
Holoplanktonic and Meroplanktonic Larvae in the Surface Waters of the Onnuri Vent Field in the Central Indian Ridge


1
Risk Assessment Research Center, Korea Institute of Ocean Science and Technology, Geoje 53201, Korea
2
Department of Ocean Science, University of Science and Technology, Daejeon 34113, Korea
3
Marine Ecosystem Research Center, Korea Institute of Ocean Science and Technology, Busan 49111, Korea
*
Author to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2022, 10(2), 158; https://doi.org/10.3390/jmse10020158
Submission received: 23 December 2021
/
Revised: 15 January 2022
/
Accepted: 19 January 2022
/
Published: 26 January 2022
(This article belongs to the Special Issue Adaptation of Marine Animals to Extreme Environments)
Abstract
:The dispersal of organisms in an isolated environment of a hydrothermal vent remains unclear. Here, we provide direct evidence that meroplanktonic larvae may migrate thousands of meters above the ocean floor. The morphological quantitative measurements of mesozooplankton were conducted in the Onnuri Vent Field (OVF), the Solitaire Fields (SF), and the reference site (ref-site). Only one species of bivalve larva that appeared at the OVF and the ref-site (0–200 m) was similar to Bathymodiolus spp. Sixteen species of gastropod larvae were distinguished, among which, species 1–4, 6, and 13 had holoplanktonic features (Atlanta, Oxygrus, and Limacina), whereas species 5, 7–12, and 14–16 had meroplanktonic features. Species 5, 11, and 12 appeared only at the OVF, 9 and 10 appeared only in the SF, 14–16 appeared only at the ref-site, and species 7 and 8 appeared in all surveyed stations. The species 5, 8, 12, 14, and 15 have morphological features similar to Vetulonia spp., and species 7 was similar to Lepetodrilidae; species 9–11 and 16 were similar to Phymorphynchus protoconchs. The morphologically distinguished mollusk larvae in the upper layers of the water column (0–200 m) indicate that larvae associated with deep-sea hydrothermal vents may disperse approximately 2000 m above the vents.
1. Introduction
Benthic animals produce planktotrophic larvae, which spend weeks to months in the pelagic environment, feeding and developing through their free-swimming stage [1,2,3,4]. Larval dispersal is important for geographic distribution, population dynamics, and evolutionary processes of vent-endemic organisms [1,5,6]. Planktonic larval duration (PLD) is the amount of time larvae take to develop to the settlement stage [5]. PLD and dispersal strategies may differ depending on the species, climate zone, and habitat types [7]. In the near-shore ecosystems, the species living on sandy bottoms have long-distance dispersal mechanisms, whereas the rocky-shore species that depend on a narrow strip of habitat have short-distance dispersal mechanisms to reduce offshore loss of larvae [8]. Shanks (2009) reported that shallow-water and coastal species are more concentrated in complex hydrodynamic and fast-speed currents than in the deep-sea. The intertidal and shallow subtidal species are transported from inshore waters into large-scale coastal currents and then eventually return to the inshore waters of another location [9]. Deep-sea species also have complicated PLD, where larval transport distance depends on the development, behavior, and physiology of the larvae [5]. For example, veliger larvae such as limpets (e.g., Leptodrilus spp.) tend to remain near the bottom and in slower currents [10]; tubeworms (e.g., Riftia pachyptila) are buoyant and more likely to be transported above the bottom (e.g., ridge-controlled currents) [11]; and swimming larvae such as mussels (e.g., Bathymodiolus childressi) migrate up into the oceanic currents near the surface [12].
The hydrothermal vent fields have distinct and isolated communities with chemosynthetic systems [13]. The majority of hydrothermal vent sites, particularly in the Pacific and Atlantic Oceans, have been studied [14,15]. A hydrothermal vent has either a discrete or a diffuse venting source, such as black and white smokers [16,17,18]. Another type of vent field is the diffused flow from porous surfaces or fissures and cracks in basalt lavas, diluted with cold seawater either from below or within the sea floor [19]. The benthic animals inhabiting the hydrothermal vent ecosystems are likely to have colonization abilities, including high rates of dispersal, growth, and reproduction [20]. The major vent fauna in the extensively studied Galapagos vents, including tubeworms (Alvinellidae), clams (e.g., Calyptogena magnifica), and mollusks (e.g., B. thermophilus), have symbiotic bacteria that obtain energy from hydrogen sulfide [21].
Most species in hydrothermal vents are benthic as adults, but they spend a portion of their lives as tiny larvae (meroplankton) in the overlying water column [5]. Previous studies on zooplankton, including meroplankton, in the water column above the hydrothermal vents were conducted in the East Pacific Rise (EPR), Guaymas Basin, Endeavour Ridge, Juan de Fuca Ridge, and Mid-Atlantic Ridge [22,23,24,25,26]. The larval stage (meroplankton) of macrobenthic animals living near the hydrothermal vent is an important phase of their life cycle [20,27]. The larval dispersal, especially migration behaviors of benthic organisms in hydrothermal vents has primarily been studied at EPR and Juan de Fuca Ridge [28,29,30,31]. Hydrodynamics and chemical and mass flux studies in the EPR demonstrated the transport mechanisms of vent fluids and larvae, implying eddy-driven impacts in transporting hydrothermal vent-derived heat, chemicals, and biota in a relatively low-energy environment [32].
Meroplanktonic larvae of mollusks in the cold methane seeps, including mytilid bivalve B. childressi and limpet gastropod Bathynerita naticoidea, have been collected from euphotic zones (0–100 m depth), implying that they migrated to surface waters above the ocean floor [33]. Yahagi et al. (2017) provided evidence of early life history traits and population genetics for the surface dispersal of vent species of the gastropod Shinkailepas myojinensis that inhabited sulfide chimneys and volcanic rocks covered by bacterial mats in diffuse venting areas. The hatched larvae swam upwards depending on temperature, and this migration was probably because of the high food source (high phytoplankton biomass) that helps in their growth and development [6].
In this study, we attempted to present the vertical distribution of the holoplanktonic and meroplanktonic larvae in the hydrothermal vents of the Central Indian Ridge. To determine the difference in distribution patterns of larvae from the deep to the sea surface in the diffuse type (Onnuri Vent Field, OVF) and chimney type vent (Solitaire Field, SF), we collected larvae using a multiple opening and closing net environmental sensing system (MOCNESS).
2. Materials and Methods
2.1. Study Area and Physicochemical Analysis
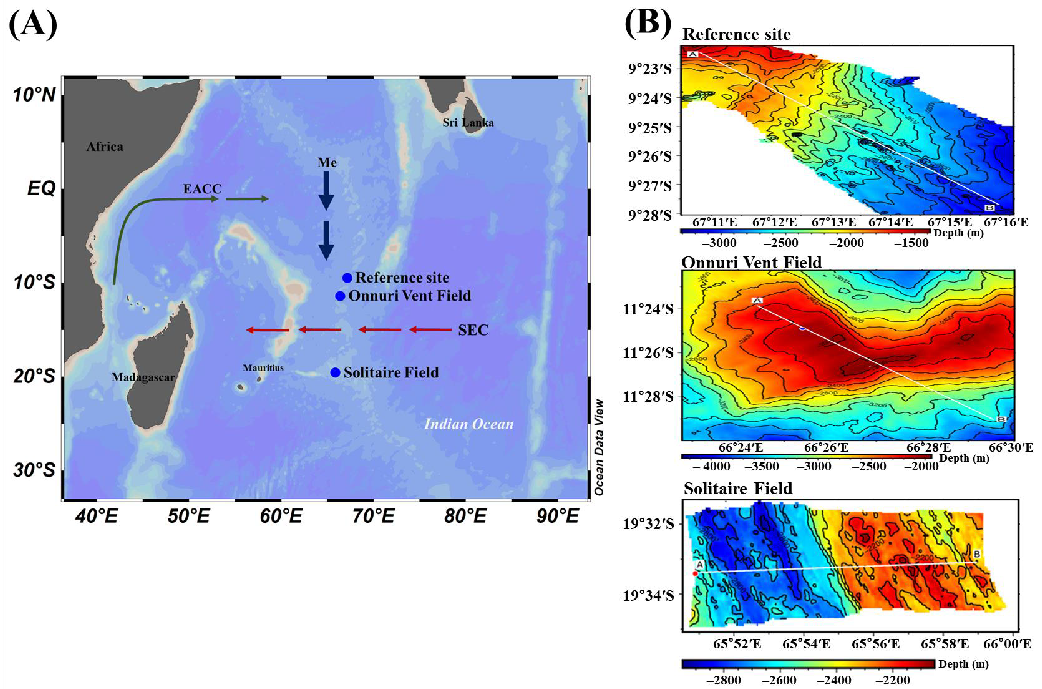
The OVF (newly discovered; 11°23.86′ S, 66°24.42′ E) and SF (19°33.4′ S, 65°50.9′ E) were the two vent fields selected in the Central Indian Ridge (CIR) to be investigated during a cruise of the R/V ISABU in June 2018 (Figure 1A,B). The CIR is known to be slow to intermediate spreading, with spreading rates varying from <12 mm/year to 50–60 mm/year [34]. The OVF is located at the summit of the Ocean Core Complex-3-2 and vents low-temperature fluids at depths of 1990–2170 m [35]. The SF is located on the Roger plateau on the western ridge of CIR segment 15 at a depth of 2634 m [15,34]. SF is characterized by chimneys that are less than 5 m in height and with fluid emissions of high pH 4.8 and approximately 10% lower chlorine levels than the ambient seawater [15]. The reference site (ref-site, MOCNESS tow: 09°22.4′–9°27.7′ S, 67°10.8′–67°15.8′ E) was selected outside of the immediate influence of the hydrothermal vent fields. The ideal location was north of OVF as the direction of transport of meridional Ekman flows from north to south during the boreal summer monsoon in June [36].
The physical properties were obtained from the conductivity (#217), temperature (#6120), and depth (#217) sensors (Sea-bird Electronics, Bellevue, WA, USA) attached to MOCNESS (Biological environmental sampling systems Inc., BESS, North Falmouth, MA, USA). PVC Niskin bottles mounted on an instrumented Rosette sampler were used to collect discrete seawater samples for chlorophyll-a (chl-a) concentration analysis. Due to limited sampling time, water samples for chl-a concentration analysis were not collected at the ref-site. Seawater samples (2 L) for analysis of chl-a concentration were collected at 5 depths, including surface and subsurface chlorophyll maximum (SCM) layers, from 0–0.5 m (surface) to 200 m at OVF and SF. Seawater samples were filtered using 47 mm GF/F glass-fiber filters (Whatman, GE Healthcare, Chicago, IL, USA) under low vacuum pressure (<125 mmHg). The filters were kept frozen in a deep freezer (−80 °C) before being extracted in 90% acetone at 4 °C in the dark for 24 h. Chl-a concentration was determined using Turner Designs 10-AU, which was calibrated using a chl-a standard solution (Turner Designs, San Jose, CA, USA) based on the method by Parsons et al. (1984) [37].
2.2. Mesozooplankton Including Larvae Sampling
The mesozooplankton samples were collected using a 1 m2 MOCNESS equipped with nine nets (mesh size 200 µm). To minimize sample contamination, net traps and bar stops were employed to restrict net bar movement when the nets were opened or closed. The system carried sensors to measure pressure, temperature, and conductivity (Sea-Bird Electronics, Bellevue, WA, USA). The MOCNESS was attached to the deep-tow winch on the R/V ISABU, and the speeds of the winch wire during pay-out and haul-in were 20–30 m min−1. The speed of the vessel was between 1.5 and 2.0 knots during towing. The net angle was maintained between 35° and 55° (optimum angle: 45°; Biological Environmental Sampling Systems Inc., BESS, North Falmouth, MA, USA) by increasing or decreasing the speed of either the ship or the winch wire. Full depth sample collection was obtained by oblique tows from 100 m above the bottom of the sea. The depth strata sampled were divided into 6–7 layers (Table 1). The volume of filtered water was calculated using a flowmeter mounted on the frame of MOCNESS (Biological Environmental Sampling Systems Inc., BESS, North Falmouth, MA, USA). Mesozooplankton samples in the cod-end bucket were transferred to 1 L sampling bottles. The samples were divided into two groups, one sample group was immediately fixed to a final concentration of 5% with borax-neutralized formalin for microscopic examination, and the other was fixed with ethyl-alcohol (99.9%) for further molecular analysis. Mesozooplankton were identified into the lowest taxonomic groups, generally species or genus, and identified and enumerated under a stereomicroscope at 10×–80× magnification (Discovery V8, SteREO, Zeiss, Germany) in accordance with the studies of Conway et al. (2003) [38], and Chihara and Murano (1997) [39]. The larvae of benthic invertebrates at the hydrothermal vents were identified using the photographic identification guide to larvae at hydrothermal vents by Mills et al. (2009) [27]. The gastropod larvae were distinguished and divided into holoplanktonic and meroplanktonic larvae by their morphological characteristics, which were analyzed using the photographic guide. The photographs of mollusks were taken by the stereomicroscope equipped with the camera at 25×, 32×, 50×, 63×, and 80× magnifications (AxioCam ICc 3, Zeiss, Germany). The number of individuals was converted to individuals per cubic meter (ind. 100 m−3). The species richness (Margalef species richness index) and diversity index (Shannon-Wiener diversity index) were calculated by using PRIMER V6.1.10 statistical package (PRIMER-E, Plymouth Marine Laboratory, Plymouth, United Kingdom).
3. Results
3.1. Physicochemical Properties and Chlorophyll-a Concentrations
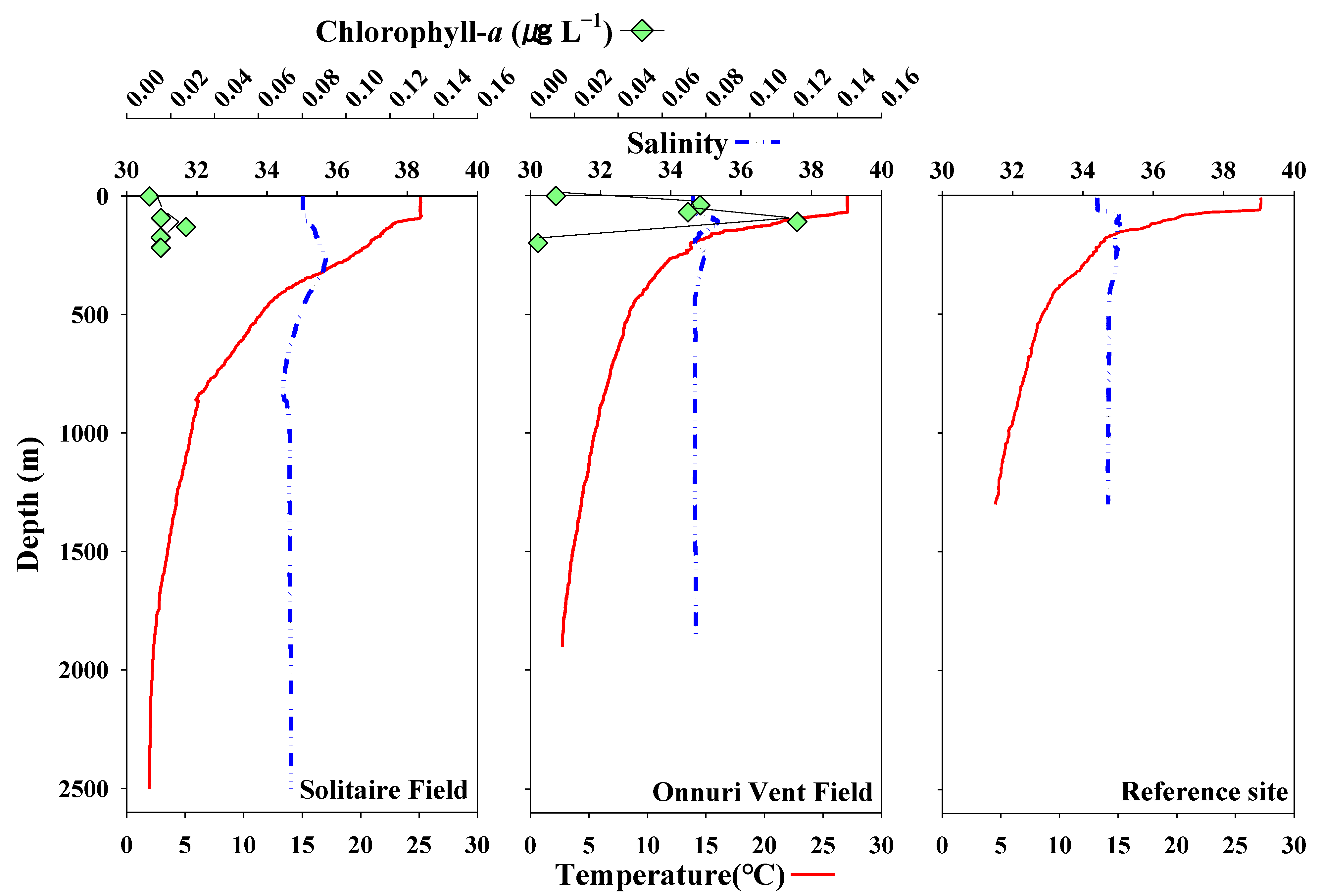
The vertical distributions of temperature and salinity in the water columns were acquired at all survey stations. The temperature observation showed that the depth of the surface mixed layer (SML) increased with an increase in latitude (°S) (avg. 79.2 m) (Figure 2). The deepest SML depth was observed in the SF (100 m; with an average temperature of 25.1 °C), and the shallowest SML depth was seen in the ref-site (63.0 m; with an average temperature of 27.2 °C). The SML depth for OVF was 74.5 m, with an average temperature of 27.0 °C. Warmer surface waters (avg. 27.2 °C) were observed in the ref-site and OVF, while relatively cooler surface water (avg. 25.1 °C) was observed in the SF. The temperature stratification in the SF and OVF was stronger from the surface to 350 m, and weakly stratified with decreasing depth. At SF and OVF, the bottom temperature was 2.7 °C (1900 m) and 4.5 °C (1300 m), respectively. The thermocline (temperature decreases rapidly from the SML) existed between 100–900 m, below which the temperature remained constant (avg. 3.2 °C) and reached 1.9 °C near the bottom (2500 m). The surface water in SF was relatively more saline (avg. 35.0) than the surface waters in OVF and ref-site (avg. 34.5). The salinity remained constant from surface to SML with increasing depth in all surveys, except between a layer of SML and 100 m. The integrated chl-a concentration within the SML was also higher in OVF (4.0 mg m−2) than in SF (1.8 mg m−2) (Figure 2).
3.2. Abundance and Taxonomic Groups of Mesozooplankton
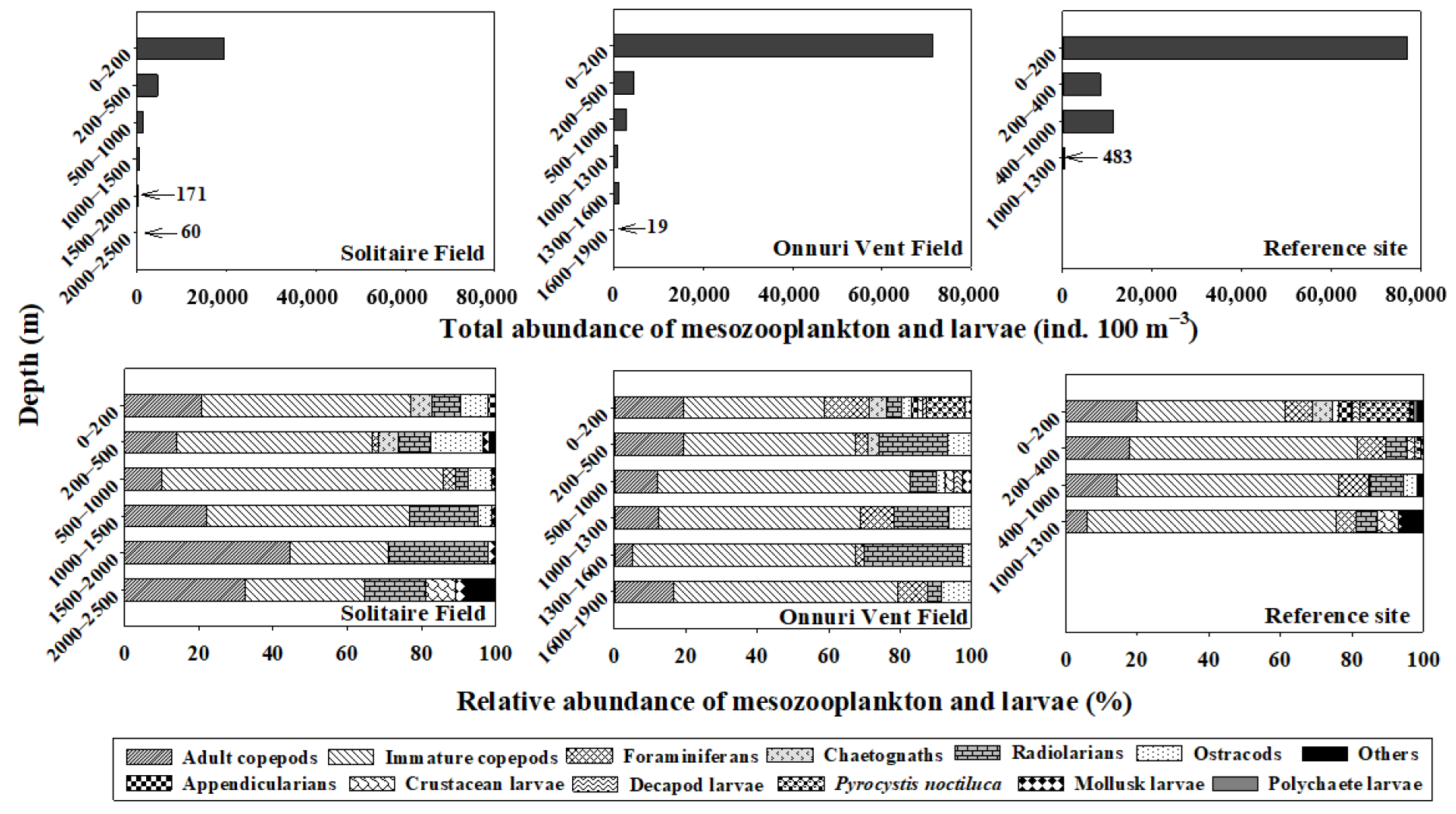
The total mesozooplankton abundance decreased with the depth, ranging from 60 to 19,522 ind. 100 m−3 (avg. 4348 ind. 100 m−3) in SF; 19 to 71,308 ind. 100 m−3 (avg. 13,452 ind. 100 m−3) in OVF; and 483 to 77,037 ind. 100 m−3 (avg. 24,352 ind. 100 m−3) in the ref-site (Figure 3). It is evident that the majority of the mesozooplankton abundance is accumulated at 0–200 m (euphotic zone) in both water columns above hydrothermal vent fields, which may be due to higher chl-a concentrations in the SCM (avg. 112.5 m). The abundance at 0–200 m was approximately 11 times higher than in other layers (200–1300 m) at the ref-site.
Most of the mesozooplankton community was composed of copepods (immature and adults) for all water columns of the surveys (Figure 3, Table 1). At the SF, copepods (both immature and adults) accounted for 65–86% (avg. 74%) of the total mesozooplankton community, followed by radiolarians that accounted for 3–27% (avg. 13%) occurring throughout the water column (0–2500 m), and ostracods and chaetognaths primarily occurring in the upper layers (0–500 m). Taxonomic groups of crustacean larvae and others included amphipod larvae and unidentified eggs, which appeared in the bottom layers (2000–2500 m). At the OVF, copepods (both immature and adults) accounted for 59–83% (avg. 71%) of the total mesozooplankton community, followed by radiolarians (avg. 13%), foraminiferans (avg. 6%), and ostracods (avg. 5%), occurring in all water columns. At the ref-site, copepods (immature and adult) accounted for 61–81% (avg. 74%) of the total mesozooplankton community, followed by foraminiferans (7%), radiolarians (5%), and Pyrocystis noctiluca (4%), occurring in all water columns. P. noctiluca accounted for 11% and 14% of the total mesozooplankton community at OVF and the ref-site, respectively, occurring between 0–200 m, probably due to relatively warmer surface waters (avg. 27.2 °C) (Figure 3, Table 1).
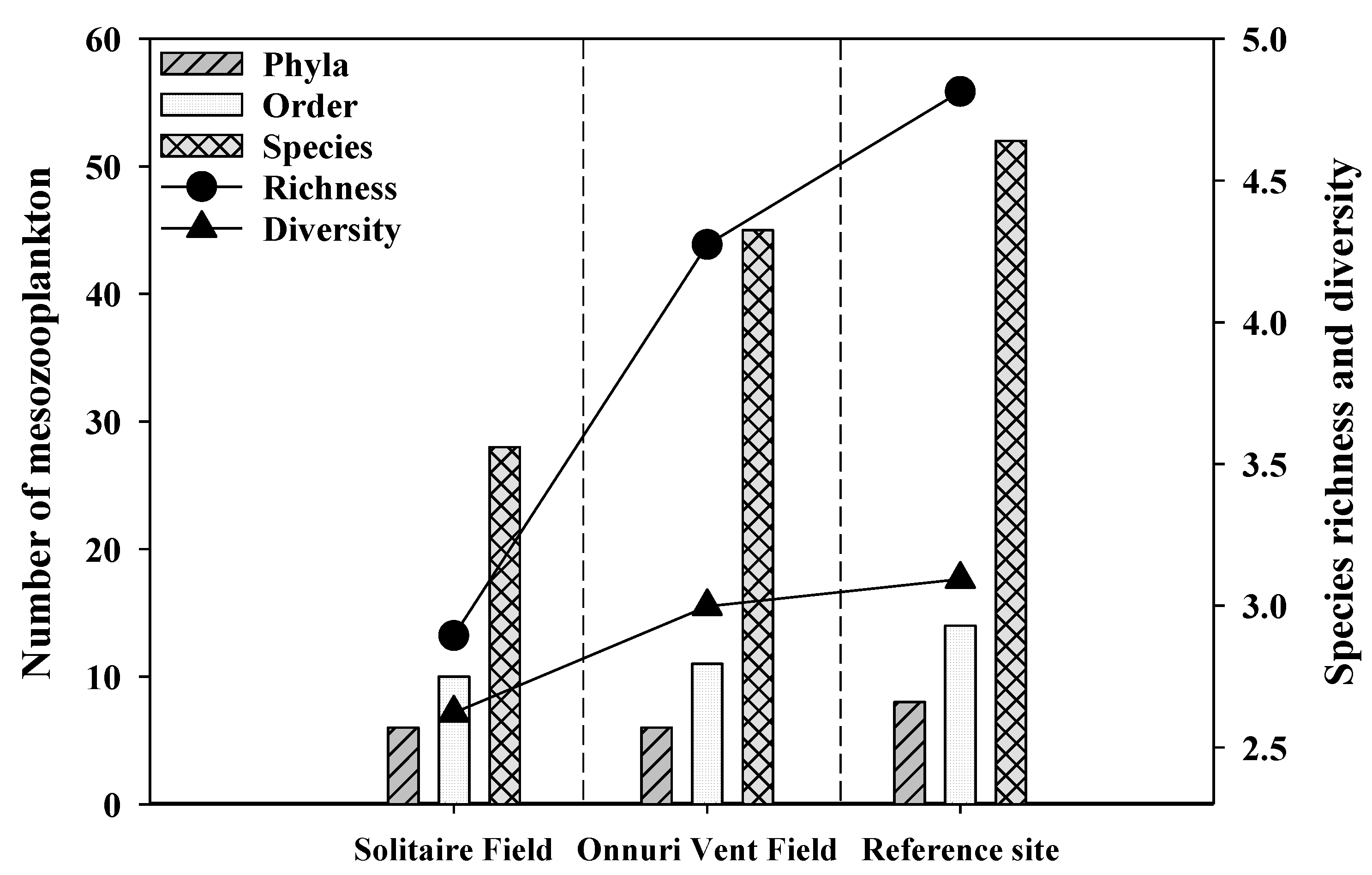
The morphologically identified numbers in the integrated water column were 28, 45, and 52 species in 6, 6, and 8 phyla at SF, OVF and the ref-site, respectively (Figure 4). The phyla Retaria (Foraminifera), Protozoa (Dinoflagellata), Crustacea (Ostracoda, Calanoida, Cyclopoida, Poecilostomatoida, Harpacticoida, Amphipoda, Euphausiacea, Decapoda), Chaetognatha (Sagittoidae), Chordata (Thaliacea, Doliolida, Appendicularia, Ichtyoplankton), and Mollusca (Gastropoda, Bivalvia) commonly appeared at both hydrothermal vents and the ref-site, while phyla Cnidaria (Hydrozoa) and Annelida (Polychaeta) only appeared at the ref-site (Figure 3 and Figure 4). Both the richness and diversity of the species were highest at the ref-site (richness: 4.8, diversity: 3.1) and lowest at SF (richness: 2.9, diversity: 2.6).
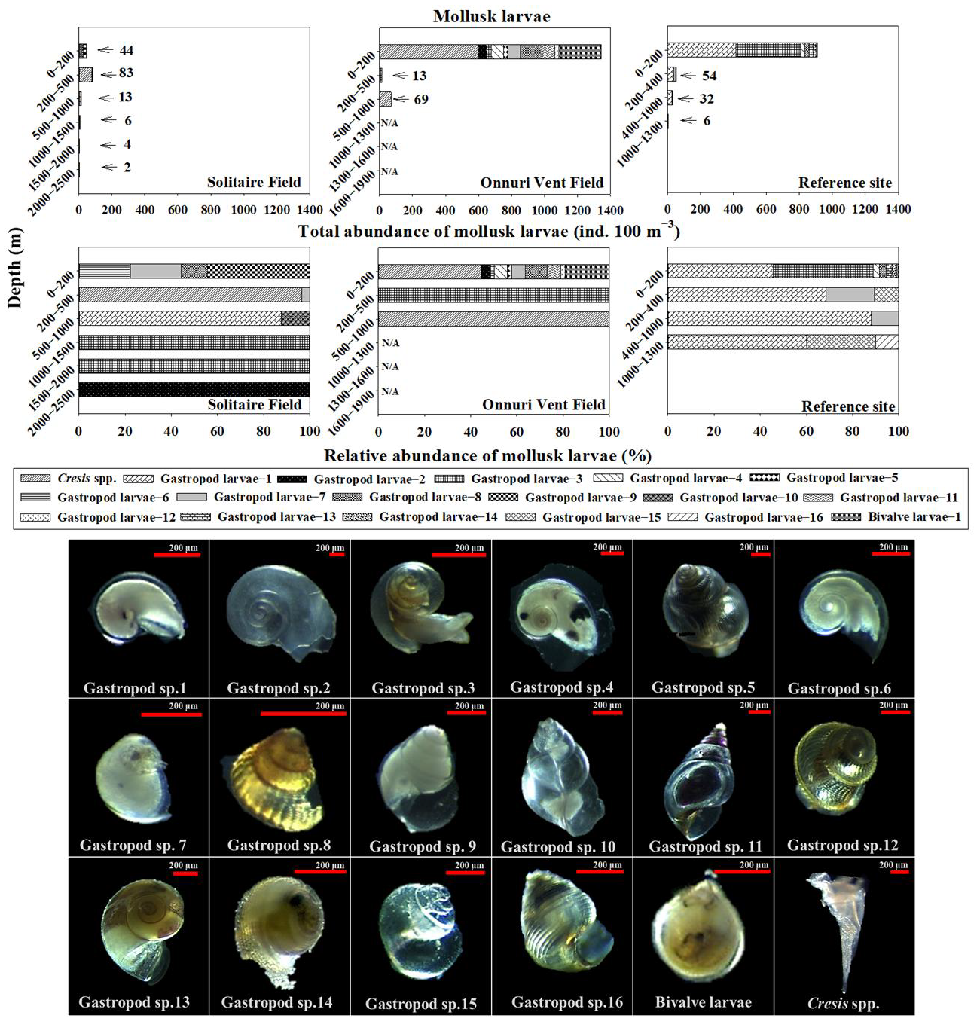
Gastropod larvae were the dominant larvae at both SF (avg. 88%) and OVF (avg. 56%) (Figure 3), followed by amphipod larvae (avg. 12%) at SF and decapod larvae (avg. 17%) at OVF (Figure 3). Copepod nauplii (avg. 49%) was the most dominant larval group at the ref-site, which was followed by gastropod larvae (avg. 27%). We examined the taxonomic groups of mollusk larvae in further detail because deep-sea hydrothermal vent communities may disperse through planktotrophic modes (Figure 5) [1]. Mollusk larvae (bivalves and gastropods) can be holoplankton (spend entire life as plankton) or meroplankton (spend part of their life as plankton). We identified 18 species of mollusk larvae, including 16 unknown species of gastropod larvae, one species of bivalve larvae, and one species of pteropod larvae (Cresis spp.) (Figure 5).
The bivalve larvae (length: 330 µm) were found at both the OVF and the ref-site between depths of 0–200 m (Figure 5). The bivalve larvae had morphological features similar to those of the planktotrophic larvae of Bathymodiolus spp. that were reported to be found at hydrothermal vents in the South Pacific Ocean [27,40]. Gastropod larval species of 1–4, 6, and 13 have morphological features of holoplanktonic larvae such as pelagic species of the genus Atlanta, Oxygrus, and Limacina [38]. Gastropod larval species of 5, 7–12, and 14–16 have morphological features of meroplankton but only appear at depths between the surface and 200 m (Figure 5). Gastropod larval species 5, 11, and 12 only appeared at the OVF; 9 and 10 only appeared at the SF; 14–16 appeared only at the ref-site; 7–8 appeared in all surveyed stations. Gastropod larval species 5, 8, 12, 14, and 15 have similar morphological features to macrobenthic gastropod larvae of Vetulonia spp. Gastropod larvae species 7 have morphological features similar to gastropod Lepetodrilidae (Lepetodrilus spp.). Gastropod larval species 9–11 and 16 have morphological features similar to those of Phymorphynchus protoconchs [27].
4. Discussion
The abundance of mesozooplankton decreased with depth and was mainly accumulated in the euphotic zone (0–200 m). In this study, the near-bottom samples were approximately 0.02% and 0.3% of the total abundance in the euphotic zone of the SF and OVF, respectively. Similar results were reported in the waters of the Guaymas Basin hydrothermal vents, where the benthopelagic zooplankton biomass was about 0.02–0.08% of the surface biomass, and the biomass of zooplankton was 10 times lower in 100–200 m of the bottom than in the surface waters [22,23]. The results of the present study demonstrated that the major accumulation of mesozooplankton was at 0–200 m in the water columns above both the hydrothermal vent fields. Among them, immature copepods of Oithona (28%) and Paracalanus (16%) were dominant in SF and OVF, respectively. This may be influenced by relatively higher temperatures (>23.7 °C) at SF and higher chl-a concentrations in the SCM (avg. 112.5 m) at OVF. The plankton distribution was shown to differ significantly above the hydrothermal vents of the Mid-Atlantic Ridge depending on the productivity of the upper layers [26]. We studied the community structure of the water columns and near-bottom mesozooplankton because the research on the copepods among mesozooplankton in the near-bottom of hydrothermal vents has been limited. The near-bottom dominant copepods were poecilostomatoids at both the OVF (Oncaea and Corycaeus) and the SF (Corycaeus), and harpacticoids appeared in relatively higher abundance at near-bottom of hydrothermal vents (OVF: avg. 16%, SF: avg. 25%). Interestingly, harpacticoids did not appear at the ref-site, implying that harpacticoids may be able to feed on chemosynthetic bacteria near or above hydrothermal vents. The near-bottom copepods are able to feed on free-living chemosynthetic bacteria associated with vent effluents in areas where the effluent signature is weak, playing an important role in the transfer of vent productivity to the deep sea [41]. In addition, in chemosynthetic habitats, harpacticoids were dominant in vent epifauna and seep epifaunal communities, showing a relatively high dominance of copepods within the meiofauna communities [42]. Skebo (2004) studied copepod densities in the near-bottom water layers of the Juan de Fuca (NE Pacific) and reported densities ranging from 0.7–3.5 ind. m−3 over smoker vent and non-vent sites to 14.6 ind. m−3 over diffuse vent sites [41]. Similar results were found at the bottom plankton tows of the Guaymas Basin, in which the copepods of siphonostomes and poecilostomes (avg. 25% of the copepod population) and cyclopoids and harpacticoids were collected in all bottom plankton tows [22].
The near-bottom depth strata range at OVF (1600–1900 m) and SF (1500–2500 m) showed interesting copepod composition, distinguished from the near-bottom depth strata of hydrothermal vents. The community analysis showed that there were distinct shallow (<800 m depth) and deep (>800 m depth) faunal assemblages [24]. The distinct deep faunal assemblages at OVF included Oncaea copepodites (21%), foraminiferans (11%), ostracods (11%), and harpacticoid copepods (11%), and at SF, included radiolarians (22%), Microsetella spp. (22%), and Oncaea copepodites (22%). The copepods can feed directly on bacteria or dissolved organics [24]. There was a possibility of underestimation of the abundance in smaller organisms such as nauplii, copepodites, meroplanktonic larvae and copepod species of Oncaea and Oithona because of the mesh size used (200 μm) for MOCNESS in this study. According to Gallienne and Robbins (2001), an 80 μm net will collect 90% of the total zooplankton abundance and finer mesh nets may result in reduced estimates of larger taxa, and larger mesh (200 μm) are likely only to catch 7% of the total zooplankters between 200 μm and 20 μm in dimension [43].
In this study, radiolarians were the dominant zooplankton throughout the water column above the hydrothermal vent sites (OVF and SF), except at the ref-site. Vereshchaka and Vinogradov (1999) investigated zooplankton aggregations at the MAR Broken Spur, finding that both gelatinous and radiolarians dominate biomass aggregation, significantly contributing to plankton biomass throughout the water column [44]. The radiolarians were dominant near the surface (60–90% of the total biomass), in the lower part of the main pycnocline (10–30% of the total biomass), and near the plume (20–30% of the total biomass) [44]. The speculation as to why radiolarians appear throughout the water column and also near the bottom is that it may be the advantageous feeding through passive absorption of particulate matter [44].
In this study, the holoplanktonic mollusk larvae made up approximately 74%, 61%, and 92% of the total abundance of mollusk larvae in the integrated water columns of SF, OVF, and the ref-site, respectively. The holoplanktonic mollusk larvae including gastropod larval species of 1–4, 6, and 13 in this study had similar morphological features with the genus Atlanta, Oxygrus, and Limacina [38]. The holoplanktonic larvae shells are relatively flat and coiled, and the swimming fins are well developed [21]. According to Lalli and Gilmer (1989) [21], the heteropods have evolved a single swimming fin, whereas pteropods have evolved paired swimming wings, and the feeding mechanisms in open ocean differ from those in the benthic habitat due to the availability of food [21]. The holoplankton are widely distributed in the world’s oceans and usually inhabit the upper ocean of generally offshore regions, where they undertake diurnal vertical migrations [21,45]. Some species of Limacina (ranked among the most abundant gastropods) and other abundant planktonic gastropods may have significant impacts on the epipelagic marine communities [21].
The meroplanktonic mollusk larvae including gastropod larval species of 5, 7–12, and 14–16 and bivalve larvae appeared mostly in the near-surface waters of the euphotic zone (0–200 m depth) of the study sites. Meroplankton, the planktotrophic larvae of benthic organisms, have shells with diamond shaped patterns and relatively large coils as compared with holoplanktonic larvae [27]. The planktotrophic phase is important for larval development because it allows larvae to feed during the free-swimming stage, promoting deep-sea community dispersal, colonization, and resilience [46,47]. The larvae interact with oceanic circulation, ocean ridge topography, and ridge flow and exhibit vertical migration behaviors [5,29]. Depending on the species, they spend hours to several weeks or months dispersing and drifting in the water columns by means of water currents before setting and metamorphosing on the sea floor [21].
The present study showed similar evidence that meroplanktonic mollusk larvae may disperse in the surface waters by the possibility of vertical migration. The bivalve larvae, which appeared in both the OVF and the ref-site, had morphologically similar features to planktotrophic larvae of the deep-sea hydrothermal vent species of Bathymodiolus spp. [27]. According to Mills et al. (2007), they collected Bathymodiolus species (length 400–450 µm) from near the settlements, of brownish color. B. marisindicus (avg. 31.1%) was one of the top-ranked microbenthic species at OVF [35]. However, we cannot be certain that the bivalve larvae are larvae of Bathymodiolus spp. because larvae of coastal mussels (Mytilus spp.) also have similar morphological features, thus further molecular analyses are needed to confirm the species [48,49]. The vent mussel B. marisindicus has been previously reported at the SF and OVF [34,35]. So, the bivalve larvae may have vertically migrated from the hydrothermal vents of the OVF to the surface waters. Arellano et al. (2014) reported cold methane seep species of mytilid bivalve B. childressi in the euphotic zone (0–100 m depth) and suggested that they had migrated to surface waters from the ocean floor [33]. The possibility of bivalve larvae being transported from the coastal, intertidal, or shallow waters maybe low because our study site is in the open ocean, approximately 1000 km away from the coasts. The dispersal of coastal, intertidal, or shallow-water bivalve larvae is through transport from inshore waters into larger scale coastal currents, and then eventually returning to the inshore waters of another location [9].
The gastropod larvae (species 7) that appeared mostly in the euphotic zone (0–200 m) at OVF and SF had morphological similarities to gastropod larvae Lepetodrilidae (Lepetodrilus spp.), which inhabit the hydrothermal vent sites of Juan de Fuca Ridge and EPR [27,44]. The abundance of vent gastropod larvae (Lepetodrilus sp. and two peltospirid species) is significantly higher in the plume than away from it (mean abundance = 21.0 ind. 1000 m−3) [50]. Gastropod larval species 5, 8, 12, 14, and 15 have similar morphological features to macrobenthic gastropod larvae of Vetulonia spp. that were found in the hydrothermal vents and hydrothermal seeps at the North Fiji Basin [51]. Gastropod larval species 9–11 and 16 have morphological features similar to those of Phymorphynchus protoconchs [27]. The vent mollusk fauna in SF includes B. marisindicus (Mitilidae), Lepetodrilus sp. (Lepetodridae), Eulepetopsis sp. (Neolepetopsidae), Scaly-foot gastropod (Peltospiridae), Shinkailepas spp. (Phenacolepadidae), Alviniconcha spp., Desbruyeresia spp. (Provannidae), and Phymorhynchus spp. (Turridae) [34]. The vent mollusk fauna in OVF includes B. marisindicus, B. sp.1, Gigantidas n. spp., Alvinocaris markensis, Paralepetopsis ferrugivora, and Lepetodrilus spp. [35]. Larval dispersal in the planktonic larval stage is an important component of the life cycle of species, particularly in isolated habitats such as hydrothermal vents [28]. Adams et al. (2012) studied and reviewed the reproductive patterns and larval durations, behaviors and vertical distributions, and the coupling of vent topography and hydrodynamics that affect dispersal [5]. Yahagi et al. (2017) provided evidence of early life history traits and population genetics for the surface dispersal of gastropod vent species Shinkailepas myojinensis that inhabited sulfide chimneys and volcanic rocks covered by bacterial mats in diffuse venting areas [6]. The hatched larvae swam upwards at 16.6–44.42 mm/min depending on temperature, and were likely to migrate to surface water because of the high food source (high phytoplankton biomass) that helps in their growth and development [6]. Therefore, in this study, the gastropod larvae in the surface waters may have dispersed or vertically migrated from the deep-sea hydrothermal vent sites (OVF and SF).
5. Conclusions
The quantitative measurements of mesozooplankton including holoplankton and meroplankton were conducted in the OVF, SF, and ref-site. The integrated abundance of mesozooplankton over the water column was relatively higher in the ref-site (77,037 ind. 100 m−3) than in the vent sites (avg. 53,398 ind. 100 m−3). It is evident that most of the mesozooplankton was accumulated in the euphotic zone (0–200 m). Mollusk larvae (gastropods and bivalves) appeared at all stations. Only one species of bivalve larva that appeared at the OVF and ref-site (0–200 m) was morphologically similar to Bathymodiolus spp. that inhabits the hydrothermal vents. The gastropod larvae were morphologically identified as holoplanktonic and meroplanktonic. Sixteen species of gastropod larvae were distinguished morphologically. Gastropod larvae species 1–4, 6, and 13 had holoplanktonic features (Atlanta, Oxygrus, and Limacina), while gastropod species 5, 7–12, and 14–16 had meroplanktonic features. Gastropod larvae species 5, 11, and 12 only appeared at OVF, 9 and 10 only appeared at SF, 4–16 appeared only at ref-site, and gastropod larvae species 7–8 appeared in all surveyed stations. Gastropod larvae species 5, 8, 12, 14, and 15 have similar morphological features to the macrobenthic gastropod larvae of Vetulonia spp. Gastropod larvae species 7 have morphological features similar to gastropods Lepetodrilidae, and species 9–11 and 16 have morphological features similar to Phymorphynchus protoconchs. The morphologically identified mollusk larvae of gastropods and bivalves in the upper layers of the water column (0–200 m) indicate that larvae associated with the hydrothermal vents may disperse approximately 2000 m above the vents.
Author Contributions
Conceptualization, M.K. and J.-H.K.; methodology, M.K. and J.-H.K.; investigation, M.K., J.-H.K. and D.K.; data curation, M.K., J.-H.K.; writing—original draft preparation, M.K., J.-H.K.; writing—review and editing, M.K., J.-H.K.; visualization, M.K., J.-H.K.; project administration, M.K., J.-H.K. and D.K.; funding acquisition, J.-H.K. and D.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was a part of the project titled “Understanding the deep-sea biosphere on seafloor hydrothermal vents in the Indian Ridge (No. 20170411)” funded by the Ministry of Oceans and Fisheries, Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used are presented in the text, figures, and tables.
Acknowledgments
The authors would like to thank the captain and the crews of R/V ISABU for supporting the MOCNESS operations and shipboard works. We thank G. Hwang for bathymetry assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lutz, R.A.; Jablonski, D.; Turner, R.D. Larval development and dispersal at deep-sea hydrothermal vents. Science 1984, 226, 1451–1454. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, P.; Warén, A. Ontogenetic migration and dispersal of deep-sea gastropod larvae. In Reproduction, Larval Biology, and Recruitment of the Deep-Sea Benthos; Young, C.M., Eckelbarger, K.J., Eds.; Columbia University Press: New York, NY, USA, 1994; pp. 98–117. [Google Scholar]
- Keough, M.J.; Swearer, S.E. Early life histories of marine invertebrates and fishes. In Marine Ecology; Connell, S.D., Gillanders, B.M., Eds.; Oxford University Press: Oxford, UK, 2007; pp. 19–46. [Google Scholar]
- Swearer, S.E.; Treml, E.A.; Shima, J.S. A review of biophysical models of marine larval dispersal. In Oceanography and Marine Biology; Hawkins, S.J., Allcock, A.L., Bates, A.E., Firth, L.B., Smith, I.P., Swearer, S.E., Todd, P.A., Eds.; CRC Press: Boca Raton, FL, USA, 2019; Volume 57, pp. 325–356. [Google Scholar]
- Adams, D.K.; Arellano, S.M.; Govenar, B. Larval dispersal: Vent life in the water column. Oceanography 2012, 25, 256–268. [Google Scholar] [CrossRef]
- Yahagi, T.; Kayama Watanabe, H.; Kojima, S.; Kano, Y. Do larvae from deep-sea hydrothermal vents disperse in surface waters? Ecology 2017, 98, 1524–1534. [Google Scholar] [CrossRef] [PubMed]
- Young, C.M.; Arellano, S.M.; Hamel, J.F.; Mercier, A. Ecology and evolution of larval dispersal in the deep sea. In Evolutionary and Ecology of Marine Invertebrate Larvae; Carrier, T.J., Reitzel, A.M., Heyland, A., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 229–245. [Google Scholar]
- Shanks, A.L. Pelagic larval duration and dispersal distance revisited. Biol. Bull. 2009, 21, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Tilburg, C.E.; McCartney, M.A.; Yund, P.O. Across-shelf transport of bivalve larvae: Can the interface between a coastal current and inshore waters act as an ecological barrier to larval dispersal? PLoS ONE 2012, 7, e48960. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, L.S.; Mills, S.W.; Sweetman, A.K.; Beaudreau, A.H.; Metaxas, A.; Hunt, H.L. Vertical, lateral and temporal structure in larval distributions at hydrothermal vents. Mar. Ecol. Prog. Ser. 2005, 293, 1–16. [Google Scholar] [CrossRef]
- Brooke, S.D.; Young, C.M. Where do the embryos of Riftia pachyptila develop? Pressure tolerances, temperature tolerances, and buoyancy during prolonged embryonic dispersal. Deep-Sea Res. Part II 2009, 56, 1599–1606. [Google Scholar] [CrossRef]
- Arellano, S.M.; Young, C.M. Temperature and salinity tolerances of embryos and larvae of the deep-sea mytilid mussel “Bathymodiolus” childressi. Mar. Biol. 2011, 158, 2481–2493. [Google Scholar] [CrossRef]
- Nakamura, K.; Watanabe, H.; Miyazaki, J.; Takai, K.; Kawagucci, S.; Noguchi, T.; Okamura, K. Discovery of new hydrothermal activity and chemosynthetic fauna on the Central Indian Ridge at 18°–20°S. PLoS ONE 2012, 7, e32965. [Google Scholar] [CrossRef] [Green Version]
- Lonsdale, P. Clustering of suspension-feeding macrobenthos near abyssal hydrothermal vents at oceanic spreading centers. Deep Sea Res. 1977, 24, 857–863. [Google Scholar] [CrossRef]
- Nakamura, K.; Takai, K. Indian Ocean Hydrothermal Systems: Seafloor Hydrothermal Activities, Physical and Chemical Characteristics of Hydrothermal Fluids, and Vent-Associated Biological Communities. In Subseafloor Biosphere Linked to Hydrothermal Systems; Ishibashi, J.-I., Okino, K., Sunamura, M., Eds.; Springer: Tokyo, Japan, 2015; pp. 147–161. [Google Scholar]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef]
- Demina, L.L.; Galkin, S.V. Trace Metal Biogeochemistry and Ecology of Deep-Sea Hydrothermal Vent Systems, 1st ed.; Springer: Cham, Switzerland, 2016; Volume 50, pp. 16–17. [Google Scholar]
- Van Dover, C.L.; Arnaud-Haond, S.; Gianni, M.; Helmreich, S.; Huber, J.A.; Jaeckel, A.L.; Metaxas, A.; Pendleton, L.H.; Petersen, S.; Ramirez-Llodra, E.; et al. Scientific rationale and international obligations for protection of active hydrothermal vent ecosystems from deep-sea mining. Mar. Policy 2018, 90, 20–28. [Google Scholar] [CrossRef]
- Trivett, D.A.; Williams, A.J., III. Effluent from diffuse hydrothermal venting: 2. Measurement of plumes from diffuse hydrothermal vents at the southern Juan de Fuca Ridge. J. Geophys. Res. Oceans 1994, 99, 18417–18432. [Google Scholar] [CrossRef]
- Beedessee, G.; Watanabe, H.; Ogura, T.; Nemoto, S.; Yahagi, T.; Nakagawa, S.; Nakamura, K.; Takai, K.; Koonjul, M.; Marie, D.E.; et al. High connectivity of animal populations in deep-sea hydrothermal vent fields in the Central Indian Ridge relevant to its geological setting. PLoS ONE 2013, 8, e81570. [Google Scholar] [CrossRef] [Green Version]
- Lalli, C.M.; Gilmer, R.W. Pelagic Snails: The Biology of Holoplanktonic Gastropod Mollusks, 1st ed.; Stanford University Press: Redwood City, CA, USA, 1989; pp. 1–6. ISBN 9780804714907. [Google Scholar]
- Berg, C.J., Jr.; Van Dover, C.L. Benthopelagic macrozooplankton communities at and near deep-sea hydrothermal vents in the eastern Pacific Ocean and the Gulf of California. Deep Sea Res. Part A Oceanogr. Res. Pap. 1987, 34, 379–401. [Google Scholar] [CrossRef]
- Wiebe, P.H.; Copley, N.; Van Dover, C.; Tamse, A.; Manrique, F. Deep-water zooplankton of the Guaymas Basin hydrothermal vent field. Deep Sea Res. Part I Oceanogr. Res. Pap. 1988, 35, 985–1013. [Google Scholar] [CrossRef]
- Burd, B.J.; Thomson, R.E. Hydrothermal venting at Endeavour Ridge: Effect on zooplankton biomass throughout the water column. Deep Sea Res. Part A Oceanogr. Res. Pap. 1994, 41, 1407–1423. [Google Scholar] [CrossRef]
- Burd, B.J.; Thomson, R.E. Distribution of zooplankton associated with the Endeavour Ridge hydrothermal plume. J. Plankton Res. 1995, 17, 965–997. [Google Scholar] [CrossRef]
- Vinogradov, G.M.; Vereshchaka, A.L.; Aleinik, D.L. Zooplankton distribution over hydrothermal fields of the Mid-Atlantic Ridge. Oceanol. C/C Okeanol. 2003, 43, 656–669. [Google Scholar]
- Mills, S.W.; Beaulieu, S.E.; Mullineaux, L.S. Photographic Identification Guide to Larvae at Hydrothermal Vents in the Eastern Pacific. 2009. Available online: http://www.whoi.edu/science/B/vent-larval-id (accessed on 13 December 2021).
- Kim, S.L.; Mullineaux, L.S.; Helfrich, K.R. Larval dispersal via entrainment into hydrothermal vent plumes. J. Geophys. Res. Oceans 1994, 99, 12655–12665. [Google Scholar] [CrossRef]
- Adams, D.K.; Mullineaux, L.S. Supply of gastropod larvae to hydrothermal vents reflects transport from local larval sources. Limnol. Oceanogr. 2008, 53, 1945–1955. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.L.; Mullineaux, L.S. Distribution and near-bottom transport of larvae and other plankton at hydrothermal vents. Deep Sea Res. Part II Top. Stud. Oceanogr. 1998, 45, 423–440. [Google Scholar] [CrossRef]
- Adams, D.K.; Mills, S.W.; Shank, T.M.; Mullineaux, L.S. Expanding dispersal studies at hydrothermal vents through species identification of cryptic larval forms. Mar. Biol. 2010, 157, 1049–1062. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.K.; McGillicuddy, D.J.; Zamudio, L.; Thurnherr, A.M.; Liang, X.; Rouxel, O.; German, C.R.; Mullineaux, L.S. Surface-generated mesoscale eddies transport deep-sea products from hydrothermal vents. Science 2011, 332, 580–583. [Google Scholar] [CrossRef] [Green Version]
- Arellano, S.M.; Van Gaest, A.L.; Johnson, S.B.; Vrijenhoek, R.C.; Young, C.M. Larvae from deep-sea methane seeps disperse in surface waters. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133276. [Google Scholar] [CrossRef]
- Watanabe, H.; Beedessee, G. Vent fauna on the Central Indian Ridge. In Subseafloor Biosphere Linked to Hydrothermal Systems; Ishibashi, J.-I., Okino, K., Sunamura, M., Eds.; Springer: Tokyo, Japan, 2015; pp. 205–212. [Google Scholar]
- Kim, J.; Son, S.K.; Kim, D.; Pak, S.J.; Yu, O.H.; Walker, S.L.; Oh, J.; Choi, S.K.; Ra, K.; Ko, Y.; et al. Discovery of active hydrothermal vent fields along the Central Indian Ridge, 8–12°S. Geochem. Geophys. Geosyst. 2020, 21, e2020GC009058. [Google Scholar] [CrossRef]
- Hood, R.R.; Beckley, L.E.; Wiggert, J.D. Biogeochemical and ecological impacts of boundary currents in the Indian Ocean. Prog. Oceanogr. 2017, 156, 290–325. [Google Scholar] [CrossRef] [Green Version]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: New York, NY, USA, 1984; pp. 101–112. [Google Scholar]
- Conway, D.V.; White, R.G.; Hugues-Dit-Ciles, J.; Gallienne, C.P.; Robins, D.B. Guide to the Coastal and Surface Zooplankton of the South-Western Indian Ocean. DEFRA Darwin Initiative Zooplankton Programme, Marine Biological Association of the United Kingdom, No. 15; Marine Biological Association of the United Kingdom: Plymouth, UK, 2003; pp. 1–354. ISSN 0260-2784. [Google Scholar]
- Chihara, M.; Murano, M. An Illustrated Guide to Marine Plankton in Japan; Tokai University Press: Hiratsuka, Japan, 1997. [Google Scholar]
- Kenk, V.C.; Wilson, B.R. A new mussel (Bivalvia, Mytilidae) from hydrothermal vents in the Galapagos Rift zone. Malacologia 1985, 26, 253–271. [Google Scholar]
- Skebo, K.M. Distribution of zooplankton and nekton above hydrothermal vents on the Juan de Fuca and Explorer ridges. Master’s Thesis, University of Victoria, Victoria, BC, Canada, 2004. [Google Scholar]
- Gollner, S.; Ivanenko, V.N.; Arbizu, P.M.; Bright, M. Advances in taxonomy, ecology, and biogeography of Dirivultidae (Copepoda) associated with chemosynthetic environments in the deep sea. PLoS ONE 2010, 5, e9801. [Google Scholar] [CrossRef] [PubMed]
- Gallienne, C.P.; Robins, D.B. Is Oithona the most important copepod in the world’s oceans? J. Plankton Res. 2001, 23, 1421–1432. [Google Scholar] [CrossRef]
- Vereshchaka, A.L.; Vinogradov, G.M. Visual observations of the vertical distribution of plankton throughout the water column above Broken Spur vent field, Mid-Atlantic Ridge. Deep Sea Res. Part I Oceanogr. Res. Pap. 1999, 46, 1615–1632. [Google Scholar] [CrossRef]
- Hart, M.B.; Wall-Palmer, D.; Janssen, A.W.; Smart, C.W. Some observations on the geological history of the holoplanktonic gastropods. Proc. Geol. Assoc. 2020, 131, 443–449. [Google Scholar] [CrossRef]
- Lutz, R.A.; Kennish, M.J. Ecology of deep-sea hydrothermal vent communities: A review. Rev. Geophys. 1993, 31, 211–242. [Google Scholar] [CrossRef]
- Baco, A.R.; Etter, R.J.; Ribeiro, P.A.; Von der Heyden, S.; Beerli, P.; Kinlan, B.P. A synthesis of genetic connectivity in deep-sea fauna and implications for marine reserve design. Mol. Ecol. 2016, 25, 3276–3298. [Google Scholar] [CrossRef] [PubMed]
- Sprung, M. Physiological energetics of mussel larvae (Mytilus edulis). I. Shell growth and biomass. Mar. Ecol. Prog. Ser. 1984, 17, 283–293. [Google Scholar] [CrossRef]
- Schönitzer, V.; Weiss, I.M. The structure of mollusc larval shells formed in the presence of the chitin synthase inhibitor Nikkomycin Z. BMC Struct. Biol. 2007, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, L.S.; Wiebe, P.H.; Baker, E.T. Larvae of benthic invertebrates in hydrothermal vent plumes over Juan de Fuca Ridge. Mar. Biol. 1995, 122, 585–596. [Google Scholar] [CrossRef]
- Warèn, A.; Bouchet, P. New records, species, genera, and a new family of gastropods from hydrothermal vents and hydrocarbon seeps. Zoologica Scripta 1993, 22, 1–90. [Google Scholar] [CrossRef]
Figure 1.
(A) Map showing the selected survey stations at the Central Indian Ridge in June 2018. Surface currents indicated are the South Equatorial Current (SEC), East Africa Coastal Current (EACC), and the direction of meridional Ekman transports (Me). The major currents are cited from Hood et al. (2017). (B) The figure on the right shows bathymetry from the start to the end of the multiple opening and closing net environmental sensing system (MOCNESS) sampling in both study stations.
Figure 1.
(A) Map showing the selected survey stations at the Central Indian Ridge in June 2018. Surface currents indicated are the South Equatorial Current (SEC), East Africa Coastal Current (EACC), and the direction of meridional Ekman transports (Me). The major currents are cited from Hood et al. (2017). (B) The figure on the right shows bathymetry from the start to the end of the multiple opening and closing net environmental sensing system (MOCNESS) sampling in both study stations.

Figure 2.
The vertical profiles of temperature (°C), salinity, and chlorophyll-a (µg L−1) of the surveyed stations at the Central Indian Ridge during a cruise of the R/V ISABU in June 2018.
Figure 2.
The vertical profiles of temperature (°C), salinity, and chlorophyll-a (µg L−1) of the surveyed stations at the Central Indian Ridge during a cruise of the R/V ISABU in June 2018.

Figure 3.
The vertical distribution of total and relative abundance of the mesozooplankton community in the surveyed stations during a cruise of the R/V ISABU in June 2018 at the Central Indian Ridge.
Figure 3.
The vertical distribution of total and relative abundance of the mesozooplankton community in the surveyed stations during a cruise of the R/V ISABU in June 2018 at the Central Indian Ridge.

Figure 4.
The number of mesozooplankton phyla, order, and species, and indices of species richness and diversity in the surveyed stations during a cruise of the R/V ISABU in June 2018 at the Central Indian Ridge.
Figure 4.
The number of mesozooplankton phyla, order, and species, and indices of species richness and diversity in the surveyed stations during a cruise of the R/V ISABU in June 2018 at the Central Indian Ridge.

Figure 5.
The vertical distribution of the total and relative abundance of mollusk larvae in the surveyed stations during a cruise of the R/V ISABU in June 2018 at the Central Indian Ridge. The 18 species of mollusk larvae, include 16 unknown species of gastropod larvae, one species of bivalve larvae, and one species of pteropod larvae (Cresis spp.). N/A represents not available.
Figure 5.
The vertical distribution of the total and relative abundance of mollusk larvae in the surveyed stations during a cruise of the R/V ISABU in June 2018 at the Central Indian Ridge. The 18 species of mollusk larvae, include 16 unknown species of gastropod larvae, one species of bivalve larvae, and one species of pteropod larvae (Cresis spp.). N/A represents not available.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The sampling strata (depth, m), and average for temperature (temp.), and salinity (sal.), integrated chlorophyll-a (chl-a), total abundance of zooplankton (zoopl.), and top three dominant zooplankton (zoopl.) are listed for each station during the study period. N.S.: not sampled.
Table 1.
The sampling strata (depth, m), and average for temperature (temp.), and salinity (sal.), integrated chlorophyll-a (chl-a), total abundance of zooplankton (zoopl.), and top three dominant zooplankton (zoopl.) are listed for each station during the study period. N.S.: not sampled.
| Station | Sampling Strata (m) | Temp. (°C) | Sal. | Chl-a (mg m−3) | Total Zoopl. (ind. 100 m−3) | Top Three Dominant Zoopl. (Relative Abundance) |
|---|---|---|---|---|---|---|
| Solitaire Field | (1) 0–200 | 23.7 | 35.2 | 3.1 | 19,522 | Oithona copepodites (28%), Radiolarians (8%), Pleuromamma copepodites (6%) |
| (2) 200–500 | 15.7 | 35.4 | N.S. | 4542 | Pleuromamma copepodites (12%), Oithona copepodites (11%), ostracod sp.1 (11%) | |
| (3) 500–1000 | 7.9 | 34.6 | N.S. | 1291 | Oncaea copepodites (26%), Pleuromamma copepodites (23%), Oithona copepodites (10%) | |
| (4) 1000–1500 | 4.4 | 34.6 | N.S. | 501 | Radiolarians (18%), Oithona copepodites (18%), Oncaea copepodites (11%) | |
| (5) 1500–2000 | 2.7 | 34.7 | N.S. | 171 | Radiolarians (27%), Oncaea copepodites (18%), Oithona fallax (18%) | |
| (6) 2000–2500 | 2.0 | 34.7 | N.S. | 60 | Radiolarians (16%), Oncaea copepodites (16%), Microsetella spp. (16%) | |
| Onnuri Vent Field | (1) 0–200 | 18.3 | 35.0 | 13.5 | 71,308 | Paracalanus copepodites (16%), Pyrocystis noctiluca (11%), foraminiferans (7%) |
| (2) 200–500 | 10.7 | 34.8 | N.S. | 4474 | Radiolarians (19%), Oncaea copepodites (13%), Scolecithrix copepodites (13%) | |
| (3) 500–1000 | 6.9 | 34.7 | N.S. | 2850 | Oncaea copepodites (32%), Harpacticoid copepodites (15%), Oithona copepodites (10%) | |
| (4) 1000–1300 | 4.9 | 34.7 | N.S. | 922 | Oncaea copepodites (25%), Radiolarians (16%), Scolecithrix copepodites (13%) | |
| (5) 1300–1600 | 3.8 | 34.7 | N.S. | 1138 | Oncaea copepodites (43%), Radiolarians (28%), Harpacticoid copepodites (8%) | |
| (6) 1600–1900 | 3.0 | 34.7 | N.S. | 19 | Oncaea copepodites (42%), Harpacticoid copepodites (13%), foraminiferans (8%) | |
| Reference site | (1) 0–200 | 20.9 | 34.8 | N.S. | 77,037 | Pyrocystis noctiluca (14%), Globigerina spp. (7%), Oncaea copepodites (7%) |
| (2) 200–400 | 11.8 | 34.9 | N.S. | 8430 | Oithona copepodites (18%), Oncaea copepodites (14%), Pleuromamma copepodites (6%) | |
| (3) 400–1000 | 7.4 | 34.7 | N.S. | 11,460 | Oncaea copepodites (25%), Oithona copepodites (12%), Scolecithrix copepodites (10%) | |
| (4) 1000–1300 | 5.1 | 34.7 | N.S. | 483 | Oncaea copepodites (46%), Oithona copepodites (12%), Scolecithrix copepodites (6%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, M.; Kang, J.-H.; Kim, D. Holoplanktonic and Meroplanktonic Larvae in the Surface Waters of the Onnuri Vent Field in the Central Indian Ridge. J. Mar. Sci. Eng. 2022, 10, 158. https://doi.org/10.3390/jmse10020158
AMA Style
Kim M, Kang J-H, Kim D. Holoplanktonic and Meroplanktonic Larvae in the Surface Waters of the Onnuri Vent Field in the Central Indian Ridge. Journal of Marine Science and Engineering. 2022; 10(2):158. https://doi.org/10.3390/jmse10020158
Chicago/Turabian StyleKim, Minju, Jung-Hoon Kang, and Dongsung Kim. 2022. "Holoplanktonic and Meroplanktonic Larvae in the Surface Waters of the Onnuri Vent Field in the Central Indian Ridge" Journal of Marine Science and Engineering 10, no. 2: 158. https://doi.org/10.3390/jmse10020158
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.