
Octorhopalona saltatrix, a New Genus and Species (Hydrozoa, Anthoathecata) from Japanese Waters

1
Kuroshio Biological Research Foundation, 560 Nishidomari, Otsuki, Hata 788-0333, Japan
2
Enoshima Aquarium, Katasekaigan, Fujisawa 251-0035, Japan
3
Ibaraki Prefectural Oarai Aquarium, 8252-3 Isohamacho, Oarai, Higashi-ibaraki 311-1301, Japan
*
Author to whom correspondence should be addressed.
Animals 2022, 12(13), 1600; https://doi.org/10.3390/ani12131600
Submission received: 21 May 2022
/
Revised: 14 June 2022
/
Accepted: 15 June 2022
/
Published: 21 June 2022
(This article belongs to the Special Issue Ecology, Evolution, and Systematics of Medusozoa (Aka Jellyfish))
Abstract
:Simple Summary
In this study, we describe a new genus and species of hydromedusa belonging to the family Halimedusidae (Hydrozoa, Anthoathecata) that is found off Oarai, Sagami Bay, and Tosa Bay, Japan. This family comprises four species in three genera: Halimedusa, Tiaricodon, and Urashimea. The new genus and species can be distinguished from all other Halimedusidae species by genetic sequences and morphological characteristics. A comparative table of the primary diagnostic characteristics of the genus Halimedusa is provided. In addition, the diagnosis of Halimedusidae was modified. Jellyfish blooms cause serious problems in fishing, industry and public health. This paper contributed to the understanding of ecology and diversity of jellyfish on our planet.
Abstract
Approximately 300 species of cnidarian jellyfish have been reported in Japanese waters. However, many specimens remain unidentified. In this study, taxonomic investigations, including morphological observations and molecular 16S phylogenetic analyses, were conducted on unknown specimens collected off Oarai, Sagami Bay, and Tosa Bay, Japan. The specimens have the following morphological characteristics: distinct peaks in jelly above the base of the manubrium, a red band on the manubrium, and cylindrical marginal bulbs, each with an abaxial ocellus that is common to the family Halimedusidae. However, the specimens can be distinguished from other Halimedusidae species by their eight radial canals, eight tentacles with numerous stalked nematocyst knobs, and eight nematocyst tracks on the exumbrella. Moreover, molecular phylogenetic analyses revealed that the Kimura two-parameter distance between the specimens and other Halimedusa species was 0.066–0.099, which is considered to represent intergeneric variability. Based on this result, we described it as a new species and established a new genus for taxonomic stabilization. We also emended the diagnostic characters of the family Halimedusidae owing to the establishment of the new genus. Halimedusidae comprises five species in four genera. This paper provides taxonomic keys for the identification of species in the family Halimedusidae.
1. Introduction
The hydrozoan family Halimedusidae is a small group that includes four species from three genera: Halimedusa Bigelow, 1916; Tiaricodon Browne, 1902; and Urashimea Kishinouye, 1910 [1,2,3]. Halimedusidae species are distributed in the shallow waters of a range of tropical, subtropical, and mild-temperature localities in the Pacific and Atlantic Oceans [1,4,5,6]. The species have sexual, planktonic medusae, and asexual benthic polyps in their life cycle [1,6,7]. Free-swimming medusae are liberated by the budding of small solitary polyps.
Historically, the taxonomy of the family Halimedusidae has been unclear because of the limited differences in morphological characteristics among the genera. The first identified Halimedusidae species, Tiaricodon coeruleus Browne, 1902, was described by Browne (1902) and classified in the family Polyorchidae [4]. Kishinouye (1910) and Bigelow (1916) described Urashimea globosa Kishinouye, 1910 (in Cladonematidae) and Halimedusa typus Bigelow, 1916 (in Pandeidae) [8,9], but Urashimea was later moved into the family Pandeidae by Uchida and Nagao (1961) [7]. Bouillon (1995) removed Urashimea to the family Pandeidae [10]. Arai and Brinckmann-Voss (1980) erected the family Halimedusidae in the suborder Filifera [11], and H. typus was moved from Pandeidae. However, Mills (2000) moved Halimedusidae to the suborder Capitata within Anthoathecata based on new information about the morphology of both the medusa and polyp phases, including their life cycle and cnidomes [1]. Additionally, Mills moved the genera Tiaricodon and Urashimea from Polyorchidae to Halimedusidae.
Halimedusidae are characterized as follows: a medusa usually with a low gastric peduncle and distinct subumbrellar pockets in jelly above the manubrium base; manubrium cruciform with basal perradial lobes; mouth quadratic to cruciform with lips lined by nematocysts; four radial canals, either with four perradial marginal tentacles or four perradial marginal tentacles and four interradial groups of tentacles, all hollow; gonads either on the manubrium or on the manubrium and perradial lobes; no mesenteries; cylindrical marginal bulbs with abaxial ocelli (after Bouillion et al., 2006) [2]; small, solitary polyp with a perisarcal base and a protective perisarcal spine above the hydranth; 3–8 (generally 4) oral capitate tentacles, with a few scattered cnidocysts along their length; and single medusa buds, just below the tentacles [12].
To date, two described Halimedusidae species, Urashimea globosa and Tiaricodon orientalis Yamamoto and Toshino, 2021, have been reported in Japanese waters [3,13]. In this study, 16 specimens of an unidentified Anthoathecata species were collected off Oarai, Sagami Bay, and Tosa Bay, Japan. Our morphological and molecular phylogenetic analyses suggest that this Anthoathecata species should be regarded as a new genus and species within the Halimedusidae family.
2. Materials and Methods
2.1. Collection and Fixing
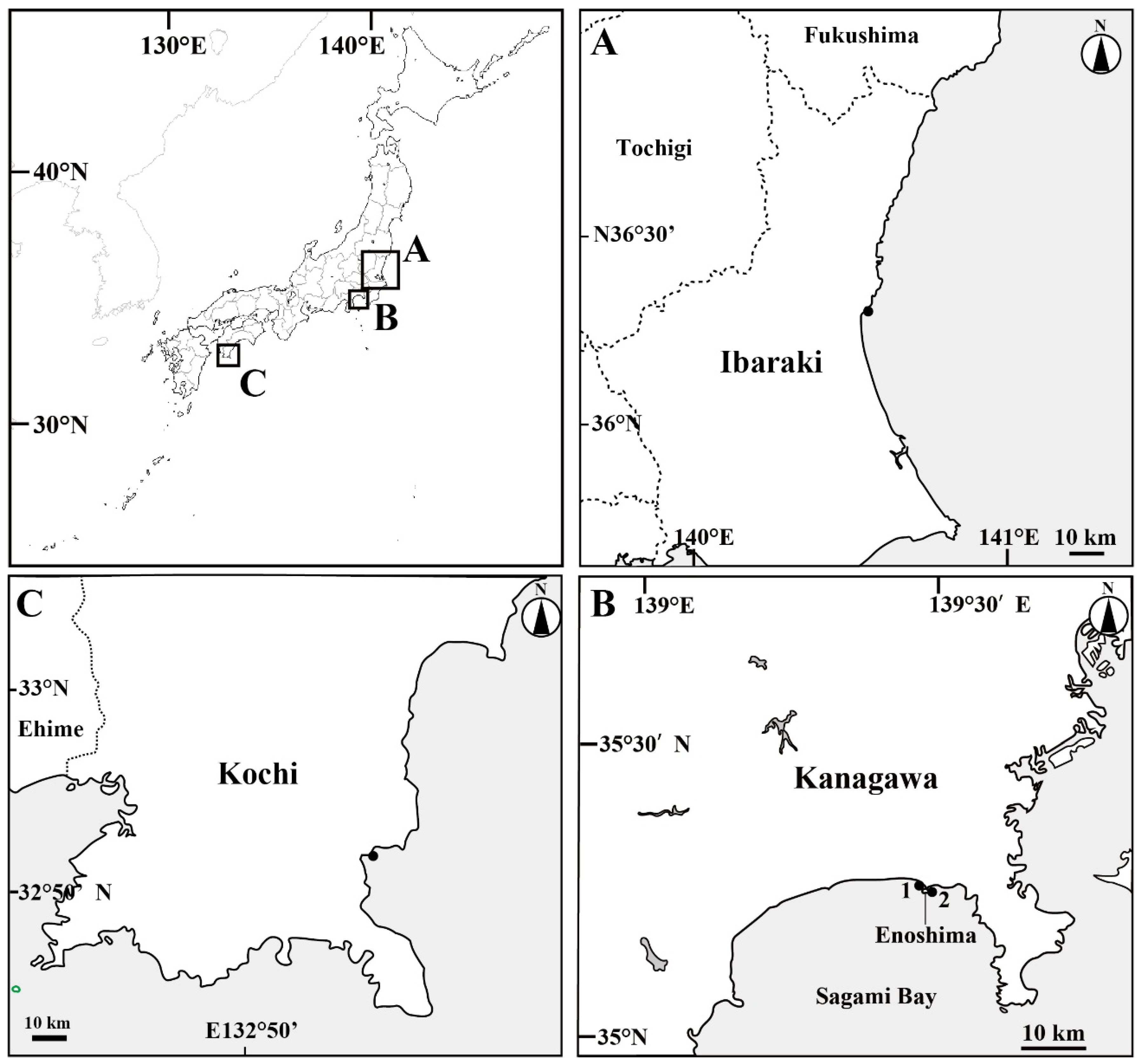
Sixteen unidentified medusae were collected near the water surface (within approximately 1 m) at Oarai Fishing Port, Higashi-Ibaraki, Ibaraki Prefecture; Katase Fishing Port, Enoshima, Fujisawa, Kanagawa Prefecture; and Shimonokae Fishing Port, Tosashimizu, Kochi Prefecture, Japan (Figure 1) between 15 December 2008, and 7 May 2021. The medusae were captured in either a dipper (diameter, 17 cm; volume, 2000 mL) or a dip net (mesh size, approximately 0.5 mm). Eight specimens were fixed in 3% formalin seawater and deposited in the Ibaraki Nature Museum, Ibaraki, Japan (INM); the National Museum of Nature and Science, Tsukuba, Japan (NSMT); and the Kuroshio Biological Research Foundation, Kochi, Japan (KBF). Seven specimens were preserved in 99.5% ethanol until molecular analysis. One specimen was used to determine the abundance of nematocyst types.
2.2. Molecular Phylogenetic Analysis
The 16S rDNA gene was used for the molecular phylogenetic analysis because it can effectively discriminate between species in Hydrozoa [14,15,16]. In this study, an approximately 600 bp fragment of mitochondrial 16S rDNA was used for phylogenetic analysis. Genomic DNA was extracted from the 99.5% ethanol-preserved tissue of cultured specimens using the DNeasy Blood and Tissue Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions. The 16S rDNA was PCR-amplified and sequenced with the forward and reverse primer pair TCGACTGTTTACCAAAAACATAGC and ACGGAATGAACTCAAATCATGTAAG [17], respectively, using the following PCR profile: an initial denaturation at 94 °C for 5 min; five cycles at 94 °C for 50 s, 45 °C for 50 s, and 72 °C for 60 s; 30 cycles at 94 °C for 50 s, 50 °C for 50 s, and 72 °C for 60 s; and a final elongation at 72 °C for 5 min [14]. The PCR products were purified using a QIAquick PCR Purification Kit (Qiagen, Germany) and sequenced in both directions using an ABI 3730 automated sequencer (Applied Biosystems, Bedford, MA, USA). The new sequences were aligned using MEGA 6.06 with built-in ClustalW [18]. Phylogenetic analysis and pairwise distance measurements were performed using the maximum likelihood method based on the Kimura 2-parameter model [19], with 1000 bootstrap replications in MEGA 6.06. Six sequences were deposited in GenBank under accession numbers LC653016-653021 for the new genus (Table 1) [14,20,21,22,23].
2.3. Morphological Investigation
Taxonomic observations and measurements were performed on live and preserved specimens (Figure 2). Measurements were made using ImageJ [24] to the nearest 0.1 mm. For nematocyst identification in the medusae, squash prepared from fresh tissues was examined under a compound microscope (ECLIPSE Ci, Nikon, Tokyo, Japan). Nematocysts were identified according to a previously described method [1,25]. To determine the abundance of nematocyst types in medusae, approximately 100 nematocysts were identified, measured, and counted in unregistered specimens. Measurements were performed using ImageJ [24] to the nearest 0.1 µm.
3. Results
3.1. Molecular Phylogenetic Analysis
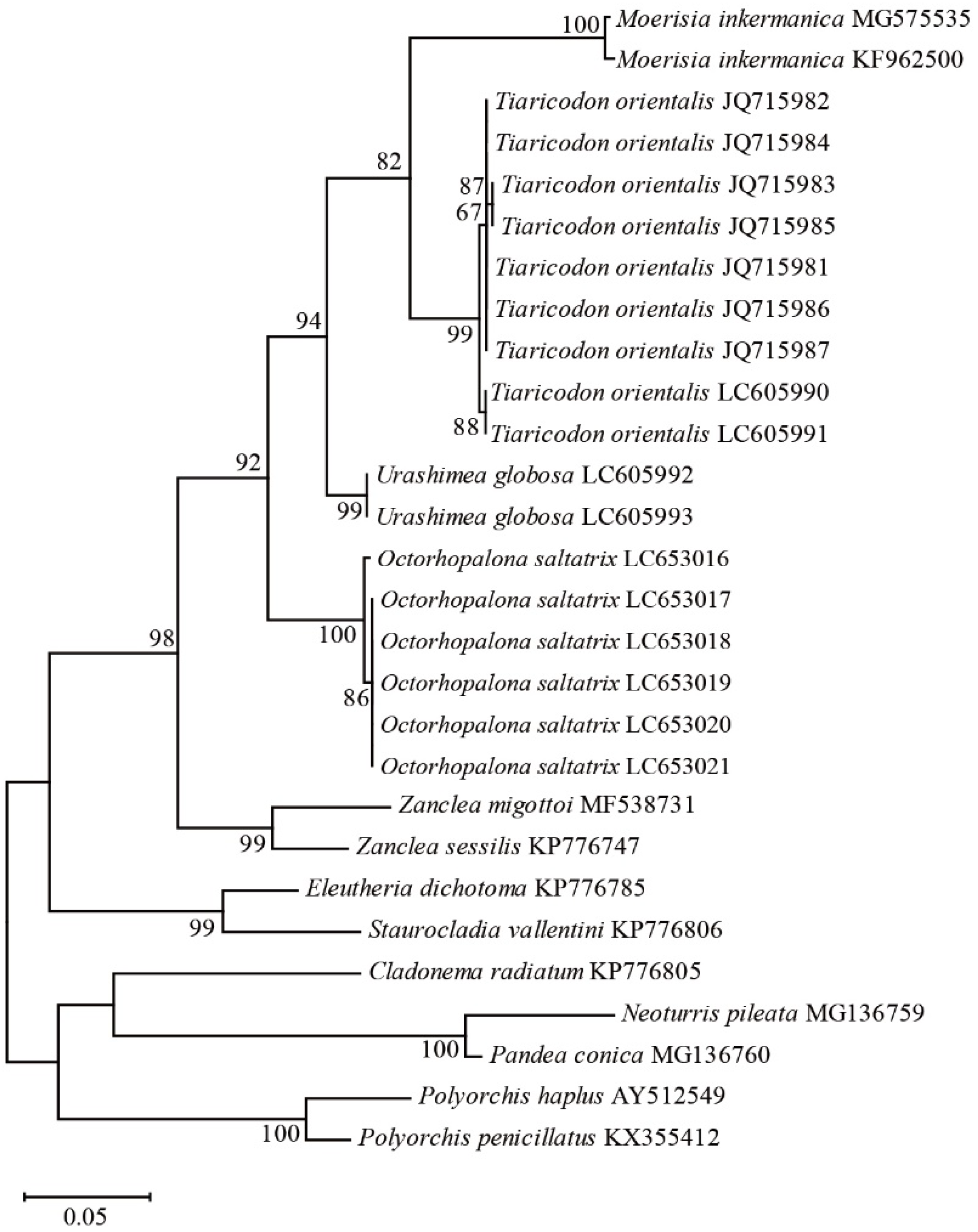
We sequenced six individuals of Octorhopalona saltatrix for the 16S rDNA fragments, in addition to 12 Anthoathecata taxa, for statistical analyses. The maximum likelihood tree constructed for the family Halimedusidae based on the 16S rDNA sequences (Figure 3) comprised four major clades formed in the suborder Capitata: (1) Moerisia inkermanica, (2) Tiaricodon orientalis, (3) Urashimea globosa, and (4) Octorhopalona. The family paraphyly of Halimedusidae, including clades 1–3 and 4, was evident in the 16S rDNA phylogenetic tree with high bootstrap values (92%), which supports the validity of the genus. Moreover, Halimedusidae includes M. inkermanica, which is presently classified in the family Moerisiidae.
The Kimura two-parameter distance was 0.094–0.099 between Octorhopalona saltatrix n. sp. and Tiaricodon, 0.066 between Octorhopalona and Urashimea, and 0.064–0.067 between Tiaricodon and Urashimea (Table 2). The distance was greater (0.093–0.189) between different Octorhopalona and other Anthoathecata families, Cladonematidae, Corynidae, Moerisiidae, Pandeidae, Polyorchidae, and Zancleidae.
3.2. Morphological Investigation
3.2.1. Systematics
Phylum Cnidaria Hatschek, 1888.
Subphylum Medusozoa Petersen, 1979.
Class Hydrozoa Owen, 1843.
Subclass Hydroidolina Collins, 2000.
Order Anthoathecata Cornelius, 1992.
Suborder Capitata Kühn, 1913.
Family Halimedusidae Arai and Brinckmann-Voss, 1980.
Genus Octorhopalona gen. nov.
New Japanese name: Otohime-kurage-zoku.
3.2.2. Genus Diagnosis
Halimedusidae, with eight nematocyst tracks in the exumbrella; eight radial canals; eight characteristic perradial and interradial blead-shaped ‘smooth peaks’ in the mesoglea between the radial canals; eight tentacles, with numerous stalked nematocyst knobs over their entire length; and tentacular bulbs swollen with an abaxial ocellus.
Type species. Octorhopalona saltatrix sp. nov. is designated here.
The genus name ‘Octorhopalona’ is taken from the Greek words ‘octo’ and ‘rhopalon’, meaning ‘eight’ and ‘club’, respectively; the gender is feminine. The name indicates that the medusa bears eight tentacles that look like eight clubs.
3.2.3. Species Description
Octorhopalona saltatrix sp. nov.
New Japanese name. Otohime-kurage.
Materials examined. Holotype: INM-1-96244; Oarai Fishing Port, Higashi-Ibaraki, Ibaraki Prefecture, eastern Japan; 36°18′39.1″ N, 140°34′39.5″ E; 15 December 2008; collector: Shinsuke Saito. Paratypes: KBF-M30; Enoshima, Fujisawa, Kanagawa Prefecture, eastern Japan; 35°17′52.4″ N, 139°28′32.2″ E; 7 May 2021; collector: Gaku Yamamoto; KBF-M31, same locality as KBF-M30; 24 September 2018; collector: Gaku Yamamoto: KBF-M32; Shimonokae Fishing Port, Tosashimizu, Kochi, Japan; 32°51′42.96″ N, 132°57′34.80″ E; 30 November 2020; collector: Sho Toshino; NSMT-Co1802. Katase Fishing Port, Fujisawa, Kanagawa Prefecture, eastern Japan; 35°18′23.6″ N 139°28′51.6″ E; 14 July 2019; collector: Haruka Onishi; NSMT-Co1803; Enoshima, Fujisawa, Kanagawa Prefecture, eastern Japan; 35°17′52.4″ N, 139°28′32.2″ E; 9 November 2018; collector: Haruka Onishi; NSMT-Co1804, same locality as NSMT-Co1803; 22 November 2018; collector: Gaku Yamamoto; and NSMT-Co1805, same locality as NSMT-Co1803; 14 July 2019; collector: Gaku Yamamoto (Table 3).
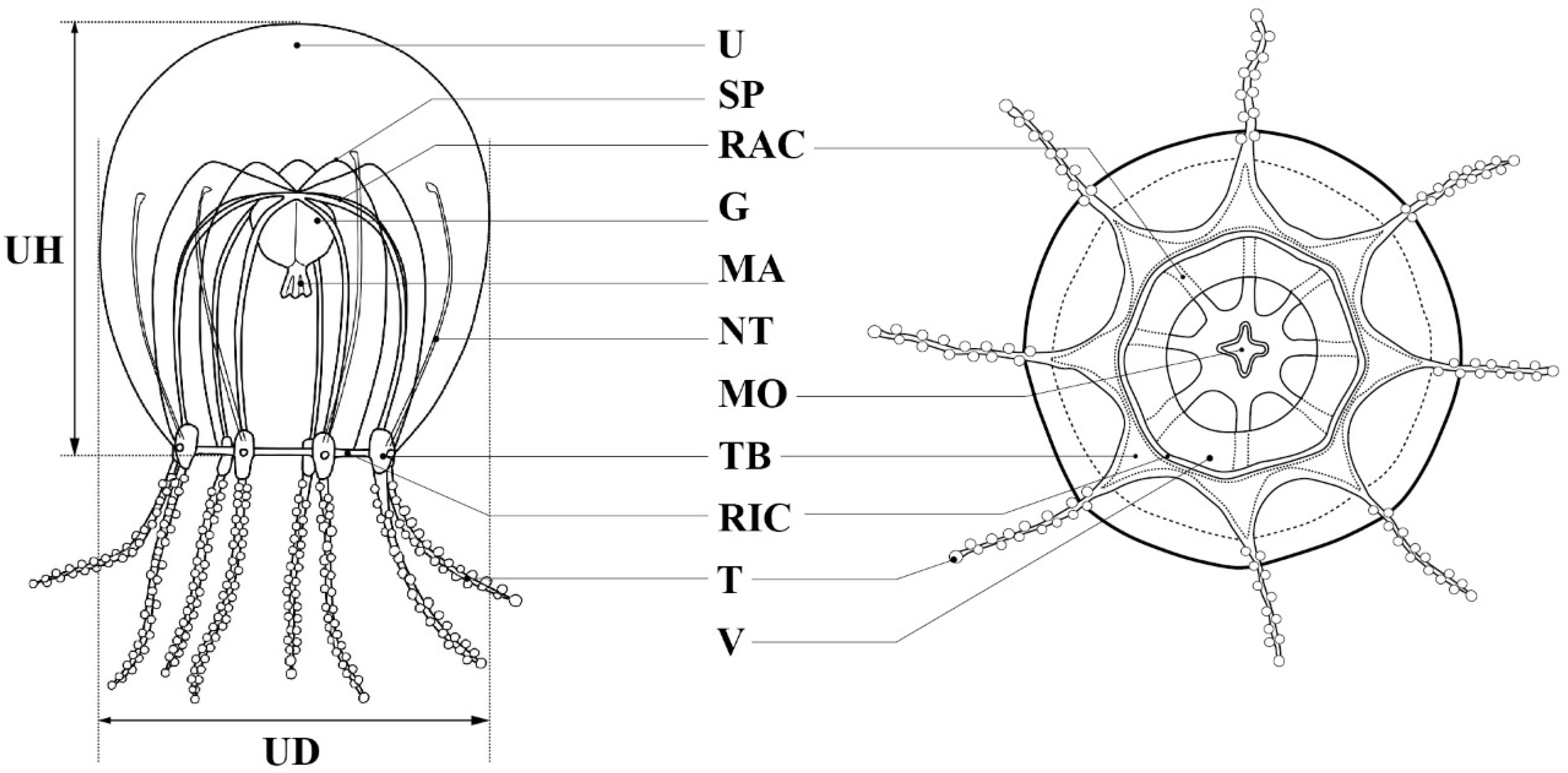
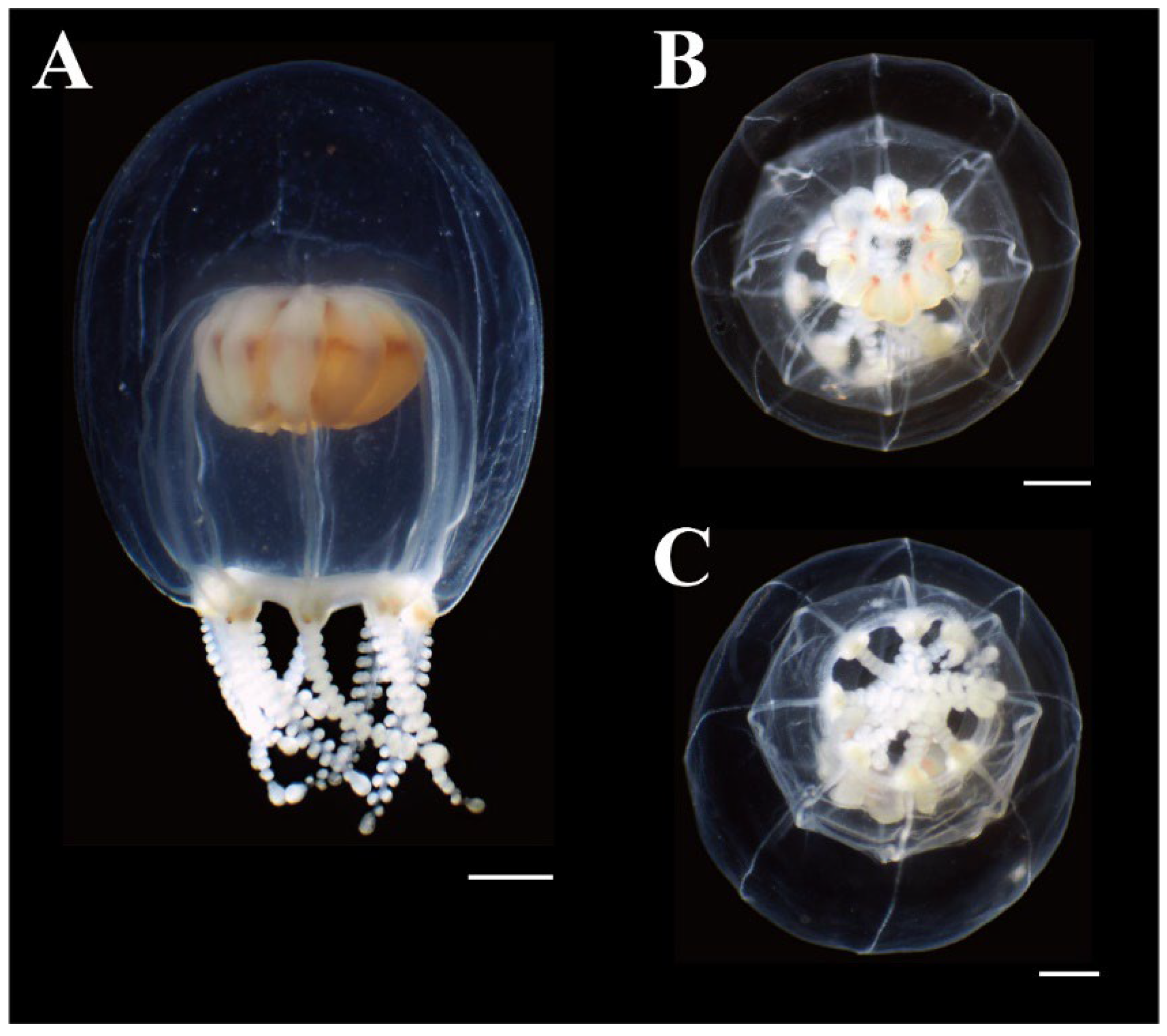
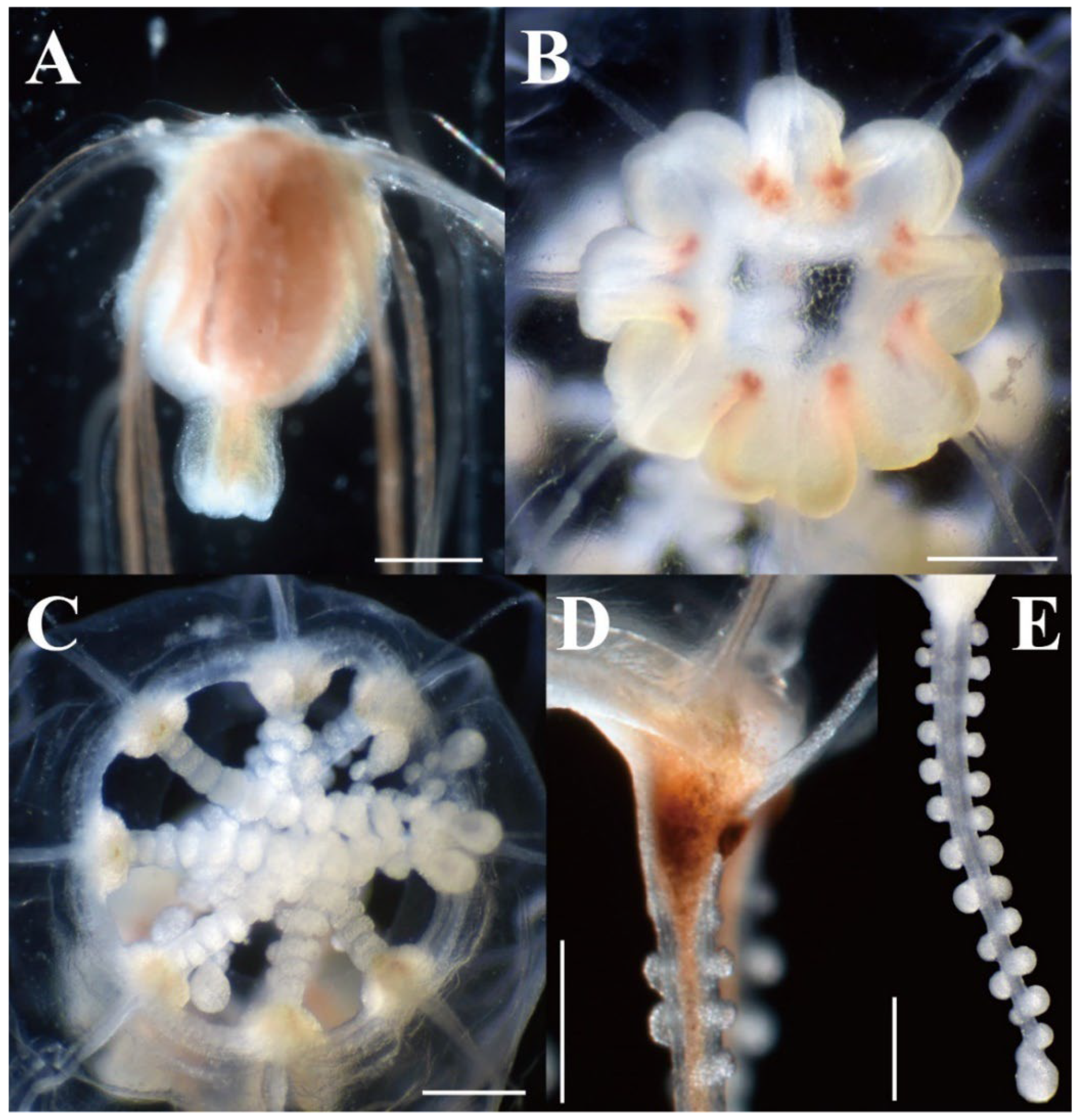
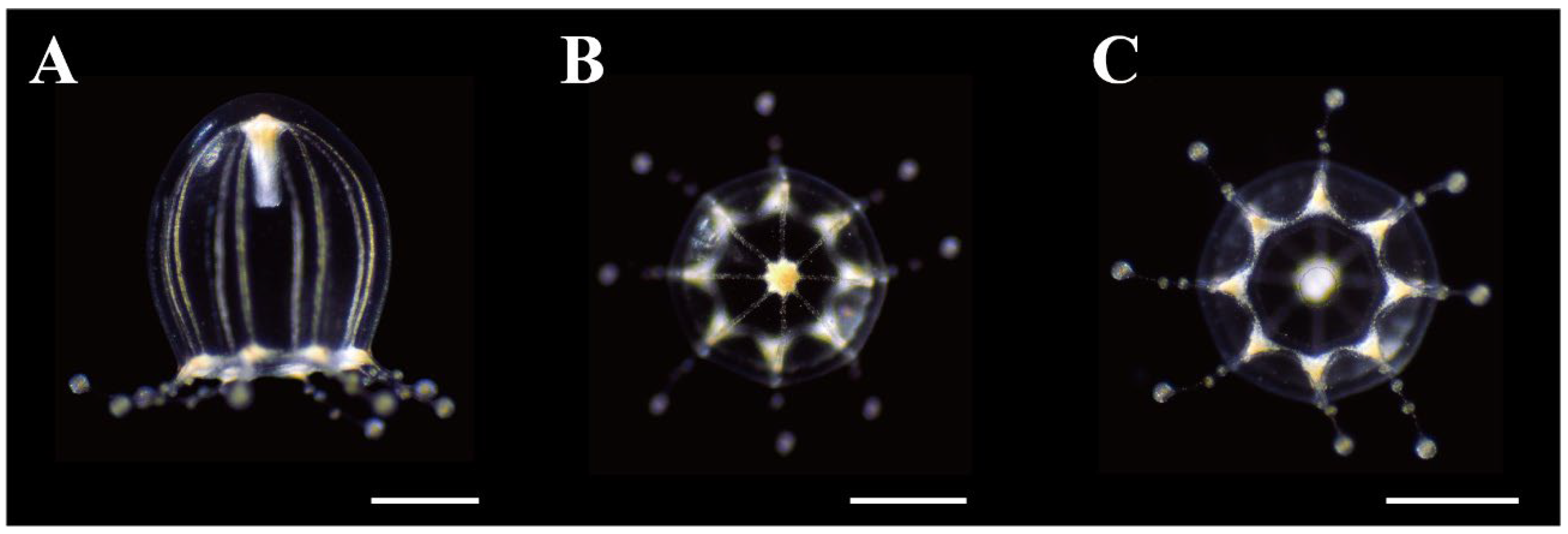
Description. Mature medusae have bell-shaped umbrellas (Figure 4 and Figure 5A–C), 9 mm in height and 9 mm in diameter (Table 3). The umbrella apex is rounded and the mesoglea is thickened (Figure 4 and Figure 5A). Exumbrella are smooth and nematocysts are sparsely scattered. Nematocysts track eight, perradial and interradial, on the exumbrella about two-thirds of the umbrella height (UH). Manubrium hang in the umbrella cavity, quadrilateral bottom, and are light brown or translucent (Figure 6A). The extended manubrium length was approximately 2 mm and did not extend beyond the umbrella margin. The gonads cover the entire surface of the manubrium, except for the mouth lip. The mouth cruciform has four lips. The stomach has short, sac-like perradial and interradial lobes (Figure 6B). There are eight characteristic interradial blead-shaped ‘smooth peaks’ in the mesoglea between the radial canals, rising above the level of the radial canals (Figure 6A). There are eight radial canals and one circular canal (Figure 5B,C and Figure 6B,C). The velum is narrow, with a velarial width of 10% of the umbrella diameter (UD) (Figure 6C). The tentacular bulbs were swollen, each with a dark brown abaxial ocellus (Figure 6D). There were eight tentacles, with numerous stalked nematocyst knobs in their entire length, approximately the same length as live umbrella height (UH) (Figure 6E).
The smallest young medusa had a UH of 1.3 mm and a UD of 0.9 mm. The mesoglea at the apex of the exumbrella was thinner than that of adults (Figure 7A). Nematocyst tracks on the exumbrella were approximately two-thirds of the UH. The manubrium was short, thin, translucent to whitish, and approximately one-third the length of the UH. Its mouth was simple and circular. It possessed eight radial canals and a single circular canal. Eight interradial peaks were absent in the mesoglea between the radial canals (Figure 7B). The velum was wide, and the velarial width was 20% of the UD (Figure 7C). The tentacular bulbs were swollen, each with a red-brown abaxial ocellus. There were eight tentacles, pearl-string-like, and 1–3 white or white-brown nematocyst batteries aligned, one-half of the UH.
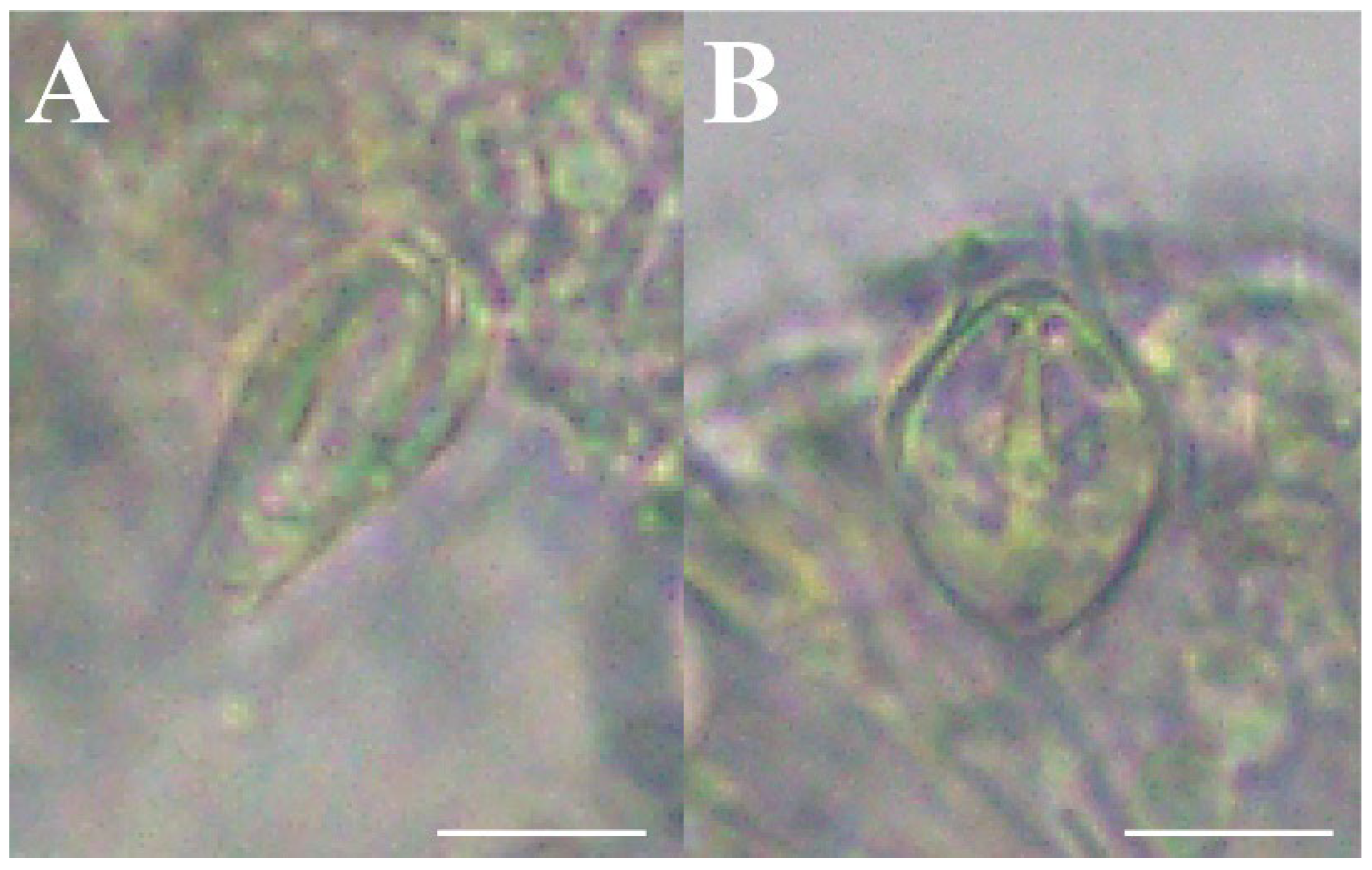
Cnidome. Two nematocyst types were identified and measured (Table 4, Figure 8). Exumbrella: stenoteles. Manubrium: desmonemes and stenoteles. Tentacle: desmonemes and stenoteles. Tentacle bulb: desmonemes and stenoteles.
Habitat and ecology. Medusae of Octorhopalona saltatrix appeared on the surface of shallow waters (5–10 m in depth) off Oarai (eastern Japan) during November and December; in Sagami Bay (western Japan) from May to November; and in Tosa Bay (Western Japan) during November. Polyps of the species have never been found in the wild; however, medusa budding seems to occur between spring and fall in Japanese waters. In some specimens, the mesoglea at the apical part was infested with flukes.
Etymology. The specific name ‘saltatrix’ is taken from the Latin word ‘saltatrix’, meaning ‘female dancer’. The gender is feminine. The name reflects the medusa swimming like a female dancer using its umbrella and eight tentacles.
4. Discussion
4.1. Molecular Phylogenetic Analysis
The paraphyly of Halimedusidae, including Octorhopalona and three species (U. globosa, T. coeruleus, and M. intermanica), was evident in the 16S rDNA phylogenetic tree with high bootstrap values (92%), which supports the validity of the new species. The maximum-likelihood tree includes Moerisia inkermanica (Moerisiidae), because Tiaricodon was placed in the family Moerisiidae [26]. The tree suggests that M. inkermanica may be derived from within Halimedusidae.
The Kimura two-parameter distance was 0.094–0.099 between Octorhopalona saltatrix n. sp. and Tiaricodon, 0.066 between Octorhopalona and Urashimea, and 0.064–0.067 between Tiaricodon and Urashimea. The distance was greater (0.093–0.189) between different Octorhopalona and other Anthoathecata families, Cladonematidae, Corynidae, Moerisiidae, Pandeidae, Polyorchidae, and Zancleidae. In the class Hydrozoa, the intergeneric variability was 0.06–0.24 [22]. Given the results of the K2P distances in this study, Octorhopalona was considered a genetically independent genus in the family Halimedusidae.
4.2. Morphological Investigation
A comparison of the key features of the Halimedusidae genera is presented in Table 5. Octorhopalona saltatrix can be distinguished from all other Halimedusidae species by the number of tentacles and radial canals, as well as the shape of the tentacles. All the species in the family Halimedusidae have a bell-shaped umbrella, interradial peaks in the subumbrella, a red band on the manubrium, and abaxial ocelli on the basal tentacle bulbs. There are four tentacles (Tiaricodon and Urashimea) [3,4,5,6,26,27,28,29,30,31], four of which are perradial, and ten to eleven are interradial (Halimedusa) [1,9]. The tentacles are moniliform (Halimedusa and Tiaricodon) and numerous stalked nematocyst knobs cover their entire length (Octorhopalona and Urashimea). All Halimedusidae species have four radial canals and four interradial peaks in the subumbrella; however, only the Octorhopalona has eight radial canals and eight interradial peaks in the subumbrella. Therefore, we propose to amend the family Halimedusidae as follows:
- Family Halimedusidae Arai and Brinckmann-Voss, 1980, sens. emend.
Diagnosis. Anthoathecata, with four or eight radial canals, a low peduncle, and distinct interradial or interradial and perradial peaks in the jelly above the base of the manubrium; red band on the manubrium, gonads extending out from the manubrium as lobes below the upper portions of the radial canals, but without mesenteries; mouth with lips lined by a row of sessile nematocyst clusters; four perradial hollow tentacles, or four perradial tentacles and four interradial single tentacles, or groups of hollow tentacles; cylindrical marginal bulbs, each with an abaxial ocellus.
Cnidomes are important for identification purposes. Two types of nematocysts, stenoteles and desmonemes, have been examined in Octorhopalona, Halimedusa, and Tiaricodon [1]. Halimedusa bears microbasic euryteles (medusae) and isorhizas or anisorhizas (polyps) [1]. Tiaricodon sp. from New Zealand and Brazil bear heteronemes and microbasic euryteles, respectively [5,28]. Cnidomes of Urashimea have not yet been examined.
5. Conclusions
Our morphological and molecular phylogenetic analyses suggest that these specimens collected off Oarai, Sagami Bay, and Tosa Bay, Japan are a new genera and new species belonging to the family Halimedusidae. This species appears in shallow waters in Japan during May and December. Polyps of this species have never been found in the wild. Additional sampling is required to understand their ecology and development.
Three genera, Halimedusa, Tiaricodon, and Urashimea are currently classified in the family Halimedusidae, defined by [1] as: Anthomedusae with four radial canals, with low peduncle and distinct interradial peaks in jelly above base of the manubrium; gonads extending out from the manubrium as lobes below the upper portions of the four radial canals, but without mesenteries; quadratic mouth with lips lined by a row of sessile nematocyst clusters, with four perradial hollow tentacles or with four perradial tentacles and four interradial groups of hollow tentacles; and possessing cylindrical marginal bulbs each with an abaxial ocellus. However, Octorhopalona saltatrix has eight tentacles, eight radial canals, and four interradial and four perradial peaks. Therefore, we propose to amend the family Halimedusidae as discussed.
Author Contributions
Conceptualization, S.T. and G.Y.; methodology, S.T., G.Y. and S.S.; validation, S.T. and G.Y.; formal analysis, S.T. and G.Y.; investigation, S.T., G.Y. and S.S.; resources, S.T., G.Y. and S.S.; data curation, S.T. and G.Y.; writing—original draft preparation, S.T. and G.Y.; writing—review and editing, S.T., G.Y. and S.S.; supervision, S.T.; funding acquisition, S.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Japan Fund for Global Environment and JSPS KAKENHI (grant numbers JP18K14791 and 21K15158, awarded to S. Toshino).
Data Availability Statement
All datasets collected and analyzed during the current study are available from the corresponding author upon request.
Acknowledgments
We would like to express our sincere thanks to Junko Fukada, Takuma Mezaki, Yuji Ise, Tatsuki Koido, Kenjiro Hinode, Takaya Kitamura, Reina Tsujimoto, Takeru Yoshioka, Chika Nagaoka (Kuroshio Biological Research Foundation), Yojiro Shiba (Ibaraki Prefectural Oarai Aquarium), Hiromi Ikezawa (Ibaraki Nature Museum), Kazuhisa Hori, Takuji Oyama, Tadao Sakiyama, Mitsugu Kitada, Aya Adachi (Enoshima Aquarium), and Haruka Onishi (Anicom Insurance, Inc.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mills, C.E. The life cycle of Halimedusa typus, with discussion of other species closely related to the family Halimedusidae (Hydrozoa, Capitata, Anthomedusae). Sci. Mar. 2000, 64 (Suppl. 1), 97–106. [Google Scholar]
- Bouillon, J.; Gravili, C.; Gili, J.M.; Boero, F. An Introduction to Hydrozoa; Mémoires du Muséum National d’Histoire Naturelle: Paris, France, 2006; Volume 194, pp. 1–591. [Google Scholar]
- Yamamoto, G.; Toshino, S. Tiaricodon orientalis sp. nov., a new species (Hydrozoa, Anthoathecata, Halimedusidae) from Sagami Bay, eastern Japan. Plankton Benthos Res. 2021, 16, 129–138. [Google Scholar] [CrossRef]
- Browne, E.T. A preliminary report on Hydromedusae from the Falkland Islands. Ann. Mag. Nat. Hist. 1902, 9, 272–284. [Google Scholar] [CrossRef] [Green Version]
- Schuchert, P. The marine fauna of New Zealand: Athecate hydroids and their medusae (Cnidaria: Hydrozoa). N. Z. Oceanogr. Mem. 1996, 106, 1–159. [Google Scholar]
- Xu, Z.; Chen, Y. Life cycle of Tiaricodon coerules from Xiamen Harbour. J. Oceanogr. Taiwan Strait 1998, 17, 129–133, (In Chinese with English abstract). [Google Scholar]
- Uchida, T.; Nagao, Z. On the systematic position of the hydroid genus Urashimea. Annot. Zool. Jpn. 1961, 34, 200–204. [Google Scholar]
- Kishinouye, K. Some medusae of Japanese waters. J. College Sci. Imp. Univ. Tokyo Japan 1910, 27, 23–35. [Google Scholar]
- Bigelow, H.B. Halimedusa, a new genus of Anthomedusae. Trans. R. Soc. Can. Sect. 1916, 4, 91–95. [Google Scholar]
- Bouillon, J. Hydromedusae of the New Zealand Oceanographic Institute (Hydrozoa, Cnidaria). N. Z. J. Zool. 1995, 22, 223–238. [Google Scholar] [CrossRef] [Green Version]
- Arai, M.N.; Brinckmann-Voss, A. Hydromedusae of British Columbia and Puget Sound. Can. Bull. Fish Aquat. Sci. 1980, 204, 1–192. [Google Scholar]
- Bouillon, J.; Boero, F. Synopsis of the families and genera of the Hydromedusae of the world, with a list of the worldwide species. Thalassia Salentina 2000, 24, 47–296. [Google Scholar]
- Kubota, S.; Gravili, C. A list of hydromedusae (excluding Siphonophora, Milleporidae and Actinulidae) in Japan. Nankiseibutsu 2007, 49, 189–204. [Google Scholar]
- Collins, A.G.; Winkelmann, S.; Hadrys, H.; Schierwater, B. Phylogeny of Capitata and Corynidae (Cnidaria, Hydrozoa) in light of mitochondrial 16S rDNA data. Zool. Scr. 2008, 34, 91–99. [Google Scholar] [CrossRef]
- Schuchert, P. DNA barcoding of some Pandeidae species (Cnidaria, Hydrozoa, Anthoathecata). Rev. Suisse Zool. 2018, 125, 101–127. [Google Scholar]
- Collins, A.G.; Bentlage, B.; Lindner, A.; Lindsay, D.; Haddock, S.H.D.; Jarms, G.; Norenburg, J.L.; Jankowski, T.; Cartwright, P. Phylogenetics of Trachylina (Cnidaria: Hydrozoa) with new insights on the evolution of some problematical taxa. J. Mar. Biol. Assoc. UK 2008, 88, 1673–1685. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, C.W.; Buss, L.W. Molecular evidence for multiple episodes of pedomorphosis in the family Hydractiniidae. Biochem. Syst. Ecol. 1993, 21, 57–69. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Restaino, D.J.; Bologna, P.A.; Gaynor, J.J.; Buchanan, G.A.; Bilinski, J.J. Who’s lurking in your lagoon? First occurrence of the invasive hydrozoan Moerisia sp. (Cnidaria: Hydrozoa) in New Jersey, USA. BioInvasions Rec. 2018, 7, 223–228. [Google Scholar] [CrossRef]
- Nawrocki, A.M.; Schuchert, P.; Cartwright, P. Phylogenetics and evolution of Capitata (Cnidaria: Hydrozoa), and the systematics of Corynidae. Zool. Scr. 2010, 39, 290–304. [Google Scholar] [CrossRef]
- Zheng, L.; He, J.; Lin, Y.; Cao, W.; Zhang, W. 16S rRNA is a better choice than COI for DNA barcoding hydrozoans in the coastal waters of China. Acta Oceanol. Sin. 2014, 33, 55–76. [Google Scholar] [CrossRef]
- Mendoza-Becerril, M.A.; Simões, N.; Genzano, G. Benthic hydroids (Cnidaria, Hydrozoa) from Alacranes Reef, Gulf of Mexico, Mexico. Bull. Mar. Sci. 2018, 94, 125–142. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Östman, C. A guideline to nematocyst nomenclature and classification, and some notes on the systematic value of nematocysts. Sci. Mar. 2000, 64 (Suppl. 1), 31–46. [Google Scholar] [CrossRef]
- Browne, E.T.; Kramp, P.L. Hydromedusae from the Falkland Islands. Discov. Rep. 1939, 18, 265–322. [Google Scholar]
- Xu, Z.; Huang, J.Q.; Lin, M.; Guo, D.H.; Wang, C.G. The Superclass Hydrozoa of the Phylum Cnidaria in China; China Ocean Press: Beijing, China, 2014; pp. 1–945, (In Chinese with English Summary). [Google Scholar]
- Nogueira, M.J.; da Costa, B.S.P.; Martinez, T.A.; Brandini, F.P.; Miyashita, L.K. Diversity of gelatinous zooplankton (Cnidaria, Ctenophora, Chaetognatha and Tunicata) from a subtropical estuarine system, southeast Brazil. Mar. Biodivers. 2019, 49, 1283–1298. [Google Scholar] [CrossRef]
- Kramp, P.L. Synopsis of the medusae of the world. J. Mar. Biol. Assoc. UK 1961, 40, 1–469. [Google Scholar] [CrossRef]
- Buecher, E.; Gibbons, M.J. Interannual variation in the composition of the assemblages of medusae and ctenophores in St Helena Bay, Southern Benguela ecosystem. Sci. Mar. 2000, 64 (Suppl. 1), 123–134. [Google Scholar] [CrossRef] [Green Version]
- Minemizu, R.; Kubota, S.; Hirano, Y.; Lindsay, D.J. A Photographic Guide to the Jellyfishes of Japan; Heibonsha: Tokyo, Japan, 2015; pp. 1–360. [Google Scholar]
Figure 1.
Map of the sampling sites. (A) Oarai Fishing Port, Higashi-Ibaraki, Ibaraki Prefecture; (B) Enoshima, Fujisawa, Kanagawa Prefecture: (1) Katase Fishing Port, (2) Enoshima; (C) Shimonokae Fishing Port, Tosashimizu, Kochi Prefecture. Black closed circles are sampling sites.
Figure 1.
Map of the sampling sites. (A) Oarai Fishing Port, Higashi-Ibaraki, Ibaraki Prefecture; (B) Enoshima, Fujisawa, Kanagawa Prefecture: (1) Katase Fishing Port, (2) Enoshima; (C) Shimonokae Fishing Port, Tosashimizu, Kochi Prefecture. Black closed circles are sampling sites.

Figure 2.
Key characters for identification and measurement of parts of the Halimedusidae: G = gonad; MA = manubrium; MO = mouth; NT = nematocyst track; RAC = radial canal; RIC = ring canal; SP = subumbrellar peak; T = tentacle; TB = tentacle bulb; U = umbrella; UD = umbrella diameter; UH = umbrella height; V = velum.
Figure 2.
Key characters for identification and measurement of parts of the Halimedusidae: G = gonad; MA = manubrium; MO = mouth; NT = nematocyst track; RAC = radial canal; RIC = ring canal; SP = subumbrellar peak; T = tentacle; TB = tentacle bulb; U = umbrella; UD = umbrella diameter; UH = umbrella height; V = velum.

Figure 3.
Nuclear 16S rDNA maximum-likelihood tree for 13 anthoathcata taxa based on the General Time Reversible model: Scale bar indicates branch length in substitutions per site. Nodal support values are presented as the ML bootstrap value; only values >50% are shown.
Figure 3.
Nuclear 16S rDNA maximum-likelihood tree for 13 anthoathcata taxa based on the General Time Reversible model: Scale bar indicates branch length in substitutions per site. Nodal support values are presented as the ML bootstrap value; only values >50% are shown.

Figure 4.
Live mature medusae of Octorhopalona saltatrix sp. nov. (unregisted specimen). Scale bar: 1 mm.
Figure 4.
Live mature medusae of Octorhopalona saltatrix sp. nov. (unregisted specimen). Scale bar: 1 mm.

Figure 5.
Preserved mature medusae of Octorhopalona saltatrix sp. nov. (NSMT-Co1802): (A) lateral, (B) apical, and (C) oral views. Scale bars: 1 mm.
Figure 5.
Preserved mature medusae of Octorhopalona saltatrix sp. nov. (NSMT-Co1802): (A) lateral, (B) apical, and (C) oral views. Scale bars: 1 mm.

Figure 6.
Live and preserved mature medusae of Octorhopalona saltatrix sp. nov. (unregisted specimen and NSMT-Co1802): (A) upper part of umbrella, (B) umbrella, apical view, (C) velum, (D) tentacular bulb, (E) tentacle. Scale bars: 0.5 mm.
Figure 6.
Live and preserved mature medusae of Octorhopalona saltatrix sp. nov. (unregisted specimen and NSMT-Co1802): (A) upper part of umbrella, (B) umbrella, apical view, (C) velum, (D) tentacular bulb, (E) tentacle. Scale bars: 0.5 mm.

Figure 7.
Live young medusae of Octorhopalona saltatrix sp. nov. (KBF-M32). (A) lateral, (B) apical and, (C) oral views. Scale bars: 0.5 mm.
Figure 7.
Live young medusae of Octorhopalona saltatrix sp. nov. (KBF-M32). (A) lateral, (B) apical and, (C) oral views. Scale bars: 0.5 mm.

Figure 8.
Nematocysts of Octorhopalona saltatrix sp. nov. (unregistered specimen): (A) desmoneme, (B) stenotele. Scale bars: 10 μm.
Figure 8.
Nematocysts of Octorhopalona saltatrix sp. nov. (unregistered specimen): (A) desmoneme, (B) stenotele. Scale bars: 10 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Taxa included in the phylogenetic analyses and GenBank accession numbers for the 16S rDNA sequences. Sequences obtained in this study are in bold.
Table 1.
Taxa included in the phylogenetic analyses and GenBank accession numbers for the 16S rDNA sequences. Sequences obtained in this study are in bold.
| Species | Accession No. | Locality (Origin) | Reference |
|---|---|---|---|
| Cladonema radiatum | KP776805 | France: Roscoff, vivier | Unpublished |
| Eleutheria dichotoma | KP776785 | Norway: Raunefjord | Unpublished |
| Moerisia inkermanica | KF962500 | China | Unpublished |
| Moerisia inkermanica | MG575535 | USA: Bayville, NJ | [20] |
| Neoturris pileata | MG136759 | France: Bay of Villefranche-sur-Mer | [21] |
| Octorhopalona saltatrix | LC653016 | Japan: Enoshima, Fujisawa, Kanagawa | This study |
| Octorhopalona saltatrix | LC653017 | Japan: Enoshima, Fujisawa, Kanagawa | This study |
| Octorhopalona saltatrix | LC653018 | Japan: Enoshima, Fujisawa, Kanagawa | This study |
| Octorhopalona saltatrix | LC653019 | Japan: Enoshima, Fujisawa, Kanagawa | This study |
| Octorhopalona saltatrix | LC653020 | Japan: Enoshima, Fujisawa, Kanagawa | This study |
| Octorhopalona saltatrix | LC653021 | Japan: Enoshima, Fujisawa, Kanagawa | This study |
| Pandea conica | MG136760 | France: Bay of Villefranche-sur-Mer | [21] |
| Polyorchis haplus | AY512549 | USA: San Francisco Bay | [21] |
| Polyorchis penicillatus | KX355412 | USA: Friday Harbour | [14] |
| Staurocladia vallentini | KP776806 | Chile: Region de los Rios, Chaihuin | Unpublished |
| Tiaricodon orientalis | JQ715981 | China | Unpublished |
| Tiaricodon orientalis | JQ715982 | China | Unpublished |
| Tiaricodon orientalis | JQ715983 | China | Unpublished |
| Tiaricodon orientalis | JQ715984 | China | Unpublished |
| Tiaricodon orientalis | JQ715985 | China | Unpublished |
| Tiaricodon orientalis | JQ715986 | China | Unpublished |
| Tiaricodon orientalis | JQ715987 | China | Unpublished |
| Tiaricodon orientalis | LC605990 | Japan: Enoshima, Fujisawa, Kanagawa | [22] |
| Tiaricodon orientalis | LC605991 | Japan: Enoshima, Fujisawa, Kanagawa | [22] |
| Urashimea globosa | LC605992 | Japan: Tokoro Fishing Port, Kitami, Hokkaido | [22] |
| Urashimea globosa | LC605993 | Japan: Tokoro Fishing Port, Kitami, Hokkaido | [22] |
| Zanclea migottoi | MF538731 | Mexico: Gulf of Mexico, Alacranes Reef | [23] |
| Zanclea sessilis | KP776747 | Spain: Mallorca, Cala Murada | Unpublished |
Table 2.
Pairwise genetic distances (K2P) based on 440 positions of 16S sequences among Anthomedusae. The analysis involved 28 sequences.
Table 2.
Pairwise genetic distances (K2P) based on 440 positions of 16S sequences among Anthomedusae. The analysis involved 28 sequences.
| No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Tiaricodon orientalis JQ715981 | |||||||||||||||||||||||||||
| 2 | Tiaricodon orientalis JQ715982 | 0.000 | ||||||||||||||||||||||||||
| 3 | Tiaricodon orientalis JQ715983 | 0.002 | 0.002 | |||||||||||||||||||||||||
| 4 | Tiaricodon orientalis JQ715984 | 0.000 | 0.000 | 0.002 | ||||||||||||||||||||||||
| 5 | Tiaricodon orientalis JQ715985 | 0.002 | 0.002 | 0.000 | 0.002 | |||||||||||||||||||||||
| 6 | Tiaricodon orientalis JQ715986 | 0.000 | 0.000 | 0.002 | 0.000 | 0.002 | ||||||||||||||||||||||
| 7 | Tiaricodon orientalis JQ715987 | 0.000 | 0.000 | 0.002 | 0.000 | 0.002 | 0.000 | |||||||||||||||||||||
| 8 | Tiaricodon orientalis LC605990 | 0.005 | 0.005 | 0.007 | 0.005 | 0.007 | 0.005 | 0.005 | ||||||||||||||||||||
| 9 | Tiaricodon orientalis LC605991 | 0.005 | 0.005 | 0.007 | 0.005 | 0.007 | 0.005 | 0.005 | 0.000 | |||||||||||||||||||
| 10 | Urashimea globosa LC605992 | 0.064 | 0.064 | 0.067 | 0.064 | 0.067 | 0.064 | 0.064 | 0.064 | 0.064 | ||||||||||||||||||
| 11 | Urashimea globosa LC605993 | 0.064 | 0.064 | 0.067 | 0.064 | 0.067 | 0.064 | 0.064 | 0.064 | 0.064 | 0.000 | |||||||||||||||||
| 12 | Octorhopalona saltatrix LC653016 | 0.094 | 0.094 | 0.096 | 0.094 | 0.096 | 0.094 | 0.094 | 0.094 | 0.094 | 0.066 | 0.066 | ||||||||||||||||
| 13 | Octorhopalona saltatrix LC653017 | 0.096 | 0.096 | 0.099 | 0.096 | 0.099 | 0.096 | 0.096 | 0.096 | 0.096 | 0.066 | 0.066 | 0.005 | |||||||||||||||
| 14 | Octorhopalona saltatrix LC653018 | 0.096 | 0.096 | 0.099 | 0.096 | 0.099 | 0.096 | 0.096 | 0.096 | 0.096 | 0.066 | 0.066 | 0.005 | 0.000 | ||||||||||||||
| 15 | Octorhopalona saltatrix LC653019 | 0.096 | 0.096 | 0.099 | 0.096 | 0.099 | 0.096 | 0.096 | 0.096 | 0.096 | 0.066 | 0.066 | 0.005 | 0.000 | 0.000 | |||||||||||||
| 16 | Octorhopalona saltatrix LC653020 | 0.096 | 0.096 | 0.099 | 0.096 | 0.099 | 0.096 | 0.096 | 0.096 | 0.096 | 0.066 | 0.066 | 0.005 | 0.000 | 0.000 | 0.000 | ||||||||||||
| 17 | Octorhopalona saltatrix LC653021 | 0.096 | 0.096 | 0.099 | 0.096 | 0.099 | 0.096 | 0.096 | 0.096 | 0.096 | 0.066 | 0.066 | 0.005 | 0.000 | 0.000 | 0.000 | 0.000 | |||||||||||
| 18 | Moerisia inkermanica KF962500 | 0.080 | 0.080 | 0.083 | 0.080 | 0.083 | 0.080 | 0.080 | 0.086 | 0.086 | 0.091 | 0.091 | 0.111 | 0.116 | 0.116 | 0.116 | 0.116 | 0.116 | ||||||||||
| 19 | Moerisia inkermanica MG575535 | 0.086 | 0.086 | 0.089 | 0.086 | 0.089 | 0.086 | 0.086 | 0.086 | 0.086 | 0.091 | 0.091 | 0.111 | 0.116 | 0.116 | 0.116 | 0.116 | 0.116 | 0.005 | |||||||||
| 20 | Zanclea migottoi MF538731 | 0.134 | 0.134 | 0.137 | 0.134 | 0.137 | 0.134 | 0.134 | 0.134 | 0.134 | 0.111 | 0.111 | 0.111 | 0.116 | 0.116 | 0.116 | 0.116 | 0.116 | 0.149 | 0.149 | ||||||||
| 21 | Zanclea sessilis KP776747 | 0.136 | 0.136 | 0.139 | 0.136 | 0.139 | 0.136 | 0.136 | 0.136 | 0.136 | 0.099 | 0.099 | 0.096 | 0.093 | 0.093 | 0.093 | 0.093 | 0.093 | 0.148 | 0.149 | 0.061 | |||||||
| 22 | Eleutheria dichotoma KP776785 | 0.161 | 0.161 | 0.164 | 0.161 | 0.164 | 0.161 | 0.161 | 0.161 | 0.161 | 0.151 | 0.151 | 0.151 | 0.157 | 0.157 | 0.157 | 0.157 | 0.157 | 0.170 | 0.170 | 0.155 | 0.154 | ||||||
| 23 | Staurocladia vallentini KP776806 | 0.186 | 0.186 | 0.190 | 0.186 | 0.190 | 0.186 | 0.186 | 0.183 | 0.183 | 0.158 | 0.158 | 0.154 | 0.160 | 0.160 | 0.160 | 0.160 | 0.160 | 0.170 | 0.164 | 0.157 | 0.154 | 0.069 | |||||
| 24 | Polyorchis haplus AY512549 | 0.193 | 0.193 | 0.197 | 0.193 | 0.197 | 0.193 | 0.193 | 0.200 | 0.200 | 0.180 | 0.180 | 0.181 | 0.178 | 0.178 | 0.178 | 0.178 | 0.178 | 0.203 | 0.210 | 0.189 | 0.160 | 0.158 | 0.190 | ||||
| 25 | Polyorchis penicillatus KX355412 | 0.168 | 0.168 | 0.171 | 0.168 | 0.171 | 0.168 | 0.168 | 0.168 | 0.168 | 0.155 | 0.155 | 0.153 | 0.159 | 0.159 | 0.159 | 0.159 | 0.159 | 0.174 | 0.174 | 0.174 | 0.164 | 0.149 | 0.183 | 0.051 | |||
| 26 | Cladonema radiatum KP776805 | 0.215 | 0.215 | 0.218 | 0.215 | 0.218 | 0.215 | 0.215 | 0.215 | 0.215 | 0.176 | 0.176 | 0.173 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.173 | 0.154 | 0.137 | 0.149 | 0.164 | 0.164 | ||
| 27 | Neoturris pileata MG136759 | 0.202 | 0.202 | 0.205 | 0.202 | 0.205 | 0.202 | 0.202 | 0.202 | 0.202 | 0.189 | 0.189 | 0.186 | 0.189 | 0.189 | 0.189 | 0.189 | 0.189 | 0.206 | 0.200 | 0.189 | 0.179 | 0.198 | 0.212 | 0.177 | 0.170 | 0.176 | |
| 28 | Pandea conica MG136760 | 0.196 | 0.196 | 0.199 | 0.196 | 0.199 | 0.196 | 0.196 | 0.196 | 0.196 | 0.185 | 0.185 | 0.179 | 0.185 | 0.185 | 0.185 | 0.185 | 0.185 | 0.196 | 0.190 | 0.173 | 0.170 | 0.185 | 0.189 | 0.176 | 0.167 | 0.154 | 0.053 |
Table 3.
Morphometrics (mm) of Octorhopalona saltatrix. * The holotype. Nos. KBF-M30-32 and NSMT-Co1802-1805 are paratypes. UH, umbrella height; UD, umbrella diameter.
Table 3.
Morphometrics (mm) of Octorhopalona saltatrix. * The holotype. Nos. KBF-M30-32 and NSMT-Co1802-1805 are paratypes. UH, umbrella height; UD, umbrella diameter.
| Specimen No. | UH (mm) | UD (mm) | Sampling Site | Date | Lat. Long. |
|---|---|---|---|---|---|
| INM-1-96244 * | 6.4 | 5.8 | Oarai Fishing Port, Oarai, Ibaraki, Japan | 15 December 2008 | 36°18′39.1″ N, 140°34′39.5″ E |
| KBF-M30 | 4.8 | 4.7 | Enoshima, Fujisawa, Kanagawa, Japan | 7 May 2021 | 35°17′52.4″ N, 139°28′32.2″ E |
| KBF-M31 | 4.7 | 4.2 | Enoshima, Fujisawa, Kanagawa, Japan | 24 September 2018 | 35°17′52.4″ N, 139°28′32.2″ E |
| KBF-M32 | 2.4 | 2.9 | Shimonokae Fishing Port, Tosashimizu, Kochi, Japan | 30 November 2020 | 32°51′42.96″ N, 132°57′34.80″ E |
| NSMT-Co1802 | 6.4 | 5.5 | Katase Fishing Port, Fujisawa, Kanagawa, Japan | 14 July 2019 | 35°18′23.6″ N 139°28′51.6″ E |
| NSMT-Co1803 | 6.0 | 5.5 | Enoshima, Fujisawa, Kanagawa, Japan | 9 November 2018 | 35°17′52.4″ N, 139°28′32.2″ E |
| NSMT-Co1804 | 9.2 | 8.9 | Enoshima, Fujisawa, Kanagawa, Japan | 22 November 2019 | 35°17′52.4″ N, 139°28′32.2″ E |
| NSMT-Co1805 | 5.0 | 4.4 | Enoshima, Fujisawa, Kanagawa, Japan | 14 July 2019 | 35°17′52.4″ N, 139°28′32.2″ E |
Table 4.
Cnidomes of Octorhopalona saltatrix. D, L represent capsule diameter and length, respectively, in μm.
Table 4.
Cnidomes of Octorhopalona saltatrix. D, L represent capsule diameter and length, respectively, in μm.
| Part | Type | Min | Max | Mean | SD | N | |
|---|---|---|---|---|---|---|---|
| Exumbrella | Stenotele | D | 4.8 | 7.4 | 6.0 | 0.6 | 31 |
| L | 6.5 | 10.5 | 8.6 | 1.1 | 31 | ||
| Manubrium | Desmoneme | D | 2.4 | 3.4 | 2.9 | 0.3 | 8 |
| L | 5.2 | 10.2 | 6.7 | 1.6 | 8 | ||
| Stenotele | D | 5.4 | 7.3 | 6.0 | 0.7 | 7 | |
| L | 7.0 | 12.7 | 9.3 | 1.9 | 7 | ||
| Tentacle | Desmoneme | D | 3.6 | 5.6 | 4.7 | 0.6 | 12 |
| L | 7.7 | 14.8 | 12.0 | 2.5 | 12 | ||
| Stenotele | D | 4.2 | 10.2 | 5.7 | 1.2 | 20 | |
| L | 7.1 | 13.8 | 9.1 | 1.5 | 20 | ||
| Tentacle bulb | Desmoneme | D | 3.7 | 5.4 | 4.4 | 0.7 | 5 |
| L | 9.8 | 14.1 | 11.7 | 1.6 | 5 | ||
| Stenotele | D | 3.7 | 5.1 | 4.4 | 0.4 | 14 | |
| L | 5.3 | 6.5 | 5.8 | 0.3 | 14 |
Table 5.
Morphology of Halimedusidae medusa in previous and the present studies.
| Octorhopalona gen. nov. | Halimedusa | Tiaricodon | Urashimea | |
|---|---|---|---|---|
| UH/UD (mm) | 9/9 | 16/13 | 25/24 | 17/- |
| No. of radial canals | 8 | 4 | 4 | 4 |
| No. of peaks in subumbrella | 8 | 4 | 4 | many |
| No. of tentacles | 4 perradial and 4 interradial tentacles | 4 perradial tentacles and 4 clusters (up to 10–11) of interradial tentacles | 4 perradial tentacles | 4 perradial tentacles |
| Tentacle shape | Stalked nematocyst knobs | Moniliform | Moniliform | Stalked nematocyst knobs |
| Ocellus on tentacle bulbs | Present | Present | Present | Present |
| Umbrella | Bell-shaped | Bell-shaped | Bell-shaped | Bell-shaped or globular |
| Nematocyst tracks | Present | Absent | Absent | Present |
| Peduncle | Absent | Broad and low | Short, broad or indistinct | Absent |
| Red band on manubrium | Present | Present | Present | Present |
| Gonad | On manubrium with lobes extending out onto radial canals, as undulating swollen sacs | On manubrium with smooth lobes extending out onto radial canals (not folded or pendant sacs) | On manubrium with lobes extending out onto radial canals as swollen pouch-like sacs | On manubrium with lobes extending out onto radial canals, as undulating swollen sacs |
| Nematocysts | Stenoteles, desmonemes | Stenoteles, desmonemes, microbasic mastigophores, isorhizas or anisorhizas | Stenoteles, desmonemes, heteronemes | Unknown |
| Distribution | Off of Oarai; Sagami Bay; Tosa Bay, Japan | The Queen Charlotte Islands and the west coast of Vancouver Island in British Columbia; bays on the outer coast of Oregon; northern to central California | Sagami Bay, Japan; Falkland Islands, Wellington Harbor, New Zealand; Yellow Sea, Taiwan Strait, South China Sea, Xiamen Harbor; Cananeia-Iguape Coastal System, Sao Paulo, Brazil | Central to northern Japan, including Hokkaido, and on Sakhalin Island; Amoy, China; St. Helena Bay, South Africa |
| References | This study | [1,9] | [3,4,5,6,26,27,28] | [7,8,29,30,31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Toshino, S.; Yamamoto, G.; Saito, S. Octorhopalona saltatrix, a New Genus and Species (Hydrozoa, Anthoathecata) from Japanese Waters. Animals 2022, 12, 1600. https://doi.org/10.3390/ani12131600
AMA Style
Toshino S, Yamamoto G, Saito S. Octorhopalona saltatrix, a New Genus and Species (Hydrozoa, Anthoathecata) from Japanese Waters. Animals. 2022; 12(13):1600. https://doi.org/10.3390/ani12131600
Chicago/Turabian StyleToshino, Sho, Gaku Yamamoto, and Shinsuke Saito. 2022. "Octorhopalona saltatrix, a New Genus and Species (Hydrozoa, Anthoathecata) from Japanese Waters" Animals 12, no. 13: 1600. https://doi.org/10.3390/ani12131600
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.