Small Jellyfish as a Supplementary Autumnal Food Source for Juvenile Chaetognaths in Sanya Bay, China
by
Lingli Wang
1,2,3, 
Minglan Guo
1,3,
Tao Li
1,3,4,
Hui Huang
1,3,4,5,
Sheng Liu
1,3,* and
Simin Hu
1,3,* 1
CAS Key Laboratory of Tropical Marine Bio-resources and Ecology, Guangdong Provincial Key Laboratory of Applied Marine Biology, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China
2
College of Earth and Planetary Sciences, University of Chinese Academy of Sciences, Beijing 100049, China
3
Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences, Guangzhou 510301, China
4
Tropical Marine Biological Research Station in Hainan, Chinese Academy of Sciences, Sanya 572000, China
5
Key Laboratory of Tropical Marine Biotechnology of Hainan Province, Sanya 572000, China
*
Authors to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2020, 8(12), 956; https://doi.org/10.3390/jmse8120956
Submission received: 10 October 2020
/
Revised: 17 November 2020
/
Accepted: 19 November 2020
/
Published: 24 November 2020
(This article belongs to the Section Marine Biology)
Abstract
:Information on the in situ diet of juvenile chaetognaths is critical for understanding the population recruitment of chaetognaths and their functional roles in marine food web. In this study, a molecular method based on PCR amplification targeted on 18S rDNA was applied to investigate the diet composition of juvenile Flaccisagitta enflata collected in summer and autumn in Sanya Bay, China. Diverse diet species were detected in the gut contents of juvenile F. enflata, including copepods, small jellyfish, anthozoa, polychaetes, echinoderms, diatoms and dinoflagellates. The diet composition showed obvious differences between summer and autumn. Copepod, such as Temora turbinata, Canthocalanus pauper and Subeucalanus crassus, dominated the diet in summer, representing up to 61% of the total prey items. However, small jellyfish, mainly consisting of Bougainvillia fulva, Solmissus marshalli and Pleurobrachia globosa, was the main food group (72.9%) in autumn. Environmental parameters showed no significant difference between summer and autumn. The mean abundance of juvenile chaetognaths in autumn was about eight times higher than that in summer, while the abundance of potential food prey was similar in both seasons. Our results suggested that juveniles chaetognaths might consume small jellyfish as a supplementary food source under enhanced feeding competition in autumn.
1. Introduction
Chaetognaths are among the most abundant macro-zooplankton in coastal ecosystems worldwide. They contribute 5% to 15% of the total zooplankton biomass [1]. As main predator of mesozooplankton, such as copepods and cladocerans, chaetognaths play a central role in the planktonic food web [2,3]. They exert considerable influence on the population dynamics of their prey groups by predation [4,5]. Besides which, chaetognaths are also a food source for fish [6,7,8]. Except for the link role between small zooplankton and top predators, chaetognaths also play an essential role in the biogeochemical cycling by making a substantial contribution to vertical carbon flux through producing large, fast-sinking fecal pellets [9,10]. Simultaneously, chaetognaths can also be competitors of fish larvae because they feed on similar preys [11,12]. Thus, it is important to study population dynamics of chaetognaths and analyze the related influencing factors.
Chaetognaths are usually present year-round and reach their highest densities in some seasons. For example, in the Ionian Sea and the Cretan Passage, chaetognaths reach the highest densities in autumn after the dominant species F. enflata breeds in late summer [2,13]. The highest abundance could reach 310 ind m−3 in October in the Gullmar fjord [14]. Among the most abundant period, juvenile individuals (mostly stage I) contribute to almost 64.9%–97.7% of total chaetognath abundance [15]. Several studies showed that the high abundance of juvenile chaetognath usually coincides with the occurrence of large numbers of their food (mostly copepods) [16,17,18,19,20]. As food was believed to be one of the most critical parameters impacting the dominance of chaetognath juveniles, it is essential to study their feeding strategy to better understand the mechanisms sustaining a thriving chaetognath population in some seasons.
Small copepods/copepod nauplii were considered to be the main prey for juveniles chaetognath [2,11,17,21,22]. Also, non-copepod prey such as tintinnids and rotifers were reported to be important in the diets of juvenile chaetognaths in the South Atlantic Bight when they reach a high abundance to obtain sufficient energy [11,23,24]. Moreover, several studies suggest that chaetognaths can feed on detritus (or marine snow) when high population abundance occurred [25,26,27,28]. Therefore, exact dietary analysis of chaetognaths, especially juveniles, is essential to understand their food source sustaining such high abundance. However, methodological limitations result in biased inferences on the composition of juvenile chaetognaths diets, because of low fractions of foods [2,29], and large amounts of “unidentified food items” (their sizes (mostly < 6 mm) in their guts) [30,31,32]. Besides this, the unidentified food in diet of chaetognath juveniles (stage I of F. enflata and Parasagitta setosa) could reach as high as 35.1% [31,33]. Molecular methods based on DNA markers provide a powerful tool to resolve this problem by retrieving DNA fragments and accurately identifying their origin from partially digested or broken pieces of food items [34]. Further, this method is particularly suitable for diet identification of small-sized zooplankton, such as copepods and larval fish, which are difficult to process for gut dissection [35,36]. Bonnet et al. (2010) successfully detected copepod Calanus helgolandicus in the gut content of chaetognath Sagitta setosa using a Calanus specific primers [37], indicating that a molecular method can be effective when unraveling the diets of chaetognaths.
Sanya Bay is a typical tropical bay in the north of South China Sea, characterized by abundant marine resources and high biodiversity [38]. Chaetognaths are distributed widely in Sanya Bay, and the highest abundance usually occurs in summer and autumn. In autumn, chaetognaths could reach an abundance of 79.68 ind m−3, with F. enflata being the most dominant species accounting for as high as 90% of the total chaetognath abundance, especially juvenile individuals [39,40]. Meanwhile, the dominant food for chaetognaths, especially copepods, was relatively low during autumn [40]. As food was believed to be one of the most critical parameters impacting the survival of chaetognath juveniles, we hypothesized that there might be other food sources other than already known preys to sustain a high abundance of juvenile-dominated chaetognath community in this tropical bay. Therefore, we analyze the diet composition of juvenile F. enflata collected from summer and autumn in Sanya Bay by molecular method, with the purpose of revealing the potential resource supporting the high abundance of chaetognaths based on the precise food detection of the molecular method.
2. Materials and Methods
2.1. Sample Collection
Sampling was conducted at the intermediate zone (W3 station) and off-shore areas (W4 station; W9 station) of Sanya Bay in July 29 and October 26 of 2014 (Figure 1) [41]. The water depth was ~19 m in W3, 25 m in W4 and 28 m in W9.
Physical and chemical parameters in the environment (temperature, salinity, dissolved oxygen concentration, pH and dissolved organic carbon concentration) were measured using the YSI6600 Water Quality System.
Chaetognath juveniles were collected using a cylindro-conical net (50 cm aperture, 145 cm height, and 505 μm mesh size) towed vertically from the bottom to the surface without replicate. The towing speed was 1 m sec−1. To prevent any possible changes in chaetognath juvenile gut content as an artifact of sampling, collected samples were fixed after transfer to the bottle by adding neutral Lugol’s solution at 2% final concentration. All sampling and preservation processes were completed within two minutes. Neutral Lugol’s (no acetic acid added) had been shown to effectively preserve samples for DNA analysis in zooplankton [42,43]. Other sets of zooplankton samples were collected and preserved in a 5% formalin-seawater solution for species identification using stereomicroscope. The sample IDs were as follows: W3-Jul-J, W4-Jul-J, W9-Jul-J, W3-Oct-J, W4-Oct-J and W9-Oct-J (Wx means the sample station, Jul means sample was collected in July, Oct means sample was collected in October, J means F. enflata juveniles).
2.2. Zooplankton Identification and Statistical Analysis
To obtain the information of potential food organisms, zooplankton samples were split using a Folsom splitter as they were abundant, and then were identified and counted under the stereomicroscope [2]. Zooplankton abundance was expressed as individuals per cubic meter (ind m−3). One-way ANOVA analysis in SPSS22.0 data analysis software was used to test for significant difference of environmental parameters.
2.3. DNA Extraction of F. enflata Juveniles
F. enflata juveniles (in stage I which had no visible ova) were identified and sorted using a wide-bore plastic pipette under stereomicroscope Leica S8APO in the laboratory [44,45,46]. The length of juvenile specimens used in this study were in the range of 3–6 mm. To avoid artifacts of prey items from cod-end feeding, 1/3 of the forward gut of all chaetognaths under examination was cut and thrown away. Then, the sorted chaetognaths were thoroughly rinsed three times with autoclaved 0.45 μm-filtered seawater and examined under the stereomicroscope to ensure that no other visible organisms were attached on the body surface. Then, F. enflata juveniles (100 individuals for each station) were homogenized in a microfuge tube using a disposable micro-pestle, re-suspended in 500 μL DNA extraction buffer (1% SDS, 100 Mm EDTA and 200 μg mL−1 proteinase K) and incubated for 1 day at 55 °C for complete cell lysis. DNA from all samples was then extracted following a CTAB protocol [47]. The specific operation steps were as follows. DNA was isolated by adding 16.5 µL each of 5 M NaCl and 10% cetyltrimethylammonium bromide (Sigma) in 0.7 M NaCl and incubating at 55 °C for 10 min, followed by one chloroform extraction and one phenol-chloroform extraction. DNA was then purified by being passed twice through DNA Clean and Concentrator columns (Zymo Re-search, Orange, CA). DNA was dissolved in 30 µL of distilled and deionized water and stored at −20 °C until PCR was performed.
2.4. Primers Design, Verification and PCR Protocol
A new sequence alignment of the 18S rDNA of F. enflata was generated using CLUSTAL W (1.8) including homologs from other chaetognath species and representatives of other marine eukaryotes deposited in the GenBank database (>100 lineages). Based on this alignment, new primers were designed on regions that were conserved in eukaryotes but unique in F. enflata and other chaetognaths. The primers were aimed to detect as many lineages of marine eukaryotes as possible but to exclude F. enflata and other chaetognaths. Non-chaetognath primer set (Forward primer: 5′-GAGCTAATACATGCNAARAVDCTC-3′ and Reverse primer: 5′-GCAAATGCTTTCGCWGTAGTYHGT-3′) was then selected. Before using it to amplify target genes extracted from the guts of F. enflata juveniles, 15 eukaryotic species from different groups, including dinoflagellate, cnidaria, copepoda, decapoda, echinodermata and fish were used to verify the accuracy of the primer set, which all showed successful amplification. PCR protocol used was as follows: an initial denaturing step at 95 °C for 30 s, 35 cycles of denaturation at 95 °C for 1 min and 20 s, annealing at 55 °C for 30 s, and extension at 72 °C for 40 s. The PCR products were concentrated using Zymo DNA Clean & Concentrator TM-25 Kit, then the target bands (~0.78 kb) were recovered from 1% agarose gel using Zymoclean TM Gel DNA Recovery Kit (ZYMO RESEARCH). The purified PCR products were then ligated into DH-5α component cells (TaKaRa), and 60 clones were randomly selected for sequencing (Invitrogen sequencing company).
2.5. Bioinformatic Analysis
Obtained sequences were searched against the GenBank database using the Basic Local Alignment Search Tool (BLASTn, https://www.ncbi.nlm.nih.gov/) after the primer sequences were trimmed. The resulting alignment was imported into MEGA 6.0 to identify the best-fit nucleotide substitution model to infer phylogeny [48]. The best-fit model Kimura 2 with gamma distribution (K2 + G) was then employed for Maximum Likelihood (ML) analysis. The reliability of the tree topology was evaluated using bootstrap analysis with 1000 replicates, and the tree is rooted with Rat sp. [35]. Diversity indices (Shannon-Wiener, Simpson) and chao1 (This index was predicted theoretical richness from sequence results. If the number of detected taxa was closer to it, the detected taxa could reflect the actual value more.) were estimated using Past 3.05 (http://folk.uio.no/ohammer/past/). The percentage of every category in each sample was calculated using the detected clones divided by the total clone libraries.
3. Results
3.1. Environmental Parameters
As a tropical bay, there were no significant seasonal variations in environmental factors (Table 1). The average water temperature in surface layer was 27.78 ± 0.10 °C and 28.15 ± 0.09 °C in summer and autumn, respectively, while the bottom layer temperature was 25.31 ± 0.74 °C in summer, indicating a stratification in summer. The salinity was 34.95 ± 0.05 and 33.32 ± 0.18 in summer and autumn, respectively, and it was evenly distributed with depth, with no significant variations in both seasons (p > 0.05). Dissolved oxygen concentrations and pH were in the range of 6.25–6.62 mg L−1 and 8.13–8.16 in summer and autumn, respectively, with no significant variations (p > 0.05). However, the mean dissolved organic carbon concentration was slightly higher in summer (3.84 mg L−1) than in autumn (1.80 mg L−1), but also with no significant variations in both seasons (p > 0.05). The mean concentration of Chla was 2.33 ug L−1 and 19.95 ug L−1 in summer and autumn, respectively, with no significant variations (p > 0.05).
3.2. Zooplankton Community
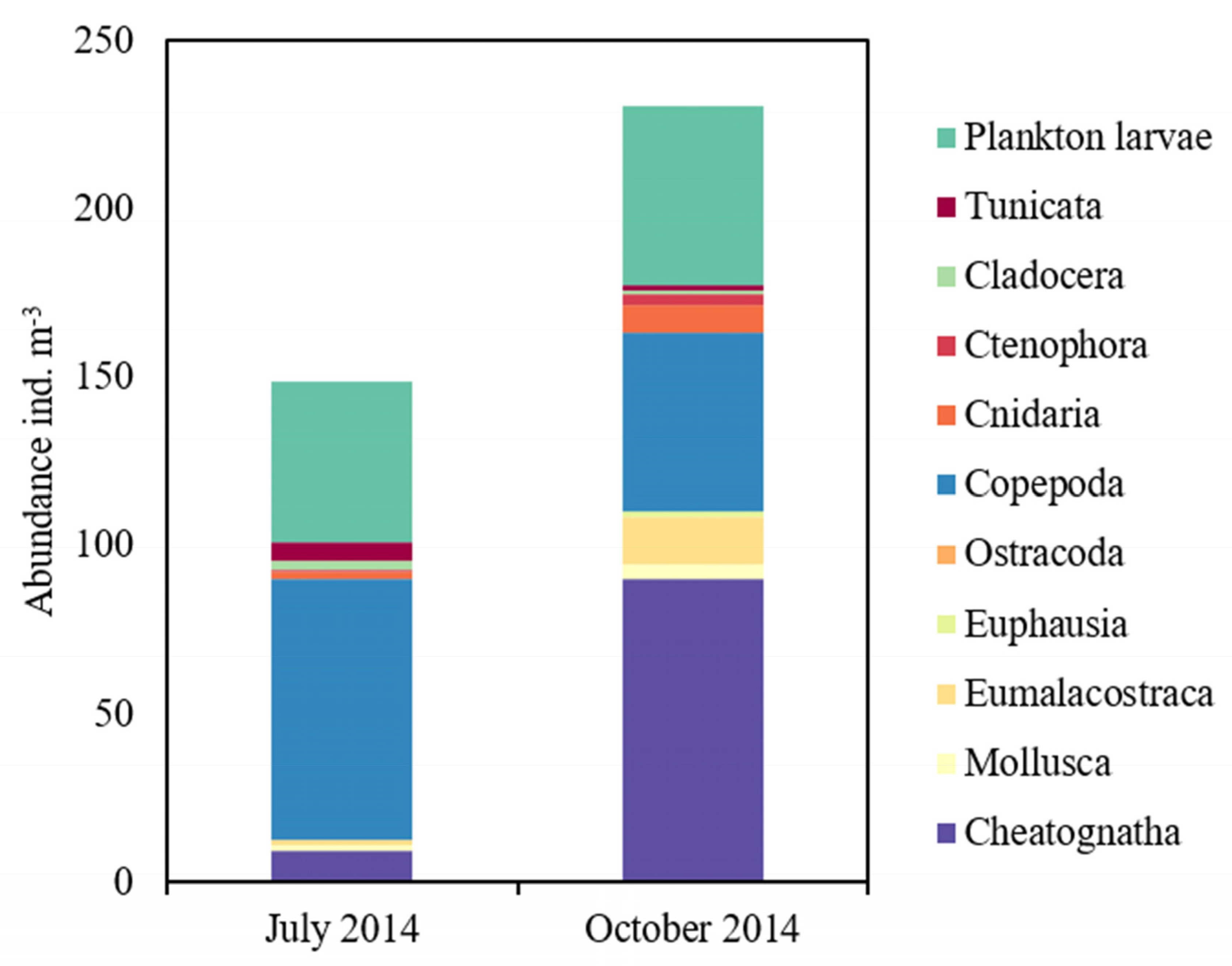
Microscopic identification of zooplankton samples found a total of 57 species belonging to copepoda, chaetognatha, eumalacostraca, ostracoda, euphausiacea, cladocera, mollusca, cnidaria, ctenophora and tunicata, as well as planktonic larvae (Figure 2). At total of 45 and 33 species were found in summer and autumn, respectively. The abundance of zooplankton ranging from 148.64 ± 50.43 ind m−3 and 230.86 ± 147.9 ind m−3(p > 0.05) in summer and autumn, respectively (Appendix A). Copepoda was one of the domianant group, constituting 51.98% and 23.01% of zooplankton abundances in summer and autumn, respectively. Among the copepods, Temora turbinata, Acartia eryhraea, Centropages orsinii, Tortanus gracilis, Acrocalanus gibber, Canthocalanus pauper and Subeucalanus subcrassus were the dominant species in summer. Subeucalanus subcrassus, Acartia eryhraea, Subeucalanus crassus, Tortanus gracilis and Canthocalanus pauper were the dominant species in autumn. Planktonic larvae were another abundant group, constituted 28.71% of zooplankton abundance, mainly including those associated with Ophiopluteus, Lucifer, Polychaeta, Macruran and Brachyura larvae.
Chaetognatha also made a large contribution to zooplankton group. Total of four species were found, including Flaccisagitta enflata, Zonosagitta bedoti, Aidanosagitta delicata and A. johorensis. They all presented in summer, but only F. enflata and A. delicata presented in autumn. The abundance of chaetogantha constituted 6.21% and 39.07% of zooplankton abundance in summer and in autumn, respectively (Figure 2). The abundance of chaetognaths was 9.24 ± 6.12 ind m−3 in summer and increased to 90.2 ± 56.17 ind m−3 in autumn. F. enlfata was the most abundant species consisting 62.77% (summer) and 93.07% (autumn) of the total chaetognaths. In addition, the mean abundance of juvenile chaetognaths in autumn (159.09 ind m−3) was about eight times higher than that in summer (17.35 ind m−3) (Appendix A).
There were also eight species of small jellyfish (Cnidaria and Ctenophora) found. The abundance of small jellyfish was higher in autumn (11.23 ± 7.63 ind m−3) than in summer (2.9 ± 2.42 ind m−3) (Appendix A). Their contribution was always <5% of the zooplankton stock, mostly coming from Lensia subtiloides, Diphyes chamissonis, Nanomia bijuga, and Pleurobrachia globosa.
3.3. Diet Composition of F. enflata Juveniles in Different Seasons
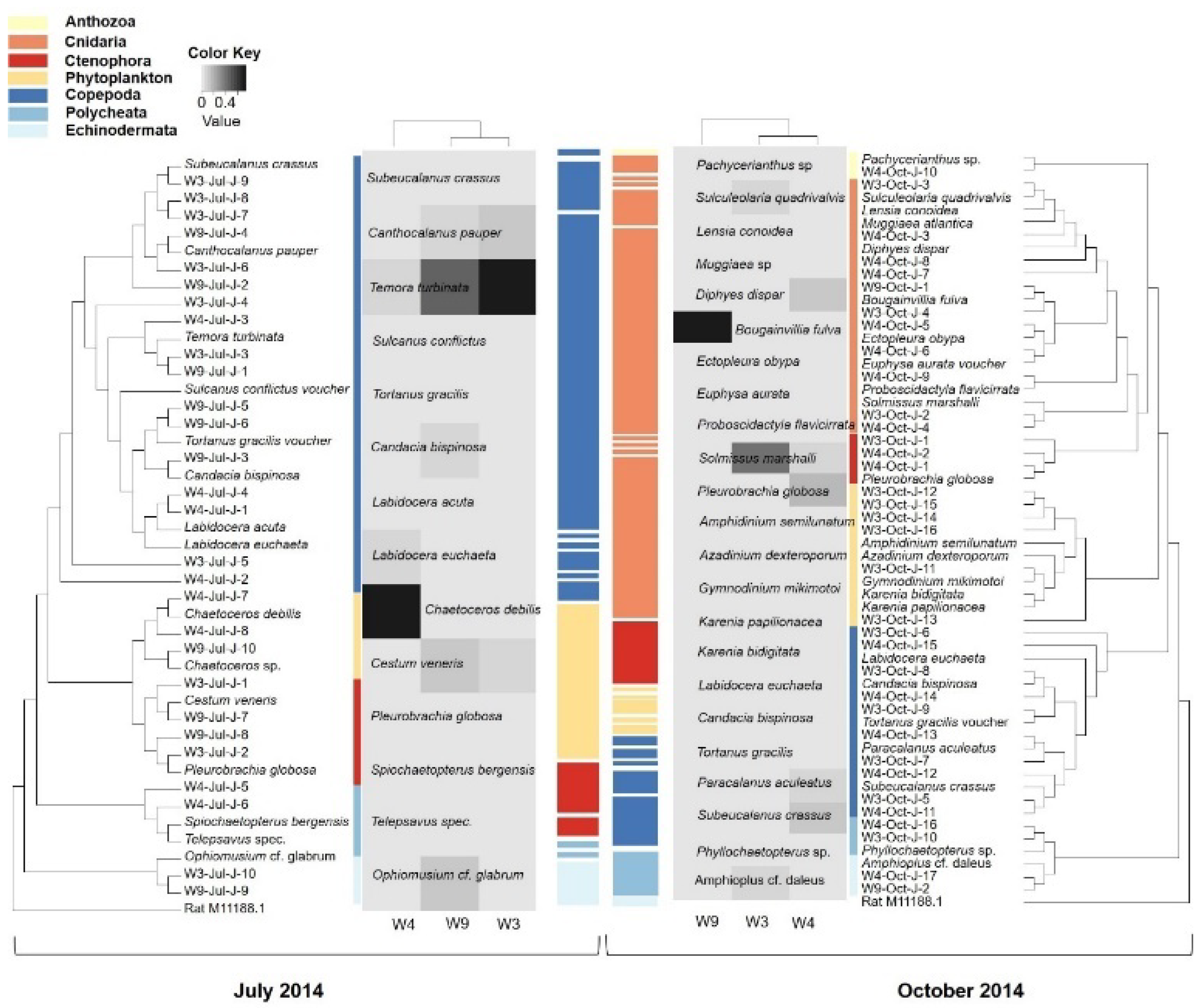
In total, 34 taxa belonging to eight groups, Ctenophora, Cnidaria, Anthozoa, Copepoda, Polychaeta, Echinodermata, Bacillariophyta and Dinophyceae, were detected from all the samples (Appendix B). Small jellyfish (Cnidaria and Ctenophora) was the most abundant prey group, accounting for 0%–62.75% (percentage of clones) of the total clone libraries in different samples. Bougainvillia fulva, Solmissus marshalli and Pleurobrachia globosa were the dominant species. The proportions of copepods were also significant (10.34%–88.89%), with Temora turbinate, S. crassus and Canthocalanus pauper dominating the community. Polychaeta, Echinodermata and Anthozoa made few contributions (>8.18%), and were occasionally found. Also notable in the clone libraries were several phytoplankton taxa (12.27%), including the dinoflagellates (Gymnodinium mikimotoi, Karenia bidigitata) and diatom (Chaetoceros debilis).
F. enflata juveniles showed little difference in their food construction between seasons (Table 2), as concerning the average number of taxa (9.3 and 11.7 in summer and autumn, respectively; p > 0.05), and Shannon diversity (1.4983 and 1.4276 in summer and autumn, respectively; p > 0.05). However, copepods were the most abundant preys in summer, accounting for 60.51% of the total diet, while small jellyfish dominated the diets of F. enflata juveniles in autumn with a relative percentage contribution of 72.90% (Figure 2).
4. Discussion
It is vital to explore the spectrum of food choice and preference of chaetognaths in the natural environment, because chaetognaths play a significant role in structuring the zooplankton community by exerting top-down control over other groups through predation [9]. Juveniles are considerably significant components of the chaetognath community and also serve as an indispensable group for recruitment. However, the diet of juvenile chaetognaths is difficult to identify due to high proportions of unidentifiable particles, sometimes making up to 100% of the gut content, due to rapid digestion of the ingested prey [31,32]. In the present study, a predator-specific primer set was used to investigate the in situ diet of juvenile chaetognaths using a molecular approach. A more diverse diet composition was accurately revealed by this primer set compared to using morphological identification alone [31]. Though cannibalism could not be detected with our method, other marine organisms known to have trophic interactions with chaetognaths were also found. In addition to the commonly reported food for chaetognaths, such as copepods, polychaeta, other species belonging to diatoms, dinoflagellates, echinoderms and anthozoans were also detected, albeit in lower abundance based on their proportion in the clone library. Strikingly, jellyfishes dominated the prey items of juvenile F. enlfata in autumn based on reconstructed food assemblage with high resolution and a semi-quantitative molecular method [34], indicating that jellyfishes might be important supplementary food sources for the chaetognath community in striving seasons.
4.1. Prey Diversity
In this study, the most abundant prey were diverse metazoan species, such as copepods, cnidaria, ctenophora and polychaeta. Most of the prey groups in this study had also been detected by microscopic methods (Appendix C), but were only mostly identified down to the genus or class levels in previous studies [2,23,49,50]. Prey belonging to echinodermata and anthozoa, which were not reported before, were also detected in this study, allowing identification of other food items not easily detected using conventional techniques.
F. enflata is thought to be an ambush-type carnivore based on its feeding behavior, mainly preying on copepods and cladocerans due to their high abundance in the ambient water [2,51,52,53]. It is not surprising then that copepods constitute a significant proportion of the diet of juvenile chaetognaths in this study (Figure 3). Classic gut content examination under the microscope is an adequate way to determine which copepod stages are preferred by chaetognath juveniles. However, the method we applied cannot discriminate between the copepod’s ontogenetic stages such as adults, copepodites and nauplii. Considering that the prey ingested by F. enflata followed a head-width-body-length ratio of 0.0758 [54], it is reasonable to assume that small calanoid copepod nauplii had an ideal size for predation by juvenile chaetognaths [17]. In addition, this had been verified both in the field and laboratory [22,55]. Although we cannot determine the exact abundance of copepod nauplii from each copepod species, this ontogenetic stage was the most abundant in the water column during our sampling time (Appendix A) [40]. The mean abundance of copepod nauplii in the surrounding waters reached 2705.84 ind m−3 and 5382.05 ind m−3 in summer and autumn, respectively (collected by 160 μm mesh size cylindro-conical net). A high abundance of copepod nauplii also increased chance encounters with chaetognath juveniles in the shallow waters.
Other prey items found in the present investigation, such as polychaetes, were also detected in chaetognath guts in previous studies [49,51]. Polychaetes and echinoderms were also common inhabitants of the macrobenthos in Sanya Bay [40,56,57]. Considering the size relation of these preys and F. enflata [54], the species detected here may have originated from their planktonic larvae [58,59,60,61,62]. Furthermore, phytoplankton also contributed significantly to clone libraries (12.27%) mostly belonging to dinoflagellates (G. mikimotoi and K. bidigitata) and diatoms (Chaetoceros sp.). Because of the small body size and their smooth ciliary motion, phytoplankton were not thought to be directly ingested by chaetognaths but were rather derived from other herbivores that were then consumed by chaetognaths [1]. However, chaetognaths also exhibit omnivory and/or detritivory based on fatty acid markers detecting green-detritus in the gut [25], indicating that phytoplankton may be accidentally ingested when juvenile chaetognath ingests/gulps waters.
4.2. Small Jellyfish as Supplementary Food Sources for Juvenile Chaetognaths in Autumn
In our results, small jellyfish was the main constituent in the diets of F. enflata juveniles with a relative percentage contribution of 72.90% in autumn. This is not due to technical errors, such as primer preference or PCR bias due to differences in 18S copy, because the genome size of copepods is usually bigger than small jellyfish [63,64]. Although the abundance of jellyfish was low in our sampled zooplankton samples due to a relatively large sampling mesh, the detected species D. dispar and P. globosa were also present in the zooplankton samples and were common in Sanya Bay in autumn [40,57]. Previous studies also found jellyfish remains in chaetognath guts, which were first thought to be artifacts of collecting and preserving processes, since jellyfish tended to be too large to pass through the mouth of chaetognaths [1]. However, Kruse el al. (2010) found that in deep dwelling Eukrohnia hamata, jellyfish remains (nematocyst) can be an important fraction of the preys [65]. Besides which, Giesecke and Gonzalez (2012) found that maximum abundances of siphonophores match with the highest proportions of jellyfish remains in E. hamata guts in the Lazarev sea [49]. These studies suggest that jellyfish may be important energy sources for chaetognaths and so do the juvenile chaetognaths in our study area.
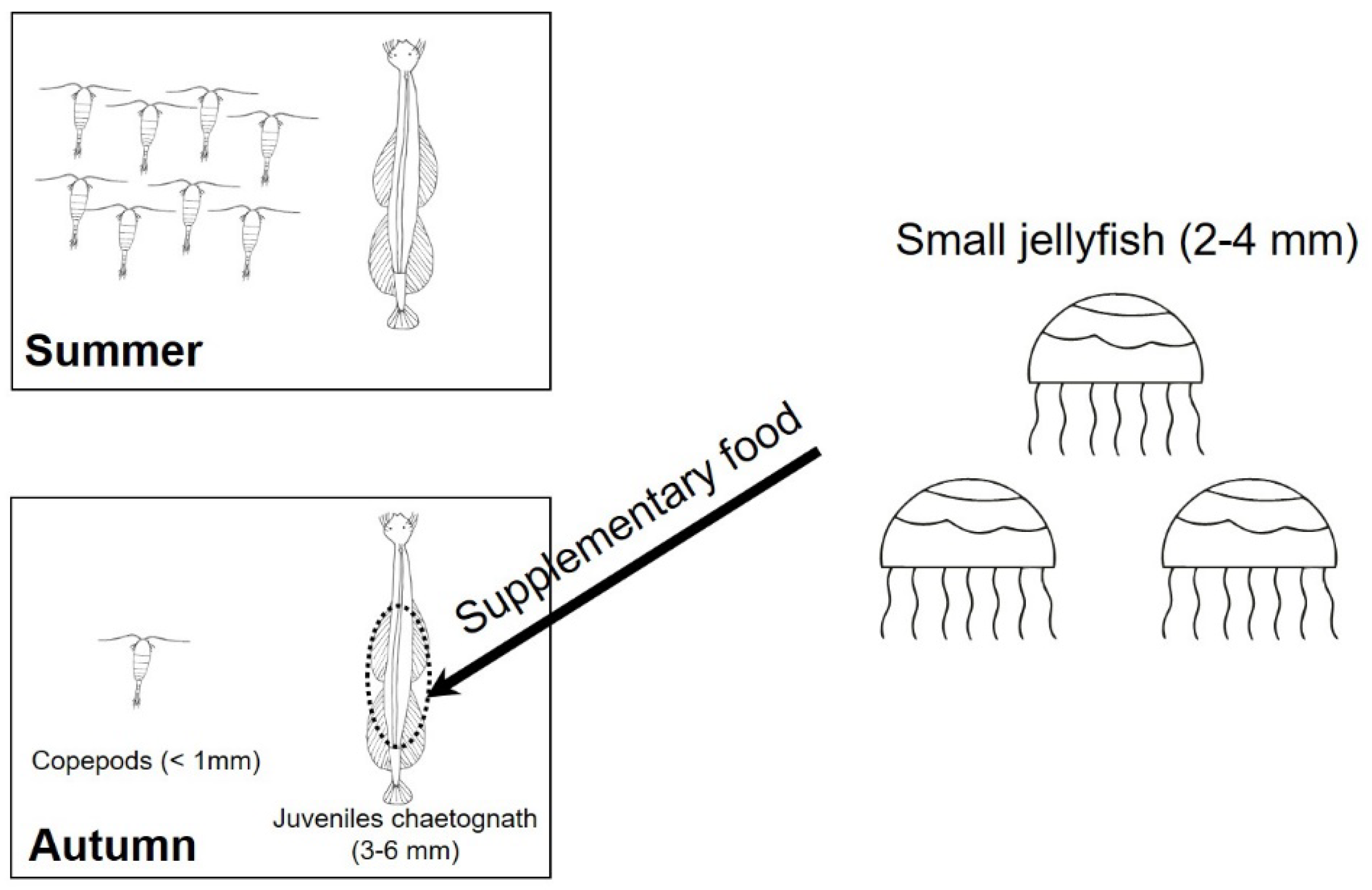
In this study, the abundance of chaetognaths increased eight times in autumn than in summer. Among which juveniles chaetognath occupied a dominant position. Meanwhile the copepod nauplii, which were considered to be the preferred prey for juvenile chaetognaths, were not increased proportionally. Conversely, the ratio of copepod nauplii to F. enflata juveniles (in forms of abundance) decreased significantly in autumn (28) compared with that in summer (228) (p < 0.05), indicating a relatively short supply of preferred prey for juvenile chaetognaths. They might consume other easily available preys in such a fierce competition food environment, likely small jellyfish. Assuming the removal of the influence of water flow, copepod nauplii showed much higher escape speeds (>500 mm s−1) from predators than small jellyfish (4.8–9 mm s−1) [44,66,67]. Thus, small jellyfish may be more easily captured than copepod nauplii by the ambushing chaetognath juveniles and the jellyfish detected here might be consumed by juvenile chaetognath directly, considering their size difference in the forms of planula, polyps, medusa (Figure 4). Although jellyfish bodies were composed of almost 95% water, they were still suitable bait for the culture of phyllosomas of Ibcus novemdentatus (Decapoda: Scyllaridae) and Pampus argenteus juveniles for their high amino acid content [68,69,70]. Other species such as thread sail filefish Stephanolepis cirrhifer and silver pomfret P. argenteus also consumed large amounts of jellyfish as food [71,72]. Previous studies also found that chaeognaths can directly feed on detritus or take in particle organic matters when gulping water [26,27]. Moreover, more unidentified food (usually classed as detritus) in juvenile chaetognath gut was observed when they reach a high abundance [33], so the small jellyfish detected in the gut here might have also originated from detritus containing body remains of jellyfish, considering the high abundance of juveniles chaetognath in our sampling station [40]. Thus, small jellyfish may be important supplementary energy sources for chaetognaths. This might be an important feeding strategy for juvenile chaetognaths to reduce competition and maintain population stability in a competitive food environment like Sanya Bay.
The abundance of jellyfish has increased in a number of regions throughout world in recent decades [73], and they were long considered as ‘dead ends’ within marine food webs [74]. They can bloom and cause numerous deleterious consequences for industry and the community, such as considerable damage to the fishery production from the competition for food with fish [75]. The notion of jellyfish as trophic dead ends has become largely obsolete in recent years, because more and more studies have found opportunistic carnivores that feed upon jellyfish [76]. In our study, small jellyfish can be consumed by other mesozooplankton, such as chaetognaths, providing a pathway by which jellyfish can participate in the pelagic food web. Chaetognaths may potentially control the population size of jellyfish owing to their large high abundance. Therefore, more feeding experiments about jellyfish and chaetognaths are needed to gain comprehensive knowledge of the trophic interactions among zooplankton community.
5. Conclusions
This study provides a snapshot of the population dynamic and distribution pattern of chaetognath from the trophic aspects. Furthermore, our results suggest that there may be more complex trophic interactions in the tropical areas. The high taxonomic resolution of potential prey and uncommon food species by molecular method documented here expanded our understanding of chaetognath dietary range. Juvenile chaetognaths consumed large amounts of jellyfish when faced with a competitive food environment. This flexible feeding strategy could reduce inter- or intra-specific competition, helping them survive and maintaining high abundance in autumn. This might be one of the reasons for their success in distributing themselves in almost any environment. Furthermore, autumn was the breeding time, and the supplementary food sources provide a guarantee for their metabolic demands and fast growth of juveniles, which was important for the population to thrive. Moreover, our results provide a possible pathway by which jellyfish transfer materials and energy to the higher trophic levels. Therefore, more attention should be paid to evaluate the role of chaetognath in the link to high trophic levels, not just as a predator in the marine food webs.
Author Contributions
Conceptualization, S.L., S.H. and L.W.; methodology, M.G.; software, L.W.; validation, L.W., S.L. and T.L.; formal analysis, L.W.; investigation, L.W.; resources, S.L. and S.H.; data curation, L.W., T.L.; writing—original draft preparation, S.H. and L.W.; writing—review and editing, L.W., S.L., H.H. and T.L.; supervision, S.L. and S.H.; project administration, S.L. and H.H.; funding acquisition, S.L. and H.H.. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (No. XDA13020100), Natural Science Foundation of China (No. 41806188), National Key Research and Development Project of China (No. 2016YFC0502800), Institution of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences (No. ISEE2018PY01), and Science and Technology Planning Project of Guangdong Province, China (No. 2017B0303014052).
Acknowledgments
We thank Youjun Wang of Chinese Academy of Sciences for his assistance in collecting the samples.
Conflicts of Interest
The authors declare no conflict of interest.
Data Availability
The partial 18S rDNA gene sequences generated in this study are available in GenBank under accession numbers MK033893-MK033955.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Mean (±SD) abundance (ind m−3), relative abundance (RA), frequency of occurrence (FO) and abundance range of all zooplankton species found in Sanya Bay.
Table A1.
Mean (±SD) abundance (ind m−3), relative abundance (RA), frequency of occurrence (FO) and abundance range of all zooplankton species found in Sanya Bay.
| July 2014 | October 2014 | |||||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SD | RA | FO | Range | Mean ± SD | RA | FO | Range | |
| Total zooplankton | 148.64 ± 50.43 | 100.00% | 100.00% | 115.42−206.67 | 230.86 ± 147.9 | 100.00% | 100.00% | 96.88−389.57 |
| Cheatognatha | 9.24 ± 6.12 | 6.21% | 100.00% | 2.5−14.44 | 90.2 ± 56.17 | 39.07% | 100.00% | 33.75−146.09 |
| Flacciagitta enflata | 5.8 ± 4.83 | 3.90% | 100.00% | 1.67−11.11 | 83.95 ± 51.27 | 36.36% | 100.00% | 33.13−135.65 |
| Zonosagitta bedoti | 0.74 ± 1.28 | 0.50% | 33.33% | 0−2.22 | ||||
| Aidanosagitta delicala | 2.19 ± 2.11 | 1.47% | 100.00% | 0.83−4.62 | 6.25 ± 5.06 | 2.71% | 100.00% | 0.63−10.43 |
| Aidanosagitta johorensis | 0.51 ± 0.89 | 0.35% | 33.33% | 0−1.54 | ||||
| Mollusca | 1.65 ± 1.46 | 1.11% | 100.00% | 0.77−3.33 | 4.09 ± 5.79 | 1.77% | 100.00% | 0.63−10.77 |
| Creseis acicula | 1.75 ± 2.5 | 0.76% | 66.67% | 0−4.62 | ||||
| Creseis clava | 1.65 ± 1.46 | 1.11% | 100.00% | 0.77−3.33 | 0.8 ± 0.77 | 0.35% | 66.67% | 0−1.54 |
| Creseis virgula | 0.51 ± 0.89 | 0.22% | 33.33% | 0−1.54 | ||||
| Trochidae | 1.03 ± 1.78 | 0.45% | 33.33% | 0−3.08 | ||||
| Eumalacostraca | 1.42 ± 0.78 | 0.95% | 100.00% | 0.83−2.31 | 14.37 ± 18.44 | 6.22% | 100.00% | 3.08−35.65 |
| Lestrigonus macrophthalmus | 0.53 ± 0.46 | 0.36% | 66.67% | 0−0.83 | ||||
| Lucifer intermedius | 0.88 ± 0.79 | 0.59% | 66.67% | 0−1.54 | 13.33 ± 19.36 | 5.77% | 100.00% | 1.25−35.65 |
| Lucifer hanseni | 1.04 ± 1.8 | 0.45% | 33.33% | 0−3.13 | ||||
| Ostracoda | 0.37 ± 0.64 | 0.25% | 33.33% | 0−1.11 | ||||
| Euconchoecia aculeata | 0.37 ± 0.64 | 0.25% | 33.33% | 0−1.11 | ||||
| Copepoda | 77.27 ± 16.5 | 51.98% | 100.00% | 61.54−94.44 | 53.11 ± 36.21 | 23.01% | 100.00% | 18.13−90.43 |
| Canthocalanus pauper | 7 ± 2.18 | 4.71% | 100.00% | 4.62−8.89 | 7.12 ± 6.23 | 3.08% | 100.00% | 0.63−13.04 |
| Undinula valgaris | 1.48 ± 2.57 | 1.00% | 33.33% | 0−4.44 | ||||
| Subeucalanus subcrassus | 2.83 ± 1.48 | 1.90% | 100.00% | 1.54−4.44 | 13.67 ± 10.22 | 5.92% | 100.00% | 1.88−20 |
| Temora turbinata | 22.31 ± 4.95 | 15.01% | 100.00% | 16.92−26.67 | 1.45 ± 2.51 | 0.63% | 33.33% | 0−4.35 |
| Paracalanus parvus | 0.37 ± 0.64 | 0.25% | 33.33% | 0−1.11 | 0.58 ± 1 | 0.25% | 33.33% | 0−1.74 |
| Acrocalanus gibber | 1.03 ± 1.78 | 0.69% | 33.33% | 0−3.08 | 0.51 ± 0.89 | 0.22% | 33.33% | 0−1.54 |
| Acrocalanus gracilis | 0.77 ± 1.33 | 0.52% | 33.33% | 0−2.31 | ||||
| Centropages orsinii | 10.83 ± 6.98 | 7.28% | 100.00% | 6.67−18.89 | ||||
| Centropages forcatus | 0.26 ± 0.44 | 0.17% | 33.33% | 0−0.77 | 3.34 ± 3.49 | 1.45% | 66.67% | 0−6.96 |
| Candacia truncata | 2.92 ± 2.28 | 1.96% | 100.00% | 1.54−5.56 | ||||
| Labidocera euchaeta | 0.51 ± 0.89 | 0.35% | 33.33% | 0−1.54 | ||||
| Corycaeus speciosus | 0.26 ± 0.44 | 0.17% | 33.33% | 0−0.77 | ||||
| Corycaeus affinis | 0.53 ± 0.46 | 0.36% | 66.67% | 0−0.83 | ||||
| Tortanus gracilis | 9.49 ± 3.45 | 6.38% | 100.00% | 6.67−13.33 | 3.78 ± 2.83 | 1.64% | 100.00% | 0.63−6.09 |
| Oncaea venusta | 0.26 ± 0.44 | 0.17% | 33.33% | 0−0.77 | ||||
| Copilia mirabilis | 0.63 ± 0.57 | 0.42% | 66.67% | 0−1.11 | ||||
| Temora stylifera | 0.26 ± 0.44 | 0.17% | 33.33% | 0−0.77 | 0.29 ± 0.5 | 0.13% | 33.33% | 0−0.87 |
| Corycaeus andrewsi | 1.11 ± 1.92 | 0.75% | 33.33% | 0−3.33 | ||||
| candacia bradyi | 0.77 ± 1.33 | 0.52% | 33.33% | 0−2.31 | 0.8 ± 0.77 | 0.35% | 66.67% | 0−1.54 |
| calanus sinicas | 0.51 ± 0.89 | 0.35% | 33.33% | 0−1.54 | ||||
| Oncaea ornata | 0.28 ± 0.48 | 0.19% | 33.33% | 0−0.83 | ||||
| Acartia eryhraea | 12.88 ± 11.22 | 8.67% | 100.00% | 6.15−25.83 | 1.88 ± 3.25 | 0.81% | 33.33% | 0−5.63 |
| Temora discaudata | 0.51 ± 0.89 | 0.22% | 33.33% | 0−1.54 | ||||
| Labidocera sp. | 1.16 ± 2.01 | 0.50% | 33.33% | 0−3.48 | ||||
| Pontellopsis inflatodigitata | 0.21 ± 0.36 | 0.09% | 33.33% | 0−0.63 | ||||
| Subeucalanus pileatus | 0 ± 0 | 0.00% | 0.00% | 0−0 | ||||
| Subeucalanus crassus | 17.81 ± 13.98 | 7.71% | 100.00% | 8.75−33.91 | ||||
| Euphausiacea | 1.74 ± 3.01 | 0.75% | 33.33% | 0−5.22 | ||||
| Pseudeuphausia sinica | 1.74 ± 3.01 | 0.75% | 33.33% | 0−5.22 | ||||
| Cnidaria | 2.16 ± 1.03 | 1.45% | 100.00% | 0.83−3.33 | 7.95 ± 4.05 | 3.45% | 100.00% | 3.13−13.04 |
| Lensia subtiloides | 0.37 ± 0.64 | 0.25% | 33.33% | 0−1.11 | 3.81 ± 1.22 | 1.65% | 100.00% | 3.08−5.22 |
| Diphyes chamissonis | 0.65 ± 0.58 | 0.44% | 66.67% | 0−1.11 | 2.03 ± 3.51 | 0.88% | 33.33% | 0−6.09 |
| Aglaura hemistoma | 0.26 ± 0.44 | 0.17% | 33.33% | 0−0.77 | 0.29 ± 0.5 | 0.13% | 33.33% | 0−0.87 |
| Euphysora bigelowi | 0.37 ± 0.64 | 0.25% | 33.33% | 0−1.11 | 0.29 ± 0.5 | 0.13% | 33.33% | 0−0.87 |
| Halyractinia carnea | 0.26 ± 0.44 | 0.17% | 33.33% | 0−0.77 | ||||
| Nanomia bijuga | 0.26 ± 0.44 | 0.17% | 33.33% | 0−0.77 | 1.03 ± 1.78 | 0.45% | 33.33% | 0−3.08 |
| Vannucci aforbesii | 0.51 ± 0.89 | 0.22% | 33.33% | 0−1.54 | ||||
| Ctenophora | 0.74 ± 1.28 | 0.50% | 33.33% | 0−2.22 | 3.28 ± 2.85 | 1.42% | 66.67% | 0−5.22 |
| Pleurobrachia globosa | 0.74 ± 1.28 | 0.50% | 33.33% | 0−2.22 | ||||
| Ctenophores | 3.28 ± 2.85 | 1.42% | 66.67% | 0−5.22 | ||||
| Cladocera | 2.86 ± 0.87 | 1.92% | 100.00% | 2.22−3.85 | 0.87 ± 1.51 | 0.38% | 33.33% | 0−2.61 |
| Penilia avirostris | 2.58 ± 1.13 | 1.73% | 100.00% | 1.67−3.85 | 0.87 ± 1.51 | 0.38% | 33.33% | 0−2.61 |
| pseudevadne tergestina | 0.28 ± 0.48 | 0.19% | 33.33% | 0−0.83 | ||||
| Tunicata | 5.3 ± 6.63 | 3.57% | 100.00% | 0.77−12.92 | 1.67 ± 1.74 | 0.72% | 66.67% | 0−3.48 |
| Oikopleura rufescens | 2.73 ± 3.81 | 1.84% | 66.67% | 0−7.08 | ||||
| Oikopleura longicauda | 1.67 ± 2.89 | 1.12% | 33.33% | 0−5 | 1.67 ± 1.74 | 0.72% | 66.67% | 0−3.48 |
| Oikopleura dioica | 0.28 ± 0.48 | 0.19% | 33.33% | 0−0.83 | ||||
| Doliolum denticulatum | 0.26 ± 0.44 | 0.17% | 33.33% | 0−0.77 | ||||
| Doliolum gegenbauri | 0.37 ± 0.64 | 0.25% | 33.33% | 0−1.11 | ||||
| Planktonic larvae | 42.68 ± 29.32 | 28.71% | 100.00% | 16.67−74.44 | 53.58 ± 28.9 | 23.21% | 100.00% | 36.88−86.96 |
| Ophiopluteus larvae | 3.1 ± 4.66 | 2.08% | 66.67% | 0−8.46 | ||||
| Lucifer larvae | 17.59 ± 11.74 | 11.84% | 100.00% | 10−31.11 | 20.86 ± 15.13 | 9.04% | 100.00% | 7.69−37.39 |
| Polychaeta larvae | 1 ± 1.13 | 0.67% | 66.67% | 0−2.22 | 0.51 ± 0.89 | 0.22% | 33.33% | 0−1.54 |
| Macruran larvae | 13.5 ± 13.34 | 9.09% | 66.67% | 0−26.67 | 10.64 ± 9.95 | 4.61% | 100.00% | 2.5−21.74 |
| Brachyura larvae | 6.58 ± 5.85 | 4.43% | 100.00% | 3.08−13.33 | 15.21 ± 5.77 | 6.59% | 100.00% | 10.77−21.74 |
| Nauplius A | 0.53 ± 0.46 | 0.36% | 66.67% | 0−0.83 | ||||
| Nauplius B | 0.37 ± 0.64 | 0.25% | 33.33% | 0−1.11 | ||||
| Radiant larvae | 4.86 ± 3.87 | 2.11% | 100.00% | 1.88−9.23 | ||||
| Echinoplutes larvae | 0.21 ± 0.36 | 0.09% | 33.33% | 0−0.63 | ||||
| Fish larva | 1.2 ± 1.25 | 0.81% | 66.67% | 0−2.5 | 0.58 ± 1 | 0.25% | 33.33% | 0−1.74 |
| Fish egg | 3.76 ± 3.41 | 2.53% | 66.67% | 0−6.67 | 0.71 ± 0.64 | 0.31% | 66.67% | 0−1.25 |
Appendix B
Table A2.
Taxonomic classification and distribution of 18S rDNA clones retrieved from F. enflata juveniles sampled in summer and autumn in Sanya Bay.
Table A2.
Taxonomic classification and distribution of 18S rDNA clones retrieved from F. enflata juveniles sampled in summer and autumn in Sanya Bay.
| Clone ID | Best Hit Species | Best Hit ACC | E−Value | Similarity | Category | Percentage |
|---|---|---|---|---|---|---|
| W3−Sagen−Jul−j_diet | ||||||
| 1 | Cestum veneris | KJ754161.1 | 0 | 99% | Ctenophora | 5.56% |
| 2 | Pleurobrachia globosa | KJ859219.1 | 0 | 99% | Ctenophora | 2.78% |
| 3 | Temora turbinata | GU969211.1 | 0 | 98−99% | Copepoda | 55.56% |
| 4 | Temora turbinata | GU969211.1 | 0 | 95−96% | Copepoda | 11.11% |
| 5 | Temora turbinata | GU969211.1 | 0 | 90−92% | Copepoda | 5.56% |
| 6 | Canthocalanus pauper | GU969164.1 | 0 | 98−99% | Copepoda | 8.33% |
| 7 | Canthocalanus pauper | GU969164.1 | 0 | 91% | Copepoda | 2.78% |
| 8 | Canthocalanus pauper | GU969164.1 | 0 | 96% | Copepoda | 2.78% |
| 9 | Subeucalanus crassus | GU969168.1 | 0 | 96% | Copepoda | 2.78% |
| 10 | Ophiomusium cf. glabrum | KU519536.1 | 0 | 97% | Echinodermata | 2.78% |
| W4−Sagen−Jul−j_diet | ||||||
| 1 | Labidocera euchaeta | GU969153.1 | 0 | 96−98% | Copepoda | 6.45% |
| 2 | Labidocera euchaeta | GU969153.1 | 0 | 90% | Copepoda | 3.23% |
| 3 | Temora turbinata | GU969211.1 | 0 | 95−97% | Copepoda | 6.45% |
| 4 | Labidocera acuta | JQ280463.1 | 0 | 98% | Copepoda | 3.23% |
| 5 | Telepsavus spec. | AF448165.1 | 0 | 98% | Polychaeta | 3.23% |
| 6 | Spiochaetopterus bergensis | DQ209214.1 | 0 | 98% | Polychaeta | 3.23% |
| 7 | Chaetoceros debilis | AY229896.1 | 0 | 98−99% | Bacillariophyta | 79.97% |
| 8 | Chaetoceros debilis | AY229896.1 | 0 | 90% | Bacillariophyta | 3.23% |
| W9−Sagen−Jul−j_diet | ||||||
| 1 | Temora turbinata | GU969211.1 | 0 | 99%−100% | Copepoda | 46.81% |
| 2 | Temora turbinata | GU969211.1 | 0 | 95% | Copepoda | 2.13% |
| 3 | Candacia bispinosa | GU969213.1 | 0 | 99% | Copepoda | 6.38% |
| 4 | Canthocalanus pauper | GU969164.1 | 0 | 99% | Copepoda | 6.38% |
| 5 | Sulcanus conflictus | HM997064.1 | 0 | 99% | Copepoda | 2.13% |
| 6 | Tortanus gracilis | HM997065.1 | 0 | 97% | Copepoda | 2.13% |
| 7 | Cestum veneris | KJ754161.1 | 0 | 99% | Ctenophora | 12.77% |
| 8 | Pleurobrachia globosa | KJ859219.1 | 0 | 99% | Ctenophora | 4.26% |
| 9 | Ophiomusium cf. | KU519536.1 | 0 | 98% | Echinodermata | 12.77% |
| 10 | Chaetoceros sp. | FR865486.1 | 0 | 99% | Bacillariophyta | 4.26% |
| W3−Sagen−Oct−j_diet | ||||||
| 1 | Pleurobrachia globosa | KJ859219.1 | 0 | 98% | Ctenophora | 1.72% |
| 2 | Solmissus marshalli | AF358060.1 | 0 | 97−99% | Cnidaria | 53.45% |
| 3 | Sulculeolaria quadrivalvis | AY937329.1 | 0 | 97−99% | Cnidaria | 6.90% |
| 4 | Bougainvillia fulva | EU305490.1 | 0 | 98%% | Cnidaria | 1.72% |
| 5 | Subeucalanus crassus | GU969168.1 | 0 | 99% | Copepoda | 3.45% |
| 6 | Labidocera euchaeta | GU969153.1 | 0 | 92% | Copepoda | 1.72% |
| 7 | Paracalanus aculeatus | GU969180.1 | 0 | 99% | Copepoda | 1.72% |
| 8 | Candacia bispinosa | GU969213.1 | 0 | 99% | Copepoda | 1.72% |
| 9 | Tortanus gracilis | HM997065.1 | 0 | 99% | Copepoda | 1.72% |
| 10 | Phyllochaetopterus sp. | DQ209216.1 | 0 | 98−99% | Polychaeta | 12.07% |
| 11 | Gymnodinium mikimotoi | JF791035.1 | 0 | 97−98% | Dinophyceae | 5.17% |
| 12 | Karenia bidigitata | HM067002.1 | 0 | 90% | Dinophyceae | 1.72% |
| 13 | Karenia bidigitata | HM067002.1 | 0 | 95% | Dinophyceae | 1.72% |
| 14 | Karenia papilionacea | HM067005.1 | 0 | 95% | Dinophyceae | 1.72% |
| 15 | Amphidinium semilunatum | JQ179860.1 | 0 | 96% | Dinophyceae | 1.72% |
| 16 | Azadinium dexteroporum | KR362890.1 | 0 | 96% | Dinophyceae | 1.72% |
| W4−Sagen−Oct−a_diet | ||||||
| 1 | Pleurobrachia globosa | KJ859219.1 | 0 | 97−99% | Ctenophora | 23.53% |
| 2 | Pleurobrachia globosa | KJ859219.1 | 0 | 94% | Ctenophora | 1.96% |
| 3 | Diphyes dispar | AY937318.1 | 0 | 99−100% | Cnidaria | 15.69% |
| 4 | Solmissus marshalli | AF358060.1 | 0 | 98−99% | Cnidaria | 9.80% |
| 5 | Ectopleura obypa | KT722393.1 | 0 | 98% | Cnidaria | 1.96% |
| 6 | Euphysa aurata | EU876562.1 | 0 | 99% | Cnidaria | 1.96% |
| 7 | Lensia conoidea | AY937360.1 | 0 | 96% | Cnidaria | 1.96% |
| 8 | Muggiaea sp. | AF358073.1 | 0 | 96% | Cnidaria | 1.96% |
| 9 | Proboscidactyla flavicirrata | EU305500.1 | 0 | 99% | Cnidaria | 1.96% |
| 10 | Pachycerianthus sp. | AB859829.1 | 0 | 97% | Anthozoa | 1.96% |
| 11 | Subeucalanus crassus | GU969168.1 | 0 | 99% | Copepoda | 15.69% |
| 12 | Subeucalanus crassus | GU969168.1 | 0 | 92% | Copepoda | 1.96% |
| 13 | Paracalanus aculeatus | GU969180.1 | 0 | 97−99% | Copepoda | 7.84% |
| 14 | Candacia bispinosa | GU969213.1 | 0 | 97% | Copepoda | 1.96% |
| 15 | Labidocera euchaeta | GU969153.1 | 0 | 96% | Copepoda | 1.96% |
| 16 | Phyllochaetopterus sp. | DQ209216.1 | 0 | 99% | Polychaeta | 5.88% |
| 17 | Amphioplus cf. daleus | KU519529.1 | 0 | 98% | Echinodermata | 1.96% |
| W9−Sagen−Oct−a_diet | ||||||
| 1 | Bougainvillia fulva | EU305490.1 | 0 | 98−99% | Cnidaria | 97.83% |
| 2 | Amphioplus cf. daleus | KU519529.1 | 0 | 99% | Echinodermata | 2.17% |
Appendix C
Table A3.
Family of taxa genetically explored in the gut contents compared to the ambient zooplankton community.
Table A3.
Family of taxa genetically explored in the gut contents compared to the ambient zooplankton community.
| Category | Species | Summer | Autumn | ||
|---|---|---|---|---|---|
| Environment | Gut | Environment | Gut | ||
| Copepoda | Calanidae | √ | √ | √ | |
| Candaciidae | √ | √ | √ | √ | |
| Centropagidae | √ | √ | |||
| Paracalanidae | √ | √ | √ | ||
| Pontellidae | √ | √ | √ | √ | |
| Subeucalanidae | √ | √ | √ | √ | |
| Temoridae | √ | √ | √ | ||
| Tortanidae | √ | √ | √ | √ | |
| Sulcanidae | √ | ||||
| Anthozoa | Cerianthidae | √ | |||
| Cnidaria | Corymorphidae | √ | √ | √ | |
| Diphyidae | √ | √ | √ | ||
| Bougainvilliidae | √ | ||||
| Cuninidae | √ | ||||
| Proboscidactylidae | √ | ||||
| Tubulariidae | √ | ||||
| Ctenophora | Pleurobrachiidae | √ | √ | √ | |
| Cestidae | √ | ||||
| Echinodermata | Amphiuridae | √ | |||
| Ophiosphalmidae | √ | ||||
| Polychaeta | Chaetopteridae | √ | √ | ||
* √ means the species existed in the sample.
References
- Bone, Q.; Kapp, H.; Pierrot-Bults, A.C. The Biology of Chaetognaths; Oxford Universtity Press: New York, NY, USA, 1991. [Google Scholar]
- Kehayias, G.; Lykakis, J.; Fragopoulu, N. The diets of the chaetognaths Sagitta enflata, S. serratodentata atlantica and S. bipunctata at different seasons in Eastern Mediterranean coastal waters. ICES J. Mar. Sci. 1996, 53, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Øresland, V. Diel feeding of the chaetognath Sagitta enflata in the Zanzibar Channel, western Indian Ocean. Mar. Ecol. Prog. Ser. 2000, 193, 117–123. [Google Scholar] [CrossRef]
- Huo, Y.; Liu, Q.; Zhang, F.; Li, C.; Tao, Z.; Bi, H.; Fan, C.; Zhang, J.; Sun, S. Biomass and estimated production, and feeding pressure on zooplankton of chaetognaths in the Yellow Sea, China. Terr. Atmos. Ocean. Sci. 2020, 31, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Kimmerer, W. Selective predation and its impact on prey of Sagitta enflata (Chaetognatha). Mar. Ecol. Prog. Ser. 1984, 15, 55–62. [Google Scholar] [CrossRef]
- Johnson, T.B.; Nakagami, M.; Ueno, Y.; Narimatsu, Y.; Suyama, S.; Kurita, Y.; Sugisaki, H.; Terazaki, M. Chaetognaths in the diet of Pacific saury (Cololabis saira) in the northwestern Pacific Ocean. Coast. Mar. Sci. 2008, 32, 39–47. [Google Scholar]
- Sun, S.; Huo, Y.Z.; Yang, B. Zooplankton functional groups on the continental shelf of the yellow sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1006–1016. [Google Scholar] [CrossRef]
- Yamamura, O.; Honda, S.; Shida, O.; Hamatsu, T. Diets of walleye pollock Theragra chalcogramma in the Doto area, northern Japan: Ontogenetic and seasonal variations. Mar. Ecol. Prog. Ser. 2002, 238, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Dilling, L.; Alldredge, A.L. Can chaetognath fecal pellets contribute significantly to carbon flux? Mar. Ecol. Prog. Ser. 1993, 92, 51–58. [Google Scholar] [CrossRef]
- Giesecke, R.; González, H.E.; Bathmann, U. The role of the chaetognath Sagitta gazellae in the vertical carbon flux of the Southern Ocean. Polar Biol. 2010, 33, 293–304. [Google Scholar] [CrossRef]
- Baier, C.T.; Purcell, J.E. Trophic interactions of chaetognaths, larval fish, and zooplankton in the South Atlantic Bight. Mar. Ecol. Prog. Ser. 1997, 146, 43–53. [Google Scholar] [CrossRef]
- Coston-Clements, L.; Waggett, R.J.; Tester, P.A. Chaetognaths of the United States South Atlantic Bight: Distribution, abundance and potential interactions with newly spawned larval fish. J. Exp. Mar. Biol. 2009, 373, 111–123. [Google Scholar] [CrossRef]
- Kehayias, G. Spatial and temporal abundance distribution of chaetognaths in eastern Mediterranean pelagic waters. Bull. Mar. Sci. 2004, 74, 253–270. [Google Scholar]
- Tönnesson, K.; Tiselius, P. Diet of the chaetognaths Sagitta setosa and S. elegans in relation to prey abundance and vertical distribution. Mar. Ecol. Prog. Ser. 2005, 289, 177–190. [Google Scholar] [CrossRef]
- Matsuda, S.; Taniguchi, A. Diel changes in vertical distribution and feeding conditions of the chaetognath Parasagitta elegans (Verill) in the Subarctic Pacific in Summer. J. Oceanogr. 2001, 57, 353–360. [Google Scholar] [CrossRef]
- Choe, N.; Deibel, D. Seasonal vertical distribution and population dynamics of the chaetognath Parasagitta elegans in the water column and hyperbenthic zone of Conception Bay, Newfoundland. Mar. Biol. 2000, 137, 847–856. [Google Scholar] [CrossRef]
- Giesecke, R.; González, H.E. Reproduction and feeding of Sagitta enflata in the Humboldt Current system off Chile. ICES J. Mar. Sci. 2008, 65, 361–370. [Google Scholar] [CrossRef] [Green Version]
- King, K.R. The life history and vertical distribution of the chaetognath, Sagitta elegans, in Dabob Bay, Washington. J. Plankton Res. 1979, 1, 153–167. [Google Scholar] [CrossRef]
- Sameoto, D. Annual life cycle and production of the chaetognath Sagitta elegans in Bedford Basin, Nova Scotia. J. Fish. Board Can. 1973, 30, 333–344. [Google Scholar] [CrossRef]
- Welch, H.E.; Siferd, T.D.; Bruecker, P. Population densities, growth, and respiration of the chaetognath Parasagitta elegans in the Canadian high Arctic. Can. J. Fish. Aquat. Sci. 1996, 53, 520–527. [Google Scholar] [CrossRef]
- Marazzo, A.; Machado, C.F.; Nogueira, C.S.R. Notes on feeding of chaetognatha in Guanabara Bay, Brazil. J. Plankton Res. 1997, 19, 819–828. [Google Scholar] [CrossRef]
- Nagasawa, S.; Marumo, R. Further studies on the feeding habits of Sagitta nagae alvariño in Suruga Bay, central Japan. J. Oceanogr. 1976, 32, 209–218. [Google Scholar] [CrossRef]
- Alvarez-Cadena, J.N. Feeding of the Chaetognath Sagitta elegans Verrill. Estuar. Coast. Shelf Sci. 1993, 36, 195–206. [Google Scholar] [CrossRef]
- Pearre, S. Feeding by chaetognatha: Energy balance and importance of various components of the diet of Sagitta elegans. Mar. Ecol. Prog. Ser. 1981, 5, 45–54. [Google Scholar] [CrossRef]
- Grigor, J.J.; Marais, A.E.; Falk-Petersen, S.; Varpe, Ø. Polar night ecology of a pelagic predator, the chaetognath Parasagitta elegans. Polar Biol. 2015, 38, 87–98. [Google Scholar] [CrossRef]
- Grigor, J.J.; Schmid, M.S.; Caouette, M.; Onge, V.S.; Brown, T.A.; Barthélémy, R.-M. Non-carnivorous feeding in Arctic chaetognaths. Prog. Oceanogr. 2020, 186. [Google Scholar] [CrossRef]
- Jeanpaul Casanova, R.B.; Michel, D.; Eric, F. Chaetognaths feed primarily on dissolved and fine particulate organic matter, not on prey: Implications for marine food webs. Hypotheses Life Sci. 2012, 2, 20–29. [Google Scholar]
- Vinogradov, M.E.; Tseitlin, V.B. Deep-sea pelagic domain (aspects of bioenergetics). Deep Sea Biol. 1983, 8, 123–166. [Google Scholar]
- Yoon, H.; Ko, A.R.; Kang, J.H.; Choi, J.K.; Ju, S.J. Diet of Chaetognaths Sagitta crassa and S. nagae in the Yellow Sea inferred from gut content and fatty acid analyses. Ocean Polar Res. 2016, 38, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Kehayias, G. Quantitative aspects of feeding of chaetognaths in the eastern Mediterranean pelagic waters. J. Mar. Biol. Assoc. U.K. 2003, 83, 559–569. [Google Scholar] [CrossRef]
- Kehayias, G.; Ntakou, E. Abundance, vertical distribution and feeding of chaetognaths in the upper 50m layer of the eastern Aegean Sea. J. Nat. Hist. 2008, 42, 633–648. [Google Scholar] [CrossRef]
- Pearre, S. Ecological studies of three west-mediterranean chaetognaths. Investig. Pesq. 1974, 38, 325–369. [Google Scholar]
- Kehayias, G.; Michaloudi, E.; Koutrakis, E. Feeding and predation impact of chaetognaths in the north Aegean Sea (Strymonikos and Ierissos Gulfs). J. Mar. Biol. Assoc. U.K. 2005, 85, 1525–1532. [Google Scholar] [CrossRef]
- King, R.A.; Read, D.S.; Traugott, M.; Symondson, W.O.C. Molecular analysis of predation: A review of best practice for DNA-based approaches. Mol. Ecol. 2008, 17, 947–963. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Guo, Z.; Li, T.; Xu, C.; Huang, H.; Liu, S.; Lin, S. Molecular analysis of in situ diets of coral reef copepods: Evidence of terrestrial plant detritus as a food source in Sanya Bay, China. J. Plankton Res. 2015, 37, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Hu, S.; Sheng, L.; Hui, H. Unexpected prey of juvenile spotted scat (Scatophagus argus) near a wharf: The prevalence of fouling organisms in stomach contents. Ecol. Evol. 2018, 8, 8547–8554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, D.; Lindeque, P.K.; Harris, R.P. Sagitta setosa predation on Calanus helgolandicus in the English Channel. J. Plankton Res. 2010, 32, 725–737. [Google Scholar] [CrossRef]
- Huang, L.M.; Tan, Y.H.; Song, X.Y.; Huang, X.P.; Wang, H.K.; Zhang, S.; Dong, J.; Chen, R. The status of the ecological environment and a proposed protection strategy in Sanya Bay, Hainan Island, China. Mar. Pollut. Bull. 2003, 47, 180. [Google Scholar] [CrossRef]
- Shi, X.; Wang, H.; Tan, Y.; Huang, L. Seasonal variation of zooplankton community structure and species composition in the Sanya Bay. Mar. Sci. Bull. 2007, 26, 42–49. [Google Scholar] [CrossRef]
- Yin, J.Q.; Zhang, G.X.; Tan, Y.H.; Huang, L.M.; Kai-Zhi, L.I. Species composition and quantitative distribution of zooplankton in Sanya Bay, Hainan Province, China. J. Trop. Oceanogr. 2004, 23, 1–9. [Google Scholar] [CrossRef]
- Li, K.Z.; Wu, X.J.; Tan, Y.H.; Hui, H.; Dong, J.D.; Huang, L.M. Spatial and temporal variability of copepod assemblages in Sanya Bay, northern South China Sea. Reg. Stud. Mar. Sci. 2016, 7, 168–176. [Google Scholar] [CrossRef]
- Guo, Z.L.; Liu, S.; Hu, S.M.; Li, T.; Huang, Y.S.; Liu, G.X.; Zhang, H.; Lin, S.J. Prevalent ciliate symbiosis on copepods: High genetic diversity and wide distribution detected using small subunit ribosomal RNA gene. PLoS ONE 2012, 7, e44847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Guo, Z.; Li, T.; Carpenter, E.J.; Liu, S.; Lin, S. Detecting in situ copepod diet diversity using molecular technique: Development of a copepod/symbiotic ciliate-excluding eukaryote-inclusive PCR protocol. PLoS ONE 2014, 9, e103528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.W.; Hong, H.X.; Zhang, S.M. Fauna Sinica; Science Press: Beijing, China, 2002; Volume 27. [Google Scholar]
- Kehayias, G.; Koutsikopoulos, C.; Fragopoulu, N.; Lykakis, J. A single maturity classification key for five common Mediterranean chaetognath species. J. Mar. Biol. Assoc. U.K. 1999, 79, 1137–1138. [Google Scholar] [CrossRef]
- Lie, A.A.Y.; Tse, P.; Wong, C.K. Diel vertical migration and feeding of three species of chaetognaths (Flaccisagitta enflata, Aidanosagitta delicata and Aidanosagitta neglecta) in two shallow, subtropical bays in Hong Kong. J. Plankton Res. 2012, 34, 670–684. [Google Scholar] [CrossRef]
- Zhang, H.; Lin, S.J. Development of a cob-18S rRNA gene real-time PCR assay for quantifying Pfiesteria shumwayae in the natural environment. Appl. Environ. Microbiol. 2005, 71, 7053–7063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giesecke, R.; Gonzalez, H.E. Distribution and feeding of chaetognaths in the epipelagic zone of the Lazarev Sea (Antarctica) during austral summer. Polar Biol. 2012, 35, 689–703. [Google Scholar] [CrossRef]
- Øresland, V. Feeding of the chaetognaths Sagitta elegans and S. setosa at different seasons in Gullmarsfjorden, Sweden. Mar. Ecol. Prog. Ser. 1987, 39, 69–79. [Google Scholar] [CrossRef]
- Giesecke, R.; Gonzalez, H.E. Feeding of Sagitta enflata and vertical distribution of chaetognaths in relation to low oxygen concentrations. J. Plankton Res. 2004, 26, 475–486. [Google Scholar] [CrossRef]
- Sifford, P.J. A seasonal study of the diets of three sympatric chaetognaths. Investig. Pesq. 1976, 40, 1–16. [Google Scholar]
- Terazaki, M. Feeding of Carnivorous Zooplankton, Chaetognaths in the Pacific; Springer: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Pearre, S. Feeding by Chaetognatha: The relation of prey size to predator size in several species. Mar. Ecol. Prog. Ser. 1980, 3, 125–134. [Google Scholar] [CrossRef]
- Reeve, M.; Walter, M. Conditions of culture, food-size selection, and the effects of temperature and salinity on growth rate and generation time in Sagitta hispida Conant. J. Exp. Mar. Biol. Ecol. 1972, 9, 191–200. [Google Scholar] [CrossRef]
- Dong, D.; Li, X.Z.; Wang, H.F.; Zhang, B.L.; Kou, Q.; Gan, Z.B.; Xu, P. Macrobenthic community characters of coral reef at Sanya, Hainan. Mar. Sci. 2015, 39, 83–91. [Google Scholar] [CrossRef]
- Huang, L.M.; Zhang, S.; Wang, H.K.; Wen, W.; Zhang, Q. Ecological Environment and Bioresources for Sanya Bay, Hainan Island, China; Science Press: Beijing, China, 2007. [Google Scholar]
- Blanchard, G.F.; Paterson, D.M.; Stal, L.J.; Richard, P.; Galois, R.; Huet, V.; Kelly, J.; Honeywill, C.; de Brouwer, J.; Dyer, K.; et al. The effect of geomorphological structures on potential biostabilisation by microphytobenthos on intertidal mudflats. Cont. Shelf Res. 2000, 20, 1243–1256. [Google Scholar] [CrossRef]
- Hendler, G. Development of Amphioplus abditus (Verrill) (Echinodermata: Ophiuroidea): I. Larval Biology. Biol. Bull. 1977, 152, 51–63. [Google Scholar] [CrossRef]
- Nagasawa, S. The digestive efficiency of the chaetognath Sagitta crassa Tokioka, with observations on the feeding process. J. Exp. Mar. Biol. Ecol. 1985, 87, 271–282. [Google Scholar] [CrossRef]
- Reeve, M. Complete cycle of development of a pelagic chaetognath in culture. Nature 1970, 227, 381. [Google Scholar] [CrossRef]
- Schoener, A. Post-larval development of five deep-sea ophiuroids. Deep Sea Res. Oceanogr. Abstr. 1967, 14, 645–660. [Google Scholar] [CrossRef]
- Wyngaard, G.A.; Rasch, E.M. Patterns of genome size in the copepoda. Hydrobiologia 2000, 417, 43–56. [Google Scholar] [CrossRef]
- Moroz, L.L.; Kocot, K.M.; Citarella, M.R.; Dosung, S.; Norekian, T.P.; Povolotskaya, I.S.; Grigorenko, A.P.; Dailey, C.; Berezikov, E.; Buckley, K.M.; et al. The ctenophore genome and the evolutionary origins of neural systems. Nature 2014, 510, 109–114. [Google Scholar] [CrossRef]
- Kruse, S.; Hagen, W.; Bathmann, U. Feeding ecology and energetics of the Antarctic chaetognaths Eukrohnia hamata, E. bathypelagica and E. bathyantarctica. Mar. Biol. 2010, 157, 2289–2302. [Google Scholar] [CrossRef]
- Buskey, E.J.; Lenz, P.H.; Hartline, D.K. Escape behavior of planktonic copepods in response to hydrodynamic disturbances: High speed video analysis. Mar. Ecol. Prog. Ser. 2002, 235, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Bae, B.S.; Kim, I.O.; Yoon, W.D. Measurement of swimming speed of giant jellyfish Nemopilema nomurai using acoustics and visualization analysis. Fish. Sci. 2010, 76, 893–899. [Google Scholar] [CrossRef]
- Doyle, T.K.; Houghton, J.D.R.; Mcdevitt, R.; Davenport, J.; Hays, G.C. The energy density of jellyfish: Estimates from bomb-calorimetry and proximate-composition. J. Exp. Mar. Biol. 2007, 343, 239–252. [Google Scholar] [CrossRef]
- Liu, C.S.; Chen, S.Q.; Zhuang, Z.M.; Yan, J.P.; Liu, C.L.; Cui, H.T. Potential of utilizing jellyfish as food in culturing Pampus argenteus juveniles. Hydrobiologia 2015, 754, 189–200. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Sato, R.; Ishii, H.; Akiba, T.; Nogata, Y.; Tanaka, Y. Culture of phyllosomas of Ibacus novemdentatus (Decapoda: Scyllaridae) in a closed recirculating system using jellyfish as food. Aquaculture 2012, 330, 162–166. [Google Scholar] [CrossRef]
- Liu, J.; Li, C.C.; Li, X.S. Phylogeny and biogeography of Chinese pomfret fishes (Pisces: Stromateidae). Studia Mar. Sin. 2002, 2002, 235–239. [Google Scholar]
- Miyajima, Y.; Masuda, R.; Kurihara, A.; Kamata, R.; Yamashita, Y.; Takeuchi, T. Juveniles of threadsail filefish, Stephanolepis cirrhifer, can survive and grow by feeding on moon jellyfish Aurelia aurita. Fish. Sci. 2011, 77, 41–48. [Google Scholar] [CrossRef]
- Boero, F.; Brotz, L.; Gibbons, M.J.; Piraino, S.; Zampardi, S. Explaining Ocean Warming: Causes, Scale, Effects and Consequences; IUCN: Gland, Switzerland, 2016; pp. 213–237. [Google Scholar]
- Marques, R.; Bouvier, C.; Darnaude, A.M.; Molinero, J.-C.; Przybyla, C.; Soriano, S.; Tomasini, J.-A.; Bonnet, D. Jellyfish as an alternative source of food for opportunistic fishes. J. Exp. Mar. Biol. Ecol. Evol. 2016, 485, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Olesen, N.J. Clearance potential of jellyfish Aurelia aurita, and predation impact on zooplankton in a shallow cove. Mar. Ecol. Prog. Ser. 1995, 124, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Fleming, N.E.; Harrod, C.; Newton, J.; Houghton, J.D. Not all jellyfish are equal: Isotopic evidence for inter-and intraspecific variation in jellyfish trophic ecology. PeerJ 2015, 3, e1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Sampling stations in Sanya Bay separately visited on July and October, 2014.

Figure 2.
Zooplankton community on the sampling date of July and October, 2014.

Figure 3.
Phylogenetic affiliations, heatmap and composition of the diet organisms of juvenile F. enflata in two seasons. Maximum Likelihood (ML) tree was inferred from 780 bp fragment of the 18S rDNA amplified from the gut.
Figure 3.
Phylogenetic affiliations, heatmap and composition of the diet organisms of juvenile F. enflata in two seasons. Maximum Likelihood (ML) tree was inferred from 780 bp fragment of the 18S rDNA amplified from the gut.

Figure 4.
Schematic of the main food groups for juveniles chaetognath in Sanya Bay. (Copepods including copepod nauplii and copepodites; Small jellyfish could be fed by juveniles chaetognath in forms of planula, polyps, medusa and detritus).
Figure 4.
Schematic of the main food groups for juveniles chaetognath in Sanya Bay. (Copepods including copepod nauplii and copepodites; Small jellyfish could be fed by juveniles chaetognath in forms of planula, polyps, medusa and detritus).

Table 1.
Environmental parameters sampled in 2014 in the northern South China Sea.
| Station | Sampling Date | Temperature (°C) | Salinity (‰) | pH | Dissolved Oxygen (mg L−1) | Dissolved Organic Carbon (mg L−1) |
|---|---|---|---|---|---|---|
| W3 | 29 July | 27.64 | 34.98 | 8.13 | 6.37 | 4.17 |
| W4 | 29 July | 27.83 | 35.00 | 8.15 | 6.36 | 3.39 |
| W9 | 29 July | 27.86 | 34.89 | 8.14 | 6.25 | 3.96 |
| W3 | 26 October | 28.16 | 33.22 | 8.16 | 6.62 | 1.83 |
| W4 | 26 October | 28.04 | 33.17 | 8.16 | 6.56 | 1.77 |
| W9 | 26 October | 28.25 | 33.58 | 8.16 | 6.53 | 1.80 |
Table 2.
Diversity indices of prey organisms in the F. enflata juveniles samples analyzed.
| Sample ID * | Taxa | Individuals/Clones | Simpson_D | Shannon_H | Chao1 |
|---|---|---|---|---|---|
| W3-Jul-J | 10 | 36 | 0.662 | 1.597 | 13.33 |
| W4-Jul-J | 8 | 31 | 0.4828 | 1.151 | 11.33 |
| W9-Jul-J | 10 | 47 | 0.7352 | 1.747 | 11 |
| W3-Oct-J | 16 | 58 | 0.6879 | 1.814 | 43.5 |
| W4-Oct-J | 17 | 51 | 0.872 | 2.364 | 72 |
| W9-Oct-J | 2 | 46 | 0.04253 | 0.1047 | 2 |
* Jul, July; Oct, October; J, juveniles. Simpson’s diversity index and Shannon-Wiener index reflect the diversity of prey organisms. Chao1 is used to estimate the number of prey OTUs (operational taxonomic units) in the community.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, L.; Guo, M.; Li, T.; Huang, H.; Liu, S.; Hu, S. Small Jellyfish as a Supplementary Autumnal Food Source for Juvenile Chaetognaths in Sanya Bay, China. J. Mar. Sci. Eng. 2020, 8, 956. https://doi.org/10.3390/jmse8120956
AMA Style
Wang L, Guo M, Li T, Huang H, Liu S, Hu S. Small Jellyfish as a Supplementary Autumnal Food Source for Juvenile Chaetognaths in Sanya Bay, China. Journal of Marine Science and Engineering. 2020; 8(12):956. https://doi.org/10.3390/jmse8120956
Chicago/Turabian StyleWang, Lingli, Minglan Guo, Tao Li, Hui Huang, Sheng Liu, and Simin Hu. 2020. "Small Jellyfish as a Supplementary Autumnal Food Source for Juvenile Chaetognaths in Sanya Bay, China" Journal of Marine Science and Engineering 8, no. 12: 956. https://doi.org/10.3390/jmse8120956
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.