
Konstantinovia Is Not Monotypic, and a New Attempt to Determine Relationships in Cephaloziellaceae–Scapaniaceae Superclade (Marchantiophyta)
 , , , , and
, , , , and
Abstract
:1. Introduction
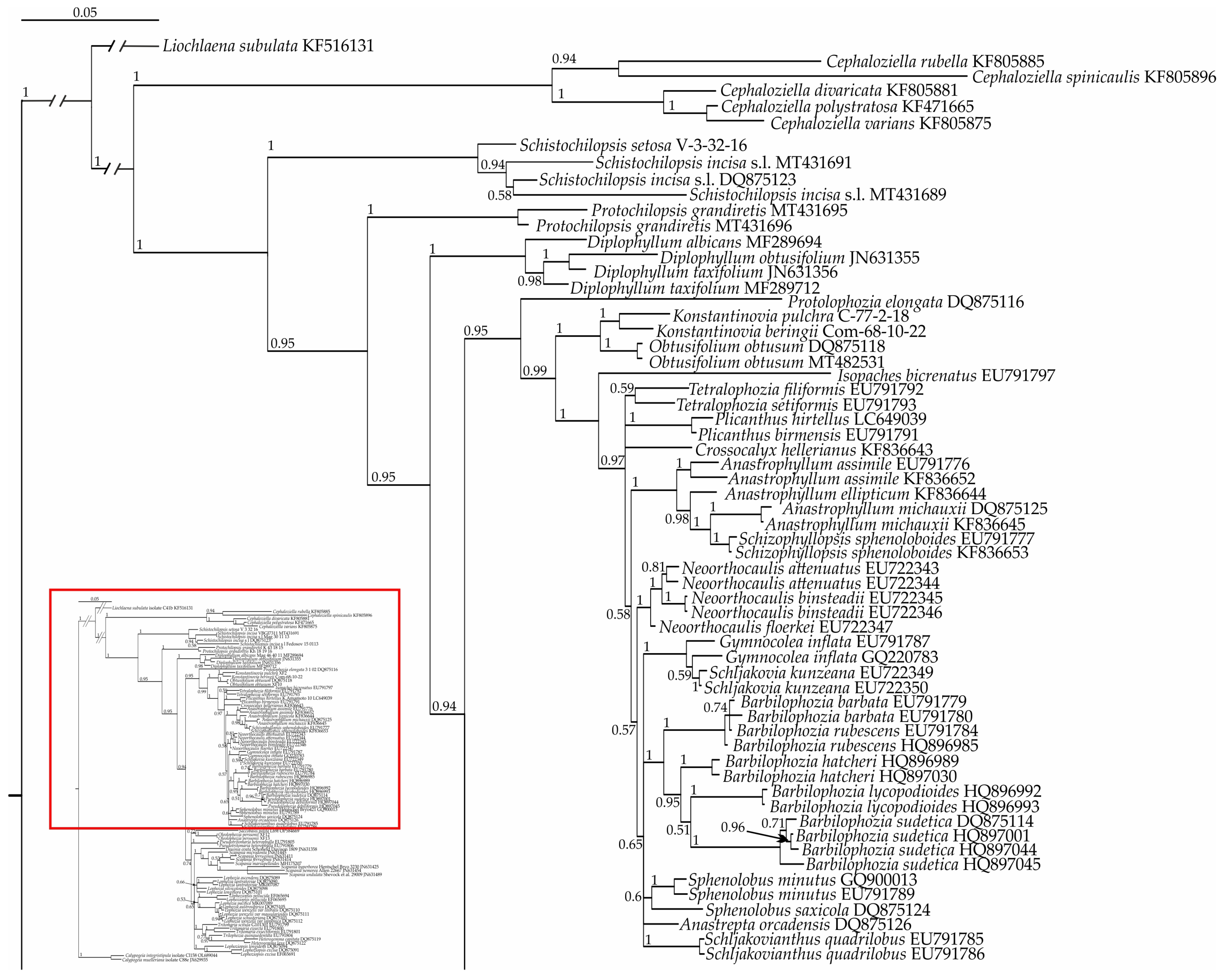
2. Results
3. Discussion
3.1. On the Circumscription of the Genus Konstantinovia
3.1.1. Morphology
3.1.2. Geographic and Ecological Considerations
3.1.3. On the Phylogenetic Relationships in the Scapaniaceae–Cephaloziellaceae Superclade
3.2. Taxonomy
3.2.1. Konstantinovia beringii Bakalin, Fedosov, Klimova et Maltseva sp. nov.
3.2.2. Oleolophoziaceae Bakalin et Fedosov fam. nov.
4. Materials and Methods
4.1. Taxon Sampling
4.2. DNA Isolation, Amplification, and Sequencing
4.3. Phylogenetic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schuster, R.M. The Hepaticae and Anthocerotae of North America East of the Hundredth Meridian. Vol. II; Columbia University Press: New York, NY, USA, 1969; pp. 1–1062. [Google Scholar]
- Schljakov, R.N. Hepaticae of the North of the USSR. 3. Lophoziaceae, Mesoptychiaceae; Nauka: Leningrad, Russia, 1980; pp. 1–188. [Google Scholar]
- De Roo, R.T.; Hedderson, T.A.; Söderström, L. Molecular insights into the phylogeny of the leafy liverwort family Lophoziaceae cavers. Taxon 2007, 56, 301–314. [Google Scholar] [CrossRef]
- Söderström, L.; De Roo, R.; Hedderson, T. Taxonomic novelties resulting from recent reclassification of the Lophoziaceae/Scapaniaceae clade. Phytotaxa 2010, 3, 47–53. [Google Scholar] [CrossRef]
- Vilnet, A.A.; Konstantinova, N.A.; Troitsky, A.V. Molecular phylogeny and systematics of the suborder cephaloziineae with special attention to the family Cephaloziaceaes.l. (Jungermanniales, Marchantiophyta). Arctoa 2012, 21, 113–132. [Google Scholar] [CrossRef]
- Váňa, J.; Söderström, L.; Hagborg, A.; Von Konrat, M. Notes on early land plants today. 40. Notes on Cephaloziellaceae (Marchantiophyta). Phytotaxa 2013, 112, 1–6. [Google Scholar] [CrossRef]
- Patzak, S.D.F.; Schäfer-Verwimp, A.; Váňa, J.; Renner, M.A.M.; Peralta, D.F.; Heinrichs, J. Chonecoleaceae (Lophocoleineae) is a synonym of Cephaloziellaceae (Cephaloziineae) and Rivulariella (Jungermanniineae) belongs to Scapaniaceae s.l. (Cephaloziineae). Phytotaxa 2016, 267, 91–102. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Fedosov, V.E.; Fedorova, A.V.; Ma, W.Z. Obtusifoliaceae, a new family of leafy liverworts to accommodate Konstantinovia, newly described from the Hengduan Mts. (South China) and Obtusifolium (Cephaloziineae, Marchantiophyta). Plant Syst. Evol. 2021, 307, 62. [Google Scholar] [CrossRef]
- Vilnet, A.A.; Konstantinova, N.A.; Troitsky, A.V. Molecular insight on phylogeny and systematics of the Lophoziaceae, Scapaniaceae, Gymnomitriaceae and Jungermanniaceae. Arctoa 2010, 19, 31–50. [Google Scholar] [CrossRef]
- Shaw, B.; Crandall-Stotler, B.; Váňa, J.; Stotler, R.E.; von Konrat, M.; Engel, J.J.; Davis, C.E.; Long, D.G.; Sova, P.; Shaw, A.J. Phylogenetic relationships and morphological evolution in a major clade of leafy liverworts (phylum Marchantiophyta, order Jungermanniales): Suborder Jungermanniineae. Syst. Bot. 2015, 40, 27–45. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Fedosov, V.E.; Maltseva, Y.D.; Milyutina, I.A.; Klimova, K.G.; Nguyen, H.M.; Troitsky, A.V. Overview of Schistochilopsis (Hepaticae) in Pacific Asia with the description Protochilopsis gen. nov. Plants 2020, 9, 850. [Google Scholar] [CrossRef]
- Hulten, E. Flora of Kamchatka and the Adjacent Islands; Almquist und WiksellsBoktryckert-A.-B: Stokholm, Sweden, 1927; Volume 1, pp. 1–346. [Google Scholar]
- Yurtsev, B.A. Problems of Phytogeography in Northeastern Asia; Nauka: Leningrad, Russia, 1974; pp. 1–160. [Google Scholar]
- Tatewaki, M. Hultenia. J. Fac. Agric. Hokkaido Univ. 1963, 53, 131–199. [Google Scholar]
- Tatewaki, M. Phytogeography of the Islands of the North Pacific Ocean. In Pacific Basin Biogeography; Proceedings of the A Symposium Tenth Pacific Science Congress, Honolulu, HI, USA, 21 August–6 September 1961; Bishop Museum Press: Honolulu, HI, USA, 1963; pp. 23–28. [Google Scholar]
- Bakalin, V.A.; Cherdantseva, V.Y. Bryophyte flora of Mednyj Island and bryogeographyoa Aleutians (North Pacific). In Conservation of Biodiversity of Kamchatka and Coastal Waters, Proceedings of the Proceedings of VIII International Scientific, Petropavlovsk-Kamchatsky, Russia, 27–28 November 2007; Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2008; pp. 36–56. [Google Scholar]
- Fedosov, V.E.; Ignatova, E.A.; Ignatov, M.S.; Maksimov, A.I.; Zolotov, V.I. Moss flora of Bering Island (Commander Islands, North Pacific). Arctoa 2012, 21, 133–164. [Google Scholar] [CrossRef]
- Bakalin, V.A. The liverwort flora of Bering Island (North-West Pacific, Russia). Lindbergia 2005, 30, 79–92. [Google Scholar]
- Davison, P.G. Floristic and Phytogeographic Studies of the Hepatic Flora of the Aleutian Islands, Alaska. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, 1993. [Google Scholar]
- Schofield, W.B. Bryophyte disjunction in Northern Hemisphere: Europe and North America. Bot. J. Linnean Soc. 1988, 98, 211–224. [Google Scholar] [CrossRef]
- Davis, P.H. Cliff Vegetation in the Eastern Mediterranean. J. Ecol. 1951, 39, 63–93. [Google Scholar] [CrossRef]
- Natcheva, R.; Cronberg, N. What do we know about hybridization among bryophytes in nature? Can. J. Bot. 2004, 82, 1687–1704. [Google Scholar] [CrossRef]
- Bakalin, V.; Fedosov, V.; Long, D.G.; Fedorova, A.; Maltseva, Y. Protoharpanthus gen. nov. (Harpanthaceae)—A relict relative of Harpanthus from the Sino-Himalaya. Bryologist 2021, 124, 218–229. [Google Scholar] [CrossRef]
- Buch, H. Lophozia perssonii Buch et S. Arnell, spec. nov. Bot. Not. 1944, 97, 381–387. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Worldclim, 2020–2022. Available online: https://www.worldclim.org/ (accessed on 24 February 2020).
- Groth, H.; Helms, G.; Heinrichs, J. The systematic status of Plagiochila Sects. Bidentes Carl and Caducilobae Inoue (Hepaticae) inferred from NrDNA ITS sequences. Taxon 2002, 51, 675–684. [Google Scholar] [CrossRef]
- Milyutina, I.A.; Goryunov, D.V.; Ignatov, M.S.; Ignatova, E.A.; Troitsky, A.V. The phylogeny of Schistidium (Bryophyta, Grimmiaceae) based on the primary and secondary structure of nuclear RDNA internal transcribed spacers. Mol. Biol. 2010, 44, 883–897. [Google Scholar] [CrossRef]
- Pacak, A.; Szweykowska-Kulińska, Z. Molecular data concerning alloploid character and the origin of chloroplast and mitochondrial genomes in the liverwort Pellia borealis. Plant Biotechnol. J. 2000, 2, 101–108. [Google Scholar]
- Fedosov, V.E.; Fedorova, A.V.; Ignatova, E.A.; Bobrova, V.K.; Troitsky, A.V. RPS4 and NAD5 sequences indicate the polyphyly of ditrichaceae and parallelisms in the evolution of haplolepidous mosses. Mol. Biol. 2015, 49, 993–997. [Google Scholar] [CrossRef]
- Fedosov, V.E.; Fedorova, A.V.; Fedosov, A.E.; Ignatov, M.S. On the taxonomic position of Pseudoditrichaceae (Bryophyta). Bot. J. Linn. Soc. 2016, 181, 139–155. [Google Scholar] [CrossRef]
- Cox, C.J.; Goffinet, B.; Newton, A.E.; Shaw, A.J.; Hedderson, T.A.J. Phylogenetic relationships among the diplolepideous-alternate mosses (bryidae) inferred from nuclear and chloroplast DNA sequences. Bryologist 2000, 103, 224–241. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Maltseva, Y.D.; Vilnet, A.A.; Choi, S.S. The transfer of Tritomaria koreana to Lophozia has led to recircumscription of the genus and shown convergence in Lophoziaceae (Hepaticae). Phytotaxa 2021, 512, 041–056. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Simmons, M.P.; Ochoterena, H. Gaps as Characters in Sequence-Based Phylogenetic Analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef]
- Müller, K. SeqState—Primer design and sequence statistics for phylogenetic DNA data sets. Appl. Bioinform. 2005, 4, 65–69. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree v.1.4.4. Computer Program and Documentation. Distributed by the Author. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 20 November 2023).
- Bechteler, J.; Peñaloza-Bojacá, G.; Bell, D.; Burleigh, J.G.; McDaniel, S.F.; Christine Davis, E.; Sessa, E.B.; Bippus, A.; Christine Cargill, D.; Chantanoarrapint, S.; et al. Comprehensive phylogenomic time tree of bryophytes reveals deep relationships and uncovers gene incongruences in the last 500 million years of diversification. Am. J. Bot. 2023. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Konstantinovia beringii | Konstantinovia pulchra |
|---|---|---|
| Ventral branches | Not present (at least unknown) | Sparse |
| Tooth near antical leaf base | Rare | Common |
| Tooth near postical leaf base | Absent | Sparse |
| Underleaves in sterile shoots | Absent | Present, nearly regular |
| Gemmae | Absent (unknown) | Sporadically present |
| Bioclimates | Data for Konstantinovia pulchra Type Locality | Data for Konstantinovia beringii Type Locality |
|---|---|---|
| BIO1 = Annual Mean Temperature, °C | 5.0625 | 1.76667 |
| BIO4 = Temperature Seasonality (standard deviation × 100) | 548.76358 | 617.13169 |
| BIO5 = Max Temperature of Warmest Month, °C | 17.6 | 13.4 |
| BIO6 = Min Temperature of Coldest Month, °C | −7.4 | −8.6 |
| BIO12 = Annual Precipitation, mm/year | 859.00 | 916.00 |
| BIO15 = Precipitation Seasonality (Coefficient of variation) | 64.11567 | 26.12135 |
| BIO16 = Precipitation of Wettest Quarter, mm/quarter | 396.00 | 305.00 |
| BIO17 = Precipitation of Driest Quarter, mm/quarter | 51.00 | 166.00 |
| Locus | Sequence (5′-3′) | Direction | Reference |
|---|---|---|---|
| ITS 1–2 nrDNA | CGGTTCGCCGCCGGTGACG | forward | [27] |
| ITS 1–2 nrDNA | GATATGCTTAAACTCAGCGG | reverse | [28] |
| trnG cpDNA | ACCCGCATCGTTAGCTTG | forward | [29] |
| trnG cpDNA | GCGGGTATAGTTTAGTGG | reverse | [29] |
| rps4 cpDNA | TACCGAGGGTTCGAATCCCT | forward | this study |
| rps4 cpDNA | ATGTCCCGTTATCGAGGACCT | reverse | [30] |
| rbcL cpDNA | ATGTCACCACAAACGGA | forward | [31] |
| rbcL cpDNA | GTATCTATTGTTTCATATTC | reverse | this study |
| psbA cpDNA | GACGAGTTCCGGGTTCGA | forward | [32] |
| psbA cpDNA | TGGAATGGGTGCATAAGG | reverse | [32] |
| trnL–F cpDNA | CGAAATTGGTAGACGCTGCG | forward | [33] |
| trnL–F cpDNA | TGCCAGAAACCAGATTTGAAC | reverse | [33] |
| Initial denaturation | 3 min–94 °C | |
| Denaturation | 30 s–95 °C | 35 cycles |
| Annealing | 20 s (trnL–F), 30 s (trnG, ITS 1–2), 1 min (rbcL, rps4, psbA) | |
| at 50 °C (rbcL), 56 °C (trnG, psbA), 58 °C (trnL–F), 60 °C (ITS 1–2), 62 °C (rps4) | ||
| Elongation | 30 s–72 °C (ITS 1–2, trnL–F, trnG), 1 min (rbcL, rps4, psbA) | |
| Final elongation | 3 min–72 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakalin, V.A.; Fedosov, V.E.; Klimova, K.G.; Maltseva, Y.D.; Fedorova, A.V.; Choi, S.S. Konstantinovia Is Not Monotypic, and a New Attempt to Determine Relationships in Cephaloziellaceae–Scapaniaceae Superclade (Marchantiophyta). Plants 2024, 13, 15. https://doi.org/10.3390/plants13010015
Bakalin VA, Fedosov VE, Klimova KG, Maltseva YD, Fedorova AV, Choi SS. Konstantinovia Is Not Monotypic, and a New Attempt to Determine Relationships in Cephaloziellaceae–Scapaniaceae Superclade (Marchantiophyta). Plants. 2024; 13(1):15. https://doi.org/10.3390/plants13010015
Chicago/Turabian StyleBakalin, Vadim A., Vladimir E. Fedosov, Ksenia G. Klimova, Yulia D. Maltseva, Alina V. Fedorova, and Seung Se Choi. 2024. "Konstantinovia Is Not Monotypic, and a New Attempt to Determine Relationships in Cephaloziellaceae–Scapaniaceae Superclade (Marchantiophyta)" Plants 13, no. 1: 15. https://doi.org/10.3390/plants13010015