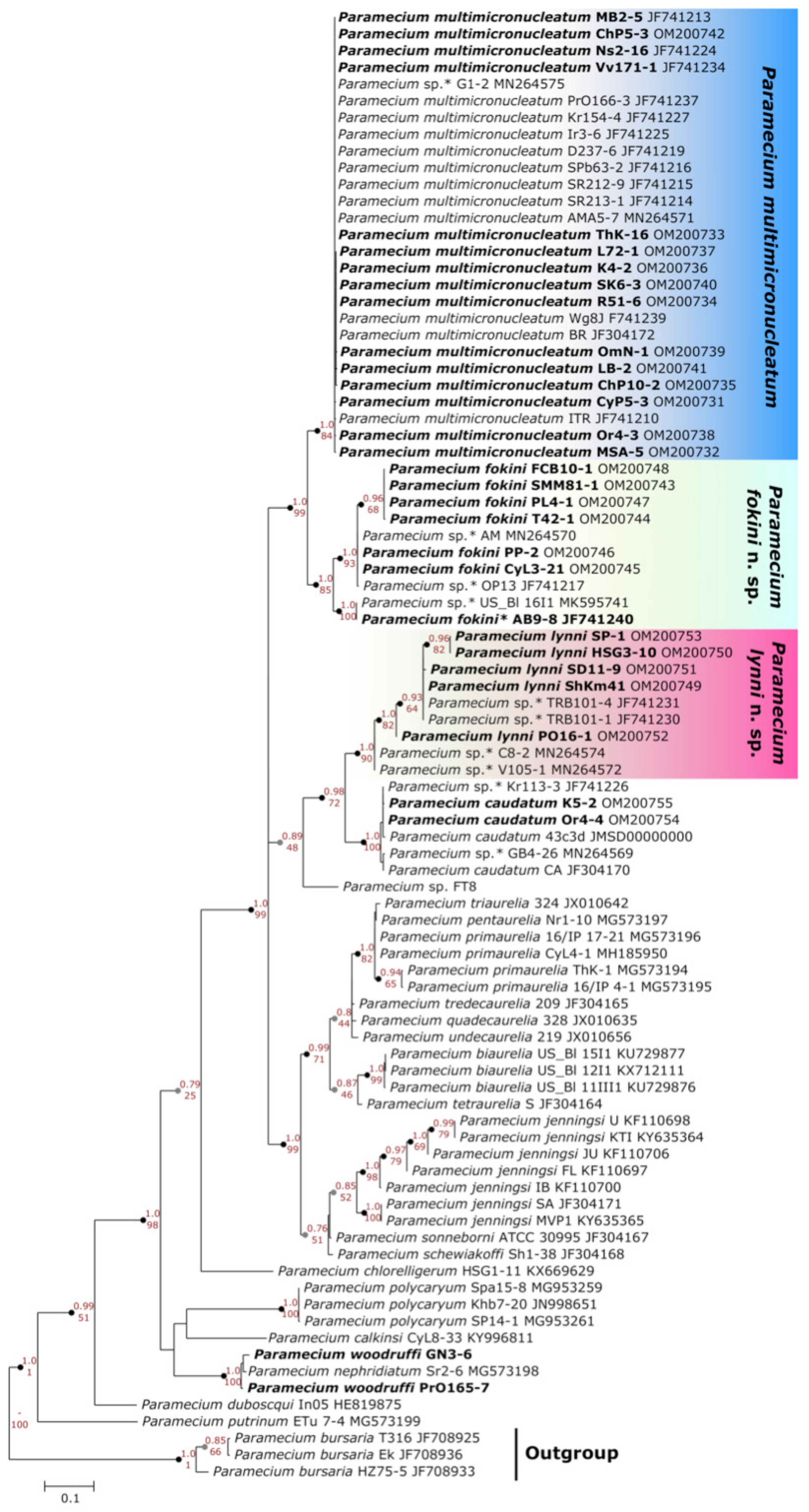
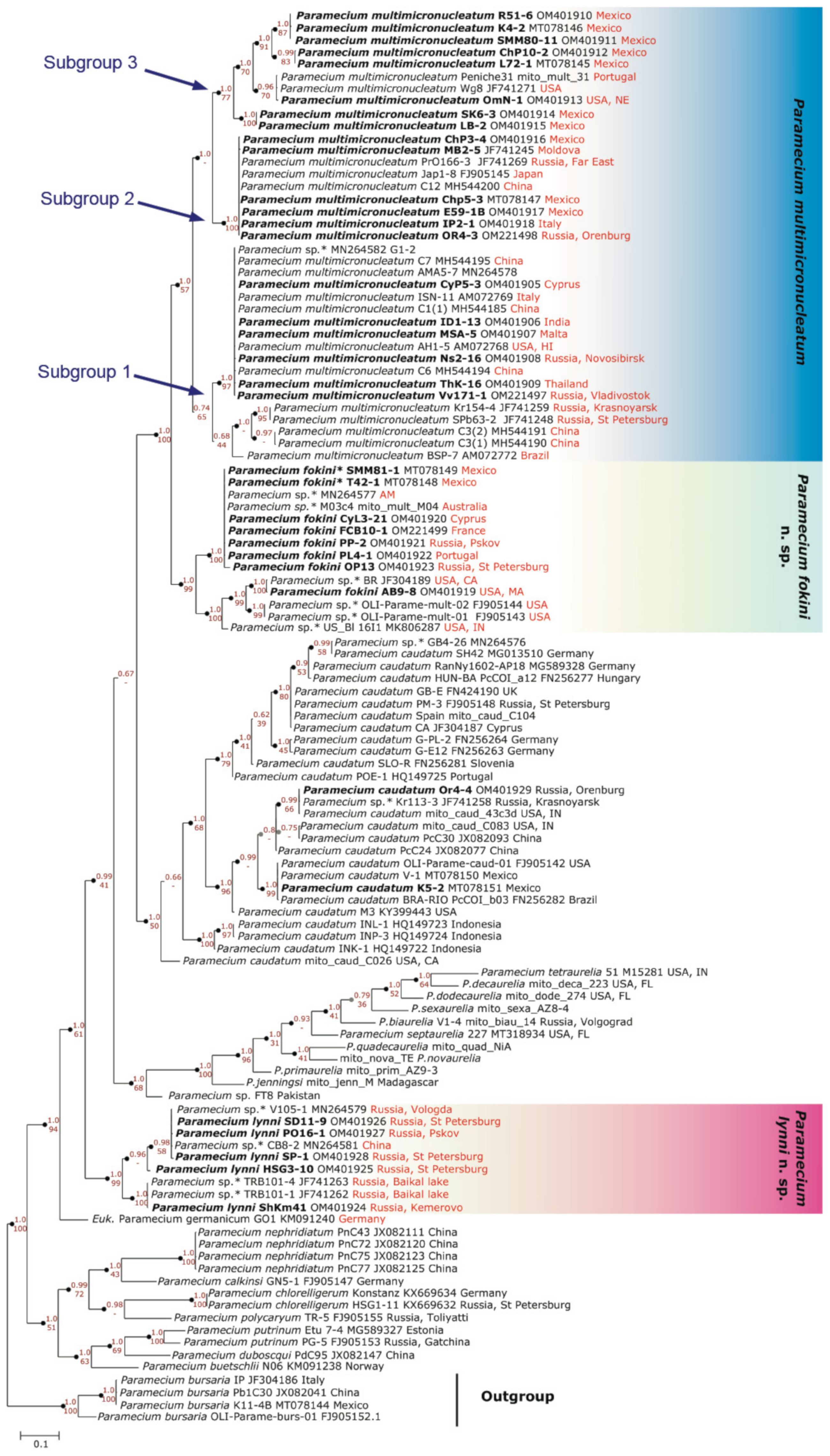
3.2. Two Groups Revealed within Paramecium multimicronucleatum Cluster
3.2.1. Molecular Methods Suggest Two Cryptic Species within P. multimicronucleatum
3.2.2. Morphological Variability within Paramecium multimicronucleatum
4. Discussion
4.1. Characteristics of Micronuclei and Paramecium Systematics
4.2. Dubious Paramecium Species
4.3. Paramecium lynni n. sp., the “Stealth Species”
4.4. Paramecium fokini n. sp., the Cryptic Species within Paramecium multimicronucleatum Clade
4.5. New Insights into Biogeography of Paramecium
4.6. The New Paramecium Species Formal Descriptions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Landis, W.G. Ecology. In Paramecium; Görtz, H.D., Ed.; Springer: Berlin/Heidelberg, Germany, 1988; pp. 419–437. [Google Scholar]
- Sonneborn, T.M. Paramecium aurelia. In Handbook of Genetics; King, R.C., Ed.; Plenum: New York, NY, USA, 1974; Volume 2, pp. 469–594. [Google Scholar]
- Beisson, J.; Bétermier, M.; Bre, M.H.; Cohen, J.; Duharcourt, S.; Duret, L.; Kung, C.; Malinsky, S.; Meyer, E.; Preer, J.R.; et al. Maintaining clonal Paramecium tetraurelia cell lines of controlled age through daily reisolation. Cold Spring Harb. Protoc. 2010, 2010, pdb-prot5361. [Google Scholar] [CrossRef] [PubMed]
- Wichterman, R. The Biology of Paramecium, 2nd ed.; Plenum Press: New York, NY, USA; London, UK, 1986; 599p. [Google Scholar]
- Sonneborn, T.M. The Paramecium aurelia complex of fourteen sibling species. Trans. Am. Microsc. Soc. 1975, 94, 155–178. [Google Scholar] [CrossRef]
- Fokin, S.I.; Stoeck, T.; Schmidt, H.J. Paramecium duboscqui Chatton, Brachon, 1933. Distribution, ecology and taxonomy. Eur. J. Protistol. 1999, 35, 161–167. [Google Scholar] [CrossRef]
- Fokin, S.I.; Stoeck, T.; Schmidt, H.J. Rediscovery of Paramecium nephridiatum Gelei, 1925 and its characteristics. J. Eukaryot. Microbiol. 1999, 46, 416–426. [Google Scholar] [CrossRef]
- Kreutz, M.; Stoeck, T.; Foissner, W. Morphological and molecular characterization of Paramecium (Viridoparamecium nov. subgen.) chlorelligerum Kahl (Ciliophora). J. Eukaryot. Microbiol. 2012, 59, 548–563. [Google Scholar] [CrossRef] [Green Version]
- Serra, V.; Fokin, S.I.; Gammuto, L.; Nitla, V.; Castelli, M.; Basuri, C.K.; Satyaveni, A.; Sandeep, B.V.; Modeo, L.; Petroni, G. Phylogeny of Neobursaridium reshapes the systematics of Paramecium (Oligohymenophorea, Ciliophora). Zool. Scr. 2021, 50, 241–268. [Google Scholar] [CrossRef]
- Fokin, S.I.; Przyboś, E.; Chivilev, S.M.; Beier, C.L. Morphological and molecular investigations of Paramecium schewiakoffi sp. nov. (Ciliophora, Oligohymenophorea) and current status of distribution and taxonomy of Paramecium spp. Eur. J. Protistol. 2004, 40, 225–243. [Google Scholar] [CrossRef]
- Krenek, S.; Berendonk, T.U.; Fokin, S.I. New Paramecium (Ciliophora, Oligohymenophorea) congeners shape our view on its biodiversity. Org. Divers. Evol. 2015, 15, 215–233. [Google Scholar] [CrossRef]
- Tarcz, S.; Potekhin, A.; Rautian, M.; Przyboś, E. Variation in ribosomal and mitochondrial DNA sequences demonstrates the existence of intraspecific groups in Paramecium multimicronucleatum (Ciliophora, Oligohymenophorea). Mol. Phylogenet. Evol. 2012, 63, 500–509. [Google Scholar] [CrossRef]
- Tarcz, S.; Rautian, M.; Potekhin, A.; Sawka, N.; Beliavskaya, A.; Kiselev, A.; Nekrasova, I.; Przyboś, E. Paramecium putrinum (Ciliophora, Protozoa): The first insight into the variation of two DNA fragments—Molecular support for the existence of cryptic species. Mol. Phylogenet. Evol. 2014, 73, 140–145. [Google Scholar] [CrossRef]
- Greczek-Stachura, M.; Potekhin, A.; Przyboś, E.; Rautian, M.; Skoblo, I.; Tarcz, S. Identification of Paramecium bursaria syngens through molecular markers—Comparative analysis of three loci in the nuclear and mitochondrial DNA. Protist 2012, 163, 671–685. [Google Scholar] [CrossRef]
- Jankowski, A.V. Proposed classification of Paramecium genus Hill, 1752 (Ciliophora). Zoologicheskii Zhurnal 1969, 48, 30–39, (In Russian with English Summary). [Google Scholar]
- Fokin, S.I. Paramecium genus: Biodiversity, some morphological features and the key to the main morphospecies discrimination. Protistology 2010, 6, 227–235. [Google Scholar]
- Przyboś, E.; Tarcz, S.; Potekhin, A.; Rautian, M.; Prajer, M. A two-locus molecular characterization of Paramecium calkinsi. Protist 2012, 163, 263–273. [Google Scholar] [CrossRef]
- Aufderheide, K.J.; Daggett, P.-M.; Nerad, T.A. Paramecium sonneborni n. sp., a new member of the Paramecium aurelia species complex. J. Protozool. 1983, 30, 128–131. [Google Scholar] [CrossRef]
- Potekhin, A.; Mayen-Estrada, R. Paramecium diversity and a new member of the Paramecium aurelia species complex described from Mexico. Diversity 2020, 12, 197. [Google Scholar] [CrossRef]
- Sawka-Gadek, N.; Potekhin, A.; Singh, D.P.; Grevtseva, I.; Arnaiz, O.; Penel, S.; Sperling, L.; Tarcz, S.; Duret, L.; Nekrasova, I.; et al. Evolutionary plasticity of mating-type determination mechanisms in Paramecium aurelia sibling species. Genome Biol. Evol. 2021, 13, evaa258. [Google Scholar] [CrossRef]
- Fokin, S.I. Revision of the Paramecium Müller, 1773 Genus: Comparative-Biological Analysis, Systematics and Phylogenetical Relations. Ph.D. Thesis, Saint Petersburg State University, Saint Petersburg, Russia, 2002. [Google Scholar]
- Paiva, T.S.; Borges, B.; Harada, M.L.; da Silva-Neto, I.D. Description and molecular phylogeny of Paramecium grohmannae sp. nov. (Ciliophora, Peniculida) from a wastewater treatment plant in Brazil. Revista Brasileira Zoociências 2016, 17, 7–19. [Google Scholar]
- Shakoori, F.R.; Tasneem, F.; Al-Ghanim, K.; Mahboob, S.; Al-Misned, F.; Jahan, N.; Shakoori, A.R. Variability in secondary structure of 18S ribosomal RNA as topological marker for identification of Paramecium species. J. Cell. Biochem. 2014, 115, 2077–2088. [Google Scholar] [CrossRef]
- De Souza, B.A.; Dias, R.J.P.; Senra, M.V.X. Intrageneric evolutionary timing and hidden genetic diversity of Paramecium lineages (Ciliophora: Oligohymenophorea). Syst. Biodivers. 2020, 18, 662–674. [Google Scholar] [CrossRef]
- Powers, J.H.; Mitchell, C. A new species of Paramecium (P. multimicronucleata) experimentally determined. Biol. Bull. 1910, 19, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Fokin, S.I. Morphological diversity of the micronuclei in Paramecium. Archiv Protistenkunde 1997, 148, 375–387. [Google Scholar] [CrossRef]
- Fokin, S.I.; Chivilev, S.M. Paramecium Morphometric analysis and taxonomy. Acta Protozool. 2000, 39, 1–14. [Google Scholar]
- Yan, Y.; Jiang, L.; Aufderheide, K.; Wright, G.A.; Terekhov, A.; Costa, L.; Qin, K.; McCleery, W.T.; Fellenstein, J.J.; Ustione, A.; et al. A microfluidic-enabled mechanical microcompressor for the immobilization of live single- and multi-cellular specimens. Microsc. Microanal. 2014, 20, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Zharikov, V.V. Cadastre of Free-Living Ciliates of Volga Reservoirs; IEVB RAS: Tolyatti, Russia, 1996; pp. 1–76. (In Russian) [Google Scholar]
- Foissner, W. An update of ‘basic light and scanning electron microscopic methods for taxonomic studies of ciliated protozoa. Int. J. Syst. Evol. Microbiol. 2014, 64, 271–292. [Google Scholar] [CrossRef] [Green Version]
- Boscaro, V.; Fokin, S.I.; Verni, F.; Petroni, G. Survey of Paramecium duboscqui using three markers and assessment of the molecular variability in the genus Paramecium. Mol. Phylogenet. Evol. 2012, 65, 1004–1013. [Google Scholar] [CrossRef]
- Strüder-Kypke, M.C.; Lynn, D.H. Comparative analysis of the mitochondrial cytochrome c oxidase subunit I (COI) gene in ciliates (Alveolata, Ciliophora) and evaluation of its suitability as a biodiversity marker. Syst. Biodivers. 2010, 8, 131–148. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; 2100p. [Google Scholar]
- Manz, W.; Amann, R.; Ludwig, W.; Wagner, M.; Schleifer, K.-H. Phylogenetic oligodeoxynucleotide probes for the major subclasses of Proteobacteria: Problems and solutions. Syst. Appl. Microbiol. 1992, 15, 593–600. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, F. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, F.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Serra, F.; Bork, P. ETE 3: Reconstruction, analysis, and visualization of phylogenomic data. Mol. Biol. Evol. 2016, 33, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Strüder-Kypke, M.C.; Wright, A.G.; Fokin, S.I.; Lynn, D. Phylogenetic relationships of the genus Paramecium inferred from small subunit rRNA gene sequences. Mol. Phylogenet. Evol. 2000, 14, 122–130. [Google Scholar] [CrossRef]
- Weisse, T. Distribution and diversity of aquatic protists: An evolutionary and ecological perspective. Biodivers. Conserv. 2007, 17, 243–259. [Google Scholar] [CrossRef]
- Santoferrara, L.F.; Grattepanche, J.D.; Katz, L.A.; McManus, G.B. Patterns and processes in microbial biogeography: Do molecules and morphologies give the same answers? ISME J. 2016, 10, 1779–1790. [Google Scholar] [CrossRef]
- Foissner, W.; Moon-van der Staay, S.Y.; van der Staay, G.W.M.; Hackstein, J.H.P.; Krautgartner, W.; Berger, H. Reconciling classical and molecular phylogenies in the stichotrichines (Ciliophora, Spirotrichea), including new sequences from some rare species. Eur. J. Protistol. 2004, 40, 265–281. [Google Scholar] [CrossRef]
- Katz, L.A.; Mcmanus, G.B.; Snoeyenbos-West, O.; Griffin, A. Reframing the ‘Everything is everywhere’ debate: Evidence for high gene flow and diversity in ciliate morphospecies. Aquat. Microb. Ecol. 2005, 41, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Vd’ačný, P. Integrative taxonomy of ciliates: Assessment of molecular phylogenetic content and morphological homology testing. Eur. J. Protistol. 2017, 61, 388–398. [Google Scholar] [CrossRef]
- Vd’ačný, P.; Rajter, L. Reconciling morphological and molecular classification of predatory ciliates: Evolutionary taxonomy of dileptids (Ciliophora, Litostomatea, Rhynchostomatia). Mol. Phylogenet. Evol. 2015, 90, 112–128. [Google Scholar] [CrossRef]
- Zhao, Y.; Yi, Z.; Warren, A.; Song, W. Species delimitation for the molecular taxonomy and ecology of the widely distributed microbial eukaryote genus Euplotes (Alveolata, Ciliophora). Proc. Biol. Sci. 2018, 285, 20172159. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.Q.; Wang, G.Y.; Xiong, J.; Yang, W.T.; Sun, Z.Y.; Feng, J.M.; Warren, A.; Miao, W. Insights into the origin and evolution of Peritrichia (Oligohymenophorea, Ciliophora) based on analyses of morphology and phylogenomics. Mol. Phylogenet. Evol. 2019, 132, 25–35. [Google Scholar] [CrossRef]
- Cedrola, F.; Senra, M.V.X.; Rossi, M.F.; Fregulia, P.; D’Agosto, M.; Dias, R.J.P. Trichostomatid ciliates (Alveolata, Ciliophora, Trichostomatia) systematics and diversity: Past, present, and future. Front. Microbiol. 2020, 10, 2967. [Google Scholar] [CrossRef] [Green Version]
- Melekhin, M.; Lebedeva, N.; Lanzoni, O.; Nekrasova, I.; Nitla, V.M.; Petroni, G.; Fokin, S.I.; Potekhin, A. Cryptic species diversity in Paramecium (Ciliophora): More search, more find. In Proceedings of the 15th International Congress of Protistology, Prague, Czech Republic, 30 July–4 August 2017; p. 343. [Google Scholar]
- Przyboś, E.; Tarcz, S. Paramecium jenningsi complex: Existence of three cryptic species confirmed by multi-locus analysis and strain crosses. Syst. Biodivers. 2016, 14, 140–154. [Google Scholar] [CrossRef]
- Przybós, E.; Tarcz, S.; Rautian, M.; Lebedeva, N. The first European stand of Paramecium sonneborni (P. aurelia complex), a species known only from North America (Texas, USA). Eur. J. Protistol. 2014, 50, 236–247. [Google Scholar] [CrossRef]
- Foissner, W.; Berger, W.; Schaumburg, J. Identification and Ecology of Limnetic Plankton Ciliates. Informationsberichte des Bayerischen Landesamtes für Wasserwirtschaft 1999, 3, 793. [Google Scholar]
- Grimm, D.; Ludwig, W.; Brandt, B.C.; Michel, R.; Schleifer, K.-H.; Hacker, J.; Steinert, M. Development of 18S rRNA-targeted oligonucleotide probes for specific detection of Hartmannella and Naegleria in Legionella–positive environmental samples. Syst. Appl. Microbiol. 2001, 24, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.L.; Caron, D.A.; DeLong, E.F. Development and field application of a quantitative method for examining natural assemblages of protists with oligonucleotide probes. Appl. Environ. Microbiol. 1996, 62, 1416–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkuma, M.; Ohtoko, K.; Iida, T.; Tokura, M.; Moriya, S.; Usami, R.; Horikoshi, K.; Kudo, T. Phylogenetic identification of hypermastigotes, Pseudotrichonympha, Spirotrichonympha, Holomastigotoides, and parabasalian symbionts in the hindgut of termites. J. Eukaryot. Microbiol. 2000, 47, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Petroni, G.; Rosati, G.; Vannini, C.; Modeo, L.; Dini, F.; Verni, F. In Situ identification by fluorescently labeled oligonucleotide probes of morphologically similar, closely related ciliate species. Microb. Ecol. 2003, 45, 156–162. [Google Scholar] [CrossRef]
- Schmidt, S.L.; Bernhard, D.; Schlegel, M.; Fried, J. Fluorescence in situ hybridization with specific oligonucleotide rRNA probes distinguishes the sibling species Stylonychia lemnae and Stylonychia mytilus (Ciliophora, Spirotrichea). Protist 2006, 157, 21–30. [Google Scholar] [CrossRef]
- Allen, S.L.; Rushford, D.L.; Nerad, T.A.; Lau, E.T. Intraspecies variability in the esterases and acidphosphatases of Paramecium jenningsi and Paramecium multimicronucleatum: Assignment of unidentified paramecia, comparison with the Paramecium aurelia complex. J. Protozool. 1983, 30, 155–163. [Google Scholar] [CrossRef]
- Bomford, S. The syngenes of Paramecium bursaria: New mating types and intersyngenic mating reactions. J. Protozool. 1966, 13, 497–501. [Google Scholar] [CrossRef]
- Potekhin, A.; Przyboś, E.; Nekrasova, I.; Yashchenko, V.; Rautian, M. Species of the Paramecium aurelia complex in Russia: New stands and overall distribution. Folia Biol. 2010, 58, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Tarcz, S.; Sawka-Gądek, N.; Przyboś, E. Worldwide sampling reveals low genetic variability in populations of the freshwater ciliate Paramecium biaurelia (P. aurelia species complex, Ciliophora, Protozoa). Org. Divers. Evol. 2018, 18, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Przyboś, E.; Prajer, M. New stands of species of the Paramecium aurelia complex; Is the occurrence of particular species limited by temperature barriers? Folia Biol. 2015, 63, 215–220. [Google Scholar] [CrossRef]
- Przyboś, E.; Tarcz, S.; Dusi, E. New Paramecium quadecaurelia strains (P. aurelia spp. complex, Ciliophora) identified by molecular markers (rDNA and mtDNA). Eur. J. Protistol. 2013, 49, 477–486. [Google Scholar] [CrossRef]
- Barth, D.; Krenek, S.; Fokin, S.I.; Berendonk, T.U. Intraspecific genetic variation in Paramecium revealed by mitochondrial cytochrome C oxidase I sequences. J. Eukaryot. Microbiol. 2006, 53, 20–25. [Google Scholar] [CrossRef]
- Lu, X.; Gentekaki, E.; Xu, Y.; Huang, L.; Li, Y.; Lu, X.; Zhao, Y.; Lin, X.; Yi, Z. Intra-population genetic diversity and its effects on outlining genetic diversity of ciliate populations: Using Paramecium multimicronucleatum as an example. Eur. J. Protistol. 2019, 67, 142–150. [Google Scholar] [CrossRef]
- Yi, Z.; Strüder-Kypke, M.; Hu, X.; Lin, X.; Song, W. Sampling strategies for improving tree accuracy and phylogenetic analyses: A case study in ciliate protists, with notes on the genus Paramecium. Mol. Phylogenet. Evol. 2014, 71, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Beale, G.; Preer, J.R., Jr. Paramecium Genetics and Epigenetics; CRC Press: Boca Raton, FL, USA, 2008; pp. 1–216. [Google Scholar]
- Barnett, A. A circadian rhythm of mating type reversals in Paramecium multimicronucleatum, syngen 2, and its genetic control. Cell. Physiol. 1966, 67, 239–270. [Google Scholar] [CrossRef] [PubMed]
- Vishnyakov, A.; Rodionova, G. Motile intranuclear symbionts of ciliate Paramecium multimicronucleatum. In Endocytobiology VII; Wagner, E., Normann, J., Greppin, H., Hackstein, R.G., Kowallik, K.V., Shenk, H.E.A., Seckbach, J., Eds.; Geneva University Press: Genève, Switzerland, 1999; pp. 169–177. [Google Scholar]
- Mironov, T.; Sabaneyeva, E. A robust symbiotic relationship between the ciliate Paramecium multimicronucleatum and the bacterium Ca. Trichorickettsia mobilis. Front. Microbiol. 2020, 11, 603335. [Google Scholar] [CrossRef]
- Serra, V.; Fokin, S.I.; Castelli, M.; Basuri, C.K.; Nitla, V.; Verni, F.; Sandeep, B.V.; Kalavati, C.; Petroni, G. “Candidatus Gortzia shahrazadis”, a novel endosymbiont of Paramecium multimicronucleatum and a revision of the biogeographical distribution of Holospora-like bacteria. Front. Microbiol. 2016, 7, 1704. [Google Scholar] [CrossRef] [Green Version]
- Modeo, L.; Salvetti, A.; Rossi, L.; Castelli, M.; Szokoli, F.; Krenek, S.; Serra, V.; Sabaneyeva, E.; Di Giuseppe, G.; Fokin, S.I.; et al. “Candidatus Trichorickettsia mobilis”, a Rickettsiales bacterium, can be transiently transferred from the unicellular eukaryote Paramecium to the planarian Dugesia japonica. Peer J. 2020, 8, e8977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasneem, F.; Shakoori, F.R.; Ilyas, M.; Shahzad, N.; Potekhin, A.; Shakoori, A.R. Genetic diversity of Paramecium species on the basis of multiple loci analysis and ITS secondary structure models. J. Cell. Biochem. 2020, 121, 3837–3853. [Google Scholar] [CrossRef] [PubMed]
- Petroni, G.; Dini, F.; Verni, F.; Rosati, G. A molecular approach to the tangled intrageneric relationships underlying phylogeny in Euplotes (Ciliophora, Spirotrichea). Mol. Phylogenet. Evol. 2002, 22, 118–130. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Rosati, G.; Modeo, L.; Melai, M.; Petroni, G.; Verni, F. A multidisciplinary approach to describe protists: A morphological, ultrastructural, and molecular study on Peritromus kahli Villeneuve-Brachon, 1940 (Ciliophora, Heterotrichea). J. Eukaryot. Microbiol. 2004, 51, 49–59. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melekhin, M.; Yakovleva, Y.; Lebedeva, N.; Nekrasova, I.; Nikitashina, L.; Castelli, M.; Mayén-Estrada, R.; Romanovich, A.E.; Petroni, G.; Potekhin, A. Cryptic Diversity in Paramecium multimicronucleatum Revealed with a Polyphasic Approach. Microorganisms 2022, 10, 974. https://doi.org/10.3390/microorganisms10050974
Melekhin M, Yakovleva Y, Lebedeva N, Nekrasova I, Nikitashina L, Castelli M, Mayén-Estrada R, Romanovich AE, Petroni G, Potekhin A. Cryptic Diversity in Paramecium multimicronucleatum Revealed with a Polyphasic Approach. Microorganisms. 2022; 10(5):974. https://doi.org/10.3390/microorganisms10050974
Chicago/Turabian StyleMelekhin, Maksim, Yulia Yakovleva, Natalia Lebedeva, Irina Nekrasova, Liubov Nikitashina, Michele Castelli, Rosaura Mayén-Estrada, Anna E. Romanovich, Giulio Petroni, and Alexey Potekhin. 2022. "Cryptic Diversity in Paramecium multimicronucleatum Revealed with a Polyphasic Approach" Microorganisms 10, no. 5: 974. https://doi.org/10.3390/microorganisms10050974