

The First Report of the Acrotretoid Brachiopod Hadrotreta from the Tsinghsutung Formation Cambrian (Series 2, Stage 4), Guizhou, South China
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
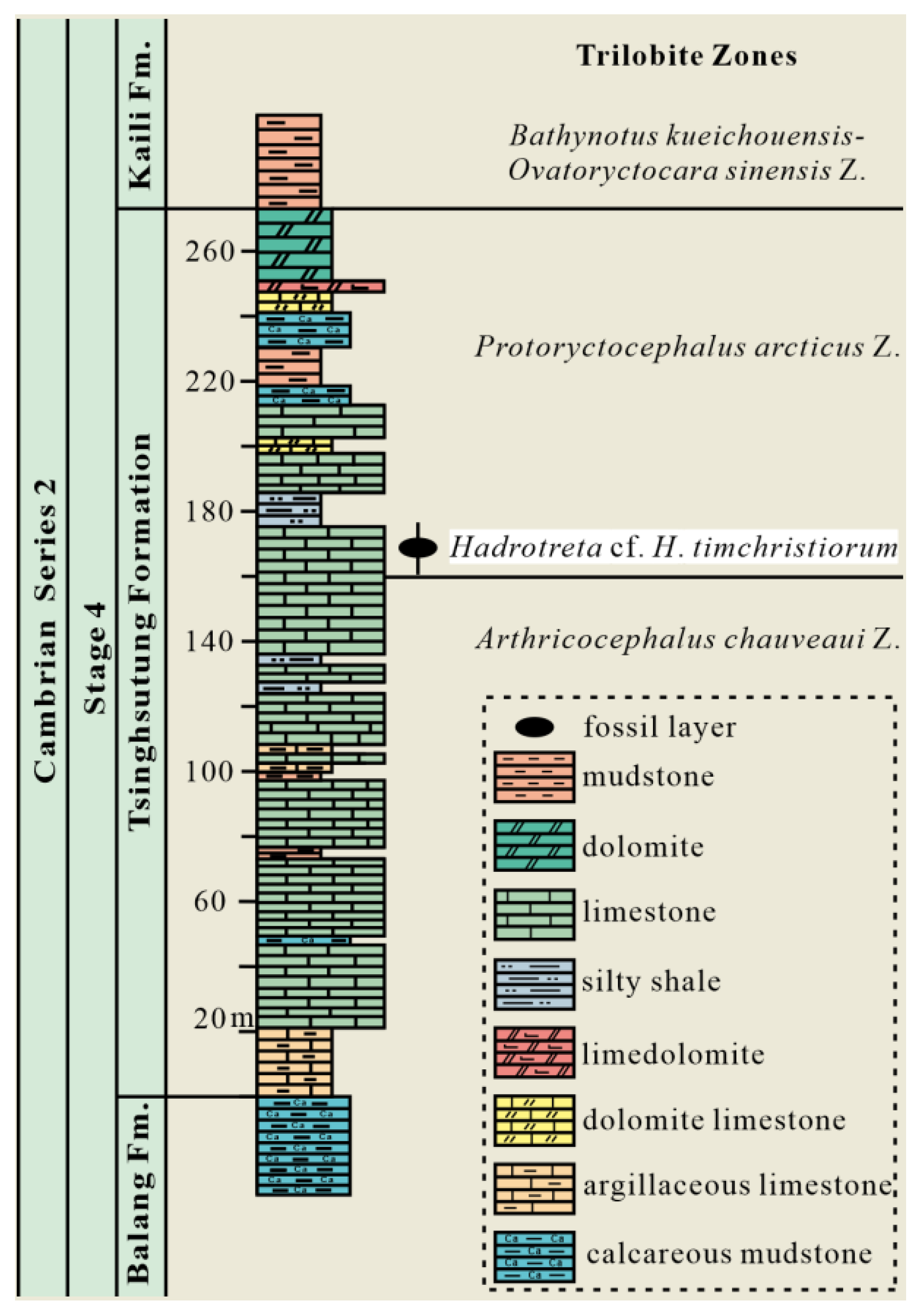
2.1. Geological Setting
2.2. Hadrotreta Material and Specimen Visualization
3. Results and Discussion
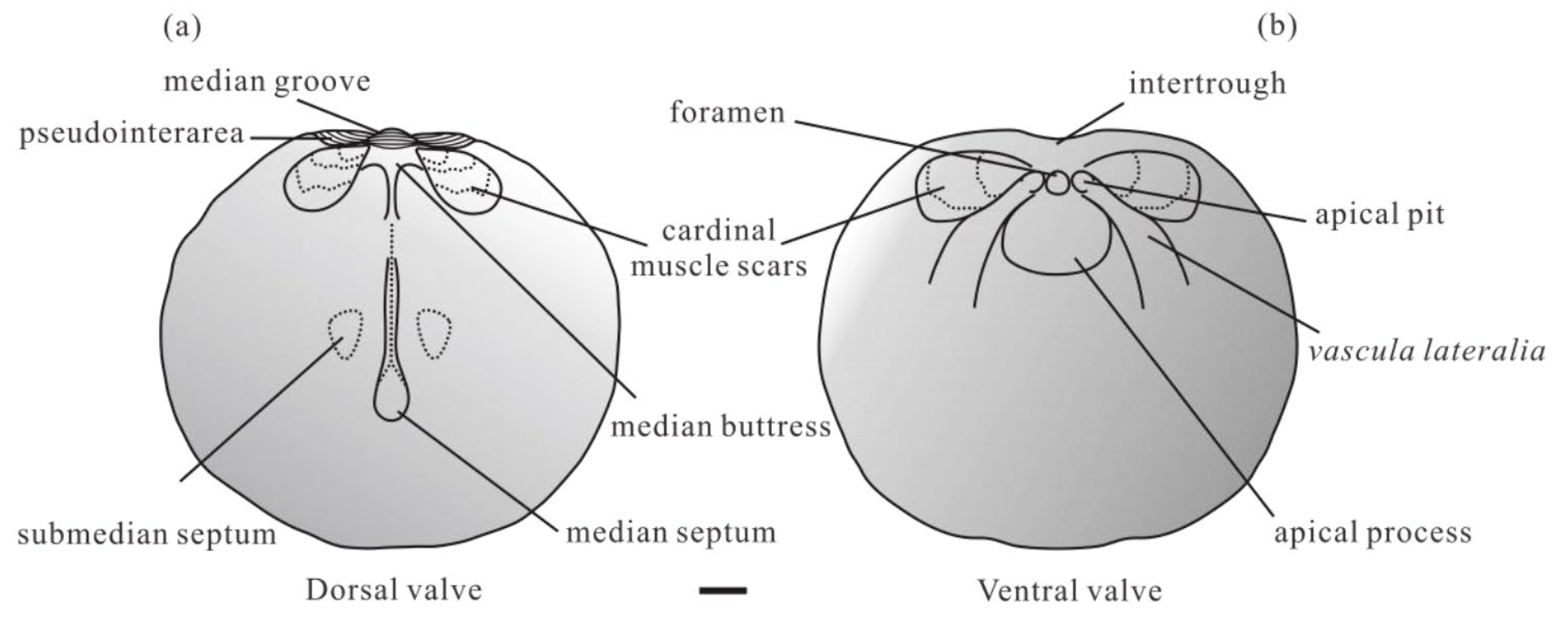
3.1. Systematic Palaeontology
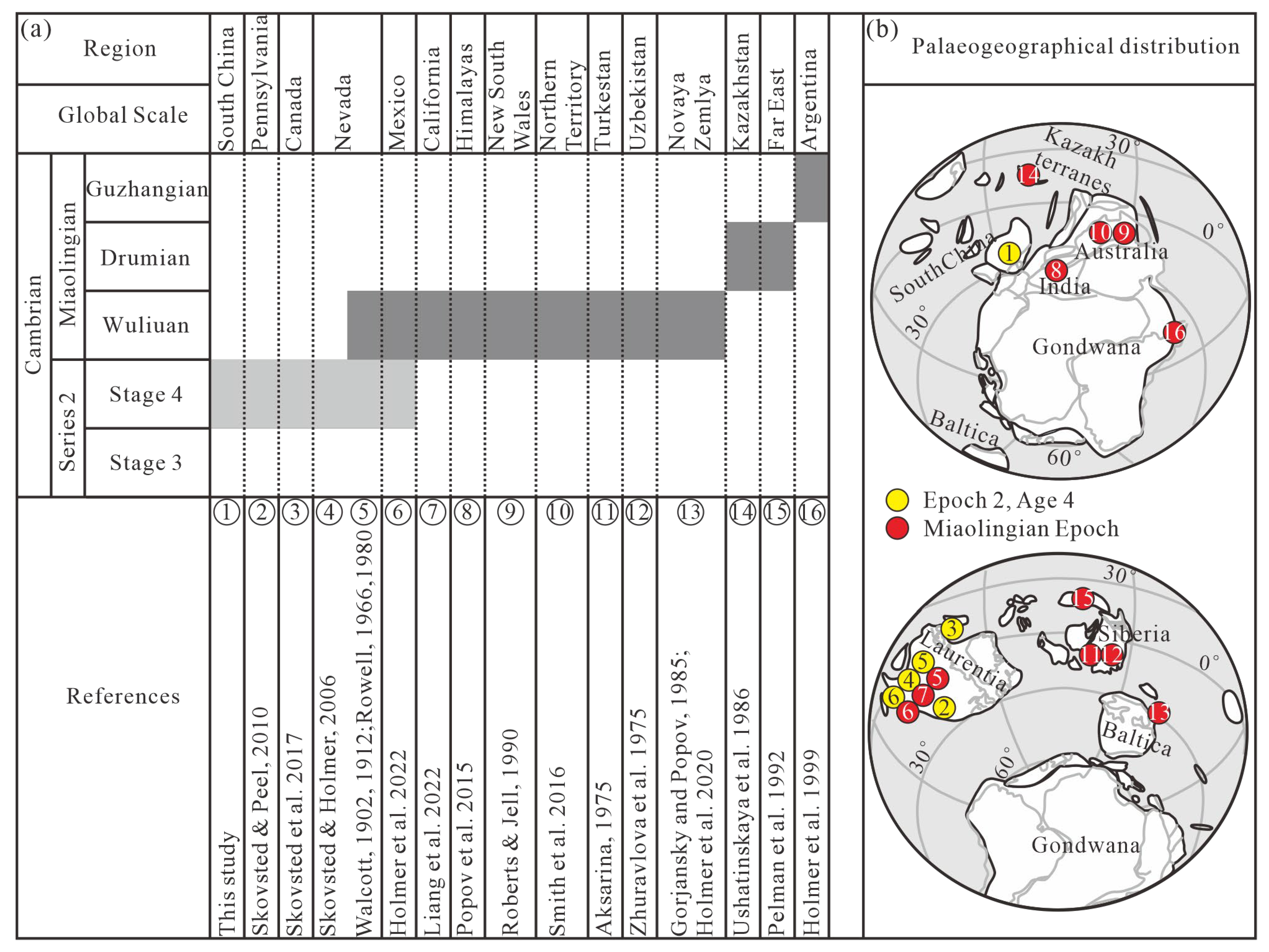
3.2. Palaeogeographical Implication of Hadrotreta
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bassett, M.G.; Popov, L.E.; Holmer, L.E. Organophosphatic brachiopods: Patterns of biodiversification and extinction in the early Palaeozoic. Geobios 1999, 32, 145–163. [Google Scholar]
- Ushatinskaya, G.T. Stratigraphic and Geographic Distribution of Acrotretids (Brachiopoda, Lingulata) in the Middle and Late Cambrian. Paleontol. J. 2010, 44, 1164–1175. [Google Scholar]
- Holmer, L.E.; Popov, L.E. Family Acrotretida. In Treatise on Invertebrate Paleontology, Part H, Brachiopoda; Kaesler, R.L., Ed.; Geological Society of America, Inc.: Boulder, CO, USA; University of Kansas: Boulder, CO, USA; Lawrence, KS, USA, 2000; Volume 2, pp. 97–122. [Google Scholar]
- Wang, H.Z.; Zhang, Z.F.; Holmer, L.E.; Hu, S.X.; Wang, X.R.; Li, G.X. Peduncular attached secondary tiering acrotretoid brachiopods from the chengjiang fauna: Implications for the ecological expansion of brachiopods during the cambrian explosion. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 323–325, 60–67. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Zhang, Z.F.; Holmer, L.E. Studies on the shell ultrastructure and ontogeny of the oldest acrotretid brachiopods from South China. Acta Palaeontol. Sin. 2017, 56, 483–503. (In Chinese) [Google Scholar]
- Zhang, Z.L.; Holmer, L.E.; Chen, F.Y.; Brock, G.A. Ontogeny and evolutionary significance of a new acrotretide brachiopod genus from Cambrian Series 2 of South China. J. Syst. Palaeontol. 2020, 18, 1569–1588. [Google Scholar] [CrossRef]
- Duan, X.L.; Liang, Y.; Holmer, L.E.; Zhang, Z.F. First report of acrotretoid brachiopod shell beds in the lower Cambrian (Stage 4) Guanshan Biota of eastern Yunnan, South China. J. Paleontol. 2020, 95, 40–55. [Google Scholar] [CrossRef]
- Li, G.X.; Holmer, L.E. Early Cambrian lingulate brachiopods from the Shaanxi Province, China. GFF 2004, 126, 193–211. [Google Scholar]
- Percival, I.G.; Kruse, P.D.; Laurie, J.R.; Paterson, J.R.; Brock, G.A. Middle Cambrian brachiopods from the southern Georgina Basin of central Australia. Mem. Assoc. Australas. Palaeontol. 2014, 45, 349–402. [Google Scholar]
- Streng, M.; Holmer, L.E. Discovery of a new type of shell structure within the organophosphatic brachiopods and the status of the family Curticiidae. GFF 2005, 127, 7–16. [Google Scholar] [CrossRef]
- Winrow, P.; Sutton, M.D. Epithelial cell moulds in acrotretoid brachiopods. Hist. Biol. 2012, 24, 557–565. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Zhang, Z.F.; Holmer, L.E.; Chen, F.Y. Post-metamorphic allometry in the earliest acrotretoid brachiopods from the lower Cambrian (Series 2) of South China, and its implications. Palaeontology 2018, 61, 183–207. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.L.; Popov, L.E.; Holmer, L.E.; Zhang, Z.F. Earliest ontogeny of early Cambrian acrotretoid brachiopods—First evidence for metamorphosis and its implications. BMC Evol. Biol. 2018, 18, 42. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.; Holmer, L.E. Ornamentation and Shell Structure of Acrotretoid Brachiopods. Palaeontology 1992, 35, 657–692. [Google Scholar]
- Duan, X.L.; Betts, M.J.; Holmer, L.E.; Chen, Y.L.; Liu, F.; Liang, Y.; Zhang, Z.F. Early Cambrian (Stage 4) brachiopods from the Shipai Formation in the Three Gorges area of South China. J. Paleontol. 2021, 95, 497–526. [Google Scholar] [CrossRef]
- Aksarina, N.A. Brachiopods. In Lower Palaeozoic Stratigraphy and Fauna of the Northern Slopes of the Turkestan and Alai Ranges; Repina, L.N., Jaskovich, B.V., Aksarina, N.A., Petrunina, Z.E., Poniklenko, I.A., Rubanov, D.A., Bolgova, G.V., Golikov, A.N., Khajrulina, T.I., et al., Eds.; Trudy Instituta Geologii i Geofziki, Akademiya Nauk SSSR Sibirskoye Otdelenye: Saint Petersburg, Russia, 1975; pp. 91–100. (In Russian) [Google Scholar]
- Zhuravlova, I.T.; Belyaeva, G.V.; Luchinina, V.A.; Nazarov, B.B.; Repina, L.N.; Sobolev, L.P. Kembriiskaia Fauna i Flora Khrebta Dzhagdy (Dal’nii Vostok); Akademiia Nauk SSSR, Sibirskoe Otdelenie; Institut Geologii i Geofiziki (IGIG), Trudy (Novosibirsk): Saint Petersburg, Russia, 1975; Volume 226, pp. 1–208. [Google Scholar]
- Rowell, A.J. Inarticulate brachiopods of the lower and middle Cambrian Pioche Shale of the Pioche District, Nevada. Univ. Kans. Paleontol. Contrib. 1980, 98, 1–26. [Google Scholar]
- Ushatinskaya, G.T.; Gidaspov, A.D.; Ryazantsev, A.V. A locality of Middle Cambrian Inarticulate Brachiopods in Northeastern Kazakhstan. Paleontol. Zhurnal 1986, 3, 35–40. [Google Scholar]
- Roberts, J.; Jell, P.A. Early middle Cambrian (Ordian) brachiopods of the Coonigan Formation, western New South Wales. Alcheringa Australas. J. Palaeontol. 1990, 14, 257–309. [Google Scholar] [CrossRef]
- Pelman, Y.L.; Aksarina, N.A.; Koneva, S.P. The Earliest Brachiopods from the Territory of Northern Eurasia; Oighim Ras: Novosibirsk, Russia, 1992; pp. 42–144. (In Russian) [Google Scholar]
- Skovsted, C.B.; Peel, J.S. Brachiopods and small shelly fossils from The Basal Kinzers Formation (Cambrian Series 2, Stage 4) of Pennsylvania. J. Paleontol. 2010, 84, 754–762. [Google Scholar] [CrossRef]
- Popov, L.E.; Holmer, L.E.; Hughes, N.C.; Ghobadi Pour, M.; Myrow, P.M. Himalayan Cambrian brachiopods. Pap. Palaeontol. 2015, 1, 345–399. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.M.; Brock, G.A.; Paterson, J.R. Linguliformean brachiopods from the early Templetonian (Cambrian Series 3, Stage 5) Giles Creek Dolostone, Amadeus Basin, Northern Territory. Australas. Palaeontol. Mem. 2016, 49, 125–143. [Google Scholar]
- Skovsted, C.B.; Knight, I.; Balthasar, K.U.; Boyce, W.D. Depth related brachiopod faunas from the lower Cambrian Forteau Formation of southern Labrador and western Newfoundland, Canada. Palaeontol. Electron. 2017, 20, 54A. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmer, L.E.; Blodgett, R.B.; Liang, Y.; Zhang, Z.F. The Early Devonian (Emsian) acrotretid microbrachiopod Opsiconidion minor Popov, 1981, from the Alaska/Yukon Territory border and Novaya Zemlya. Est. J. Earth Sci. 2020, 69, 143. [Google Scholar] [CrossRef]
- Holmer, L.E.; Clausen, S.; Popov, L.E.; Ghobadi Pour, M.; Liang, Y.; Zhang, Z.F.; Palafox Reyes, J.J.; Sosa-Leon, J.P.; Buitrón-Sánchez, B.E. Cambrian (Stage 4 to Wuliuan) brachiopods from Sonora, Mexico. J. Paleontol. 2022, 96, 1264–1284. [Google Scholar] [CrossRef]
- Liang, Y.; Holmer, L.E.; Duan, X.L.; Zhang, Z.F. Brachiopods from the Latham Shale Lagerstätte (Cambrian Series 2, Stage 4) and Cadiz Formation (Miaolingian, Wuliuan), California. J. Paleontol. 2022, 96, 61–80. [Google Scholar] [CrossRef]
- Holmer, L.E.; Popov, L.E.; Lehnert, O. Cambrian phosphatic brachiopods from the Precordillera of western Argentina. GFF 1999, 121, 227–242. [Google Scholar] [CrossRef]
- Sang, T.; Wang, L.T. General Paleogeographic Situations. In Regional Geology of Guizhou Province; Burcan of Guizhou Geology, Mineral Resources, Ed.; Geological Publishing House: Beijing, China, 1987; pp. 480–483. [Google Scholar]
- Zhao, Y.L.; Peng, J.; Yuan, J.L.; Guo, Q.J.; Tai, T.S.; Yin, L.M.; Parsley, L.R.; Yang, Y.N.; Yang, X.L.; Zhang, P.X. The Kaili Formation and Kaili Biota at the Wuliu-Zengjiayan section of Guizhou Province, China and proposed Global Standard Stratotype-section and Point, (GSSP) of the unnamed Cambrian Series 3, Stage 5. J. Guizhou Univ. 2012, 29, 108–124. [Google Scholar]
- Zhao, Y.L.; Yuan, J.L.; Babcock, L.E.; Guo, Q.J.; Peng, J.; Yin, L.M.; Yang, X.L.; Peng, S.C.; Wang, C.J.; Gaines, R.R.; et al. Global Standard Stratotype-Section and Point (GSSP) for the conterminous base of the Miaolingian Series and Wuliuan Stage (Cambrian) at Balang, Jianhe, Guizhou, China. Episodes 2019, 42, 165–184. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yang, X.L.; Zhao, Y.L.; Cao, P. Tubular Skeletal Microfossils from the Cambrian “Tsinghsutung Formation” of Guizhou, China. Acta Palaeontol. Sin. 2019, 58, 178–187. (In Chinese) [Google Scholar]
- Zhang, Y.; Zhao, Y.L. Preliminary analysis of sedimentary environment from Cambrian “Tsinghsutung Formation” in Balang area, Jianhe county, Guizhou Province-case study of Songshan section from Balang Village, Jianhe county. Acta Palaeontol. Sin. 2018, 57, 11–20. (In Chinese) [Google Scholar]
- Jeppsson, L.; Fredholm, D.; Mattiasson, B.O. Acetic acid and phosphatic fossils: A warning. J. Paleontol. 1985, 59, 952–956. [Google Scholar]
- Williams, A.; Carlson, S.J.; Bruton, C.H.C.; Holmer, L.E.; Popov, L.E. A Supra-Ordinal Classification of the Brachiopoda. Philos. Trans. R. Soc. Lond. Ser. B 1996, 351, 1171–1193. [Google Scholar] [CrossRef]
- Gorjansky, V.I.; Popov, L.E. The morphology, systematic position, and origin of inarticulate brachiopods with carbonate shells. Paleontol. J. 1985, 3, 1–11. [Google Scholar]
- Kuhn, O. Lehrbuch der Paläozoologie; E. Schweizerbart’sch Verlagsbuchhandlung: Stuttgart, Germany, 1949; p. 326. [Google Scholar]
- Schuchert, C. A classification of the Brachiopoda. Am. Geol. 1893, 11, 141–167. [Google Scholar]
- Rowell, A.J. Revision of some Cambrian and Ordovician inarticulate brachiopods. Univ. Kans. Paleontol. Contrib. 1966, 7, 1–36. [Google Scholar]
- Walcott, C.D. Cambrian Brachiopoda: Acrotreta; Linnarssonella; Obolus; with descriptions of new species. Proc. U. S. Natl. Mus. 1902, 25, 577–612. [Google Scholar] [CrossRef] [Green Version]
- Walcott, C.D. Cambrian Brachiopoda. U. S. Geol. Surv. Monogr. 1912, 51, 700. [Google Scholar]
- Popov, L.E.; Holmer, L.E.; Gorjansky, V.J. Middle Cambrian lingulate brachiopods from the Tarbagatay Range, Kazakhstan. Acta Palaeontol. Pol. 1996, 41, 299–317. [Google Scholar]
- Smith, P.M.; Brock, G.A.; Paterson, J.R. Fauna and biostratigraphy of the Cambrian (Series 2, Stage 4; Ordian) Tempe Formation (Pertaoorrta Group), Amadeus Basin, Northern Territory. Alcheringa Australas. J. Palaeontol. 2015, 39, 40–70. [Google Scholar] [CrossRef]
- Mergl, M. Inarticulate brachiopods of early middle Cambrian age from the High Atlas, Morocco. Vestn. Ustred. Ustavu Geol. 1988, 63, 291–295. [Google Scholar]
- Holmer, L.E.; Popov, L.E.; Wrona, R. Early Cambrian lingulate brachiopods from glacial erratics of King George Island (South Shetland Islands), Antarctica. Palaeontol. Pol. 1996, 55, 37–50. [Google Scholar]
- Walcott, C.D. Paleontologic notes. Am. J. Sci. 1885, 29, 114–117. [Google Scholar] [CrossRef]
- Holmer, L.E.; Popov, L.E.; Koneva, S.P.; Bassett, M.G. Cambrian-Early Ordovician brachiopods from Malyi Karatau, the western Balkhash region, and Tien Shan, central Asia. Palaeontol. Assoc. Spec. Pap. Palaeontol. 2001, 65, 1–180. [Google Scholar]
- Streng, M. Early middle Cambrian representatives of the superfamily Acrotretoidea (Brachiopoda) from Morocco. Z. Dtsch. Gelogischen Ges. 1999, 150, 27–87. [Google Scholar]
- Holmer, L.E.; Popov, L.E. Organophosphatic bivalve stem-group brachiopods. In Treatise on Invertebrate Paleontology, Part H, Brachiopoda, Revised; Williams, A., Brunton, C.H.C., Carlson, S.J., Eds.; Geological Society of America Inc.: Boulder, CO, USA; University of Kansas Press: Boulder, CO, USA; Lawrence, KS, USA, 2007; Volume 6, pp. 2581–2590. [Google Scholar] [CrossRef]
- Kruse, P.D. Cambrian Paleontology of the Daly Basin; Northern Territory Geological Survey: Darwin, Australia, 1990; pp. 28–30. [Google Scholar]
- Skovsted, C.B.; Holmer, L.E. The lower Cambrian brachiopod Kyrshabaktella and associated shelly fossils from the Harkless Formation, southern Nevada. GFF 2006, 128, 327–337. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Hadrotreta | Kostjubella | Vandalotreta | Linnarssonia | Eohadrotreta |
|---|---|---|---|---|---|
| Ventral valve | Shell transversely oval, ventral valve moderately convex to subconical with maximum height at the umbo or beak | Ventral valve strongly convex in lateral profile with maximum height anterior to the umbo | Shell transversely oval to subcircular with maximum height at the umbo or beak | Shell subcircular to transversely oval, and ventral valve convex to low subconical | Shell subcircular to transversely oval, and ventral valve convex to low subconical |
| Ventral pseudointerarea | Catacline to gently procline and divided medially by shallow intertrough | Catacline to slightly apsacline, narrow and divided by deeper intertrough | Procline to catacline, poorly defined laterally and divided by intertrough | Catacline to procline, rarely apsacline and divided by shallow intertrough | Gently procline and with shallow to vestigial intertrough |
| Apical pit | Present | Present | Present | Present | Vestigial to absent |
| Apical process | Boss-like and anterior to the internal foramen | Boss-like and raised area surrounding the internal foramen | Boss-like and thickening anterior to internal foramen but not filled apex | High, boss-like, and anterior to the foramen | Vestigial to absent |
| Dorsal valve | Gently convex and with broad, shallow sulcus | Gently convex and with shallow sulcus | Weakly convex and absent sulcus | Gently convex and absent sulcus | Gently convex and absent sulcus |
| Dorsal pseudointerarea | Narrowly triangular and with orthocline to anacline | Low | Short | Vestigial and undivided | Narrowly triangular and with orthocline |
| Median groove | Shallow median groove | Lenticular median groove | Broadly triangular median groove | Broadly median groove | Shallow median groove |
| Median ridge | Low | Strong, low to subtriangular | Vestigial | High | Well-developed submedian ridge |
| Median buttress | Well-developed and posteriorly by a low median ridge | Well-developed and posteriorly discontinuous medially | Well developed | Low | Well developed |
| Reference | Rowell, [40] | Percival & Kruse, [9]; Popov et al. [43] and Holmer et al. [48] | Streng, [49] | Holmer & Popov, [3] and Duan et al., [7] | Li and Holmer, [8] |
| D | L | W | Lp | Wp | Ls | Wg | Lc | Wc |
| N | 8 | 8 | 18 | 15 | 10 | 16 | 12 | 9 |
| S | 0.3939 | 0.4323 | 0.0121 | 0.0977 | 0.2706 | 0.0425 | 0.0775 | 0.1368 |
| X | 0.9636 | 1.0285 | 0.0434 | 0.4275 | 0.7708 | 0.1792 | 0.2759 | 0.6180 |
| max | 1.8257 | 2.0183 | 0.0670 | 0.6610 | 1.2600 | 0.2900 | 0.4180 | 0.8570 |
| min | 0.5184 | 0.5494 | 0.0229 | 0.2767 | 0.4565 | 0.1401 | 0.1729 | 0.4314 |
| D | L/W | Wp/W | Lp/Wp | Wg/Wp | Lc/L | Wc/W | Ls/L | |
| N | 8 | 3 | 15 | 15 | 4 | 3 | 6 | |
| S | 4.16 | 4.94 | 1.87 | 7.43 | 3.44 | 2.95 | 4.66 | |
| X | 0.9433 | 0.3761 | 0.0995 | 0.4226 | 0.2211 | 0.4455 | 0.6862 | |
| max | 0.9787 | 0.4263 | 0.1249 | 0.5305 | 0.2696 | 0.4663 | 0.7560 | |
| min | 0.9046 | 0.3275 | 0.0638 | 0.2953 | 0.1961 | 0.4246 | 0.6200 | |
| V | L | W | Lf | Wf | L/W | Lf/L | Wf/W | Lf/Wf |
| N | 6 | 6 | 8 | 8 | 6 | 2 | 2 | 8 |
| S | 0.2974 | 0.3301 | 0.0149 | 0.0161 | 0.0552 | 0.0029 | 0.0113 | 0.1412 |
| X | 0.4663 | 0.5223 | 0.0526 | 0.0649 | 0.8945 | 0.0584 | 0.0726 | 0.8137 |
| max | 1.5843 | 1.7599 | 0.0879 | 0.1078 | 0.9929 | 0.0613 | 0.0839 | 1.0956 |
| min | 0.7284 | 0.8019 | 0.0372 | 0.0506 | 0.8058 | 0.0554 | 0.0612 | 0.5886 |
| Species | Formation | Region | Ages | Biostratigraphy | Sedimentary Environment | Reference |
|---|---|---|---|---|---|---|
| Hadrotreta Primaeva Primaeva Hadrotreta Primaeva minor | Pioche shale | Nevada, USA | Age 4 to Wuliuan | Late Bonnia–Olenellus Zone and Pre-Albertella Zone | Sea margin of the Carbonate platform | Rowell, [18,40] |
| Hadrotreta primaea | Cadiz Fm. | California, USA | Wuliuan | Olenellus multinodus Subbiozone of the Bonnia–Olenellus Zone | Open shallow subtidal | Liang et al. [28] |
| Hadrotreta taconica | Forteau Fm. | Canada | Age 4 | Bonnia–Olenellus Zone | The inner shelf is in shallow water and the distal shelf setting in deep water | Skovsted et al. [25] |
| Hadrotreta primaeva | Coonigan Fm. | New South Wales, Australia | Wuliuan | Occurrence of Pagetia | Edge of a shallow shelf | Roberts and Jell, [20] |
| Hadrotreta timchristiorum | Parahio Fm. | Himalayas, India | Wuliuan | Orycticephalus salteri–Paramecephalus–Defossu Zone | Seaward-facing shelf setting | Popov et al. [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, B.; Wang, Y.; Yang, X.; Wu, W. The First Report of the Acrotretoid Brachiopod Hadrotreta from the Tsinghsutung Formation Cambrian (Series 2, Stage 4), Guizhou, South China. Biology 2023, 12, 1083. https://doi.org/10.3390/biology12081083
Wei B, Wang Y, Yang X, Wu W. The First Report of the Acrotretoid Brachiopod Hadrotreta from the Tsinghsutung Formation Cambrian (Series 2, Stage 4), Guizhou, South China. Biology. 2023; 12(8):1083. https://doi.org/10.3390/biology12081083
Chicago/Turabian StyleWei, Buqing, Yuan Wang, Xinglian Yang, and Weiyi Wu. 2023. "The First Report of the Acrotretoid Brachiopod Hadrotreta from the Tsinghsutung Formation Cambrian (Series 2, Stage 4), Guizhou, South China" Biology 12, no. 8: 1083. https://doi.org/10.3390/biology12081083