
Shoot Growth and Flower Bud Production of Peony Plants under Subtropical Conditions


1
Genecology Research Centre, University of the Sunshine Coast, Maroochydore, QLD 4558, Australia
2
Centre for Planetary Health and Food Security, School of Environment and Science, Griffith University, Nathan, QLD 4111, Australia
*
Author to whom correspondence should be addressed.
Horticulturae 2021, 7(11), 476; https://doi.org/10.3390/horticulturae7110476
Submission received: 15 September 2021
/
Revised: 1 November 2021
/
Accepted: 3 November 2021
/
Published: 8 November 2021
(This article belongs to the Special Issue Peony as the Newly-Emerged Queen of the Field-Grown Ornamentals)
Abstract
:Peony plants require temperate winter temperatures to break underground bud dormancy and allow shoot emergence and flowering in spring. This study assessed whether artificial chilling at 4 °C for 2–6 weeks could induce shoot emergence and flowering under subtropical conditions. It also assessed whether pre-treatment at cool temperatures prior to chilling, or gibberellin application after chilling, promoted shoot emergence and flowering. Artificial chilling at 4 °C for 4 or 6 weeks promoted the greatest shoot emergence. Pre-treatment at cool temperatures did not affect shoot growth or flower bud production but it improved shoot emergence from plants also treated with gibberellin. Gibberellin more than doubled the number of shoots per plant without affecting shoot length. The optimal treatment combination for shoot emergence, growth and flower bud production was pre-treatment from 20 °C to 8 °C over an 8-day period in autumn, chilling at 4 °C for 6 weeks in early winter, and treatment with 250 mL of 100 mg/L GA3, before returning plants to subtropical winter conditions. This treatment combination provided medians of 3 (0–7) and 8 (0–31) flower buds per plant in the second and third years of production, respectively. Peony flowers can be produced in subtropical climates using artificial chilling and gibberellin, allowing out-of-season market supply.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Herbaceous peonies (Paeonia lactiflora and related species) are highly valued for their attractive aromatic flowers [1]. Flower development in herbaceous peonies is initiated in temperate climates in late summer from renewal buds in their perennial underground crown [2]. The renewal buds continue to develop underground until the plant enters dormancy in autumn [3,4]. Peonies require exposure to a prolonged period of natural cold temperature or artificial chilling to break bud dormancy, elongate and flower in the following spring [2,5,6]. Dormancy has been considered to only be broken in temperate regions that experience freezing temperatures for 2–3 months each winter [7]. However, peony dormancy can be broken in parts of Israel, Italy and southern France that only occasionally experience freezing temperatures [8]. Dormancy of herbaceous peonies has also been broken in parts of New Zealand, Chile and Argentina that experience mild winters [5,7].
Peonies are supposedly unable to flower in the subtropics because the winter is too warm to break floral bud dormancy. Forced flowering has been accomplished in mild temperate climates using techniques that alter the environment of peony plants to break their bud dormancy [1,8]. Environmental controls have included altered temperatures and photoperiods, although low temperature appears to be the primary determinant in forcing the flowering of peony plants [3]. Artificial chilling at 0–6 °C for 4–6 weeks is typically required to break dormancy of the underground renewal buds of peony plants. Subsequent growth and blooming of floral shoots usually require temperatures of 15–25 °C, with higher temperatures causing flower bud abortion [8]. A cooling pre-treatment prior to chilling may also be needed to alleviate the physiological stress imposed on plants when there is a large temperature gradient between the ambient growing conditions and chilling treatment [9].
Peony bud release can also be stimulated and shoot growth can be enhanced by application of gibberellic acid (GA3) [4,10]. Application of 100 mg/L GA3 post-chilling to the potting mix around the underground crown of peony plants promotes shoot emergence and growth and increases the number of flowers [6,8,10,11]. A lower GA3 concentration, 50 mg/L, provides lower shoot numbers, whereas a higher concentration, 250 mg/L, causes flower bud abortion [11].
The objective of this study was to determine the extent to which artificial chilling at 4 °C could induce shoot emergence, shoot growth and flowering of peony plants under subtropical conditions. The study also assessed whether pre-treating the plants at low temperature prior to chilling, or applying GA3 after chilling, could enhance shoot emergence, shoot growth and flower bud production. The results have the potential to promote out-of-season production of peony flowers during winter in subtropical regions.
2. Materials and Methods
2.1. Plant Material
A total of 110 peony plants was obtained from Mistydowns Nursery, Springmount (37°24′ S, 143°56′ E) and Pinerise Nursery, Highcroft (43°14′ S, 147°76′ E), Australia, in late summer and autumn 2016. The number of plants available for experimentation was constrained by supply and the cultivar of each plant was not always identified. However, the approximate numbers of plants for each cultivar were 20, 20, 18, 17, 13, 12 and 10 for ‘Duchesse de Nemours’, ‘Lady Bird’, ‘Sarah Bernhardt’, ‘Mossi’, ‘Monsieur Jules Elie’, ‘Sylvia’ and ‘Pecher’, respectively. The plants were placed immediately in a glasshouse with additional 50%-shade cloth at the University of the Sunshine Coast (26°43′ S, 153°03′ E). The plants received overhead watering for 3 min at each of 07:00 h, 10:00 h and 13:00 h each day. They were transplanted into 14-L pots containing a 75/25 (v/v) mixture of shredded pine bark and perlite with 3 kg of 8–9 month slow-release Osmocote™ fertilizer (Scotts International, Heerlen, The Netherlands), 3 kg lime (Unimin, Lilydale, Australia), 1 kg Micromax™ micronutrients (Scotts Australia, Baulkham Hills, Australia), 1 kg Hydroflo™ wetting agent (Scotts Australia, Baulkham Hills, Australia), and 4 kg of gypsum incorporated per m3 to obtain a potting mix pH of approximately 7.0. The plants were then placed randomly in a translucent-white polyethylene tunnel house (Figure 1A). The plants received drip irrigation for 3–5 min each day at 09:30 h and received a 3–5-min period of drip irrigation at 14:30 h, as necessary, depending on the season. Temperatures in the glasshouse and tunnel house were recorded using Tinytag™ dataloggers (Hastings Data Loggers, Port Macquarie, Australia) (Figure 2). Shoots senesced naturally in autumn and any remaining shoots on each plant were pruned at the level of the potting mix. All plants were fertilised with 250 mL of 2 g/L Thrive™ soluble fertiliser (25.0% N, 5.0% P, 8.8% K, w/w) (Yates, Auckland, New Zealand) every 11 d during later periods of shoot growth.
2.2. Year 1: Chilling Duration
Plants were allocated randomly into four treatments that involved chilling the plants for (a) 0, (b) 2, (c) 4 or (d) 6 weeks at 4 °C in darkness in a cold room. Chilling for 2, 4 and 6 weeks occurred from 29 June–13 July, 22 June–20 July and 15 June–27 July 2016, respectively. Plants were returned to ambient conditions in the tunnel house immediately after chilling. Control plants (i.e., 0 weeks of chilling) remained in the tunnel house for the entire experimental period. The number of replicate plants per treatment was 19–34.
2.3. Year 2: Cooling Pre-Treatment and Gibberellin Application
Plants were reallocated randomly in Year 2 into four treatment combinations: (a) chilling only; (b) pre-treatment at cool temperatures followed by chilling; (c) chilling followed by GA3 application; or (d) pre-treatment at cool temperatures followed by chilling and GA3 application. Plants that were pre-treated at cool temperatures were transferred to a cold room where they were progressively cooled at 20 °C for 2 days, 16 °C for 2 days, 12 °C for 2 days, and then 8 °C for 2 days in darkness from 10 May–18 May 2017. All plants from all treatment combinations were then chilled at 4 °C for 6 weeks in darkness in a cold room from 18 May–30 June 2017. All plants were then returned to the tunnel house. Plants that received the GA3 treatment were provided with 250 mL of 100 mg/L GA3 on 30 June 2017, with most of the solution applied towards the centre of the pot. All other plants were treated with 250 mL of water. The number of replicate plants per treatment combination was 18–32.
2.4. Year 3: Commercial Proof-of-Concept
All emergent plants from Year 2 received the full combination of pre-treatment at cool temperatures, chilling, and GA3 application in Year 3. A total of 65 plants was transferred to a cold room where the plants were progressively cooled at 20 °C for 2 days, 16 °C for 2 days, 12 °C for 2 days, and then 8 °C for 2 days in darkness from 9 May–17 May 2018. All plants were then chilled at 4 °C for 6 weeks in darkness in a cold room from 17 May–28 June 2018. The plants were then returned to the tunnel house and treated with 250 mL of 100 mg/L GA3 on 28 June 2018.
2.5. Shoot Emergence and Flower Bud Production
The number of emerging shoots (Figure 1B), the length of each emerging shoot (Figure 1C), and the numbers of flower buds (Figure 1D) and open flowers on each emerged plant were counted or measured weekly. Measurements commenced immediately after chilling and continued until the end of the flowering season in late spring. The total number of leaves on each plant (Figure 1D) was recorded at the end of each experiment. The proportion of plants that emerged, the mean time to first shoot emergence on sprouted plants, the average length of the shoots on each emerged plant, and the total length of shoots on each emerged plant were then calculated for each treatment.
2.6. Statistical Analyses
Data for the proportion of plants that emerged were analysed by sequential binomial tests with significance test values adjusted using Holm–Šidák corrections. Data for all cultivars were pooled due to the limited number of plants available for each cultivar. Data for time to shoot emergence, shoot number, shoot length, leaf number, and flower bud number were analysed by Kruskal–Wallis test. Only plants that had emerged stems were included in these analyses. A post-hoc Dunn’s test with Bonferroni corrections was performed when significant differences among the medians were detected by Kruskal–Wallis test. Differences between means or medians were regarded as significant at p < 0.05. Medians for time to shoot emergence, shoot number, shoot length, leaf number, and flower bud number are presented with ranges.
3. Results
3.1. Shoot Emergence
More plants that received 4 or 6 weeks of chilling had emergent shoots in Year 1 than did plants that received 0 or 2 weeks of chilling (Figure 3A). All plants that received the full combination of pre-treatment, chilling and GA3 in Year 2 had emergent shoots, compared with 82–91% of plants having emergent shoots following other treatments (Figure 3B). All plants were pre-treated at cool temperatures, chilled and treated with GA3 in Year 3, and 94% of these plants had emergent shoots (Figure 3C).
The shoots of plants that received 6 weeks of chilling in Year 1 had earlier emergence than the shoots of plants that received 2 weeks of chilling, but differences between other chilling durations were not significant (Figure 3D). Shoot emergence from most plants in Years 2 and 3 commenced within 1 week of chilling (Figure 3E,F).
Chilling duration did not significantly affect the number of emerged shoots per plant in Year 1 (Figure 3G). Application of GA3 in Year 2 more than doubled the number of emerged shoots per plant (Figure 3H). The plants produced 10 (2–31) shoots after pre-treatment, chilling, and GA3 treatment in Year 3 (Figure 3I).
3.2. Shoot Growth
Plants that received 4 or 6 weeks of chilling in Year 1 had longer shoots than unchilled plants (Figure 4A). Pre-treatment at cool temperatures and GA3 treatments in Year 2 did not significantly affect individual shoot length (Figure 4B). Individual shoot length was 26 (11–51) cm in Year 3 (Figure 4C).
Plants that received 2, 4 or 6 weeks of chilling in Year 1 had greater total shoot lengths than unchilled plants (Figure 4D). Plants that received the full treatment combination of pre-treatment at cool temperatures, chilling and GA3 in Year 2 had greater total shoot length than plants that were chilled only or pre-treated and chilled (Figure 4E). Plants had a total shoot length of 255 (21–942) cm in Year 3 (Figure 4F).
Chilling in Year 1 increased the number of leaves per plant (Figure 4G). Plants that received the full treatment combination of pre-treatment at cool temperatures, chilling and GA3 in Year 2 had approximately twice the number of leaves of plants that received chilling only or pre-treatment followed by chilling (Figure 4H). Plants had 174 (27–488) leaves in Year 3 (Figure 4I).
3.3. Flower Bud Production
Chilling duration did not significantly affect the number of flower buds per plant in Year 1 (Figure 5A). Most flower buds in Year 1 aborted, leaving only four buds that reached full bloom. Three flowers were on plants that had been chilled for 6 weeks and the other was on a plant that had been chilled for 4 weeks. Plants that received the full treatment combination of pre-treatment at cool temperatures, chilling and GA3 in Year 2 had more flower buds than plants that were chilled only and plants that were pre-treated and chilled (Figure 5B). Most flower buds aborted during a hot period in early spring of Year 2 (Figure 2B), leaving 25 buds that reached full bloom. Eight flowers were on plants that had been chilled only; five were on plants that were pre-treated and chilled; three were on plants that received chilling and GA3; and nine were on plants that received pre-treatment, chilling and GA3. Peony plants produced 8 (0–31) flower buds in Year 3 (Figure 5C). A total of 171 flower buds reached full bloom in Year 3 (Figure 6).
4. Discussion
This study assessed whether artificial chilling could enable peony shoots to emerge, elongate and flower under subtropical conditions. The study also determined the effect of a cool-temperature pre-treatment prior to chilling, and a gibberellin application after chilling, on shoot emergence, elongation and flowering. Artificial chilling for 2, 4 or 6 weeks at 4 °C promoted shoot emergence and shoot growth from most peony plants. Pretreating the plants prior to chilling did not significantly affect shoot growth or flower bud production but pre-treatment improved shoot emergence from plants that were treated with gibberellin. Gibberellin application, either alone or in combination with pre-treatment, more than doubled the number of shoots per emergent plant. The optimal treatment combination for inducing shoot emergence, elongation and flower bud production was pre-treatment at cool temperatures, chilling and gibberellin application. This treatment combination allowed mass production of high-quality peony flowers during late winter and early spring under subtropical conditions.
Artificial chilling for 2, 4 or 6 weeks at 4 °C stimulated shoot emergence from most peony plants, although more plants displayed emergence when they experienced 4 or 6 weeks, compared with 2 weeks of chilling. Chilling for 4–6 weeks at 0–4 °C has been used to break bud dormancy of peonies in temperate parts of Japan and China [10,12] and chilling for 4 weeks at 5–6 °C has been sufficient to break dormancy in temperate northern California [3]. Chilling at greater than 6 °C is less effective, while chilling below 2 °C increases the number of emerging stems but can also cause some flower buds to abort, as found in Israel [1,2,6,8].
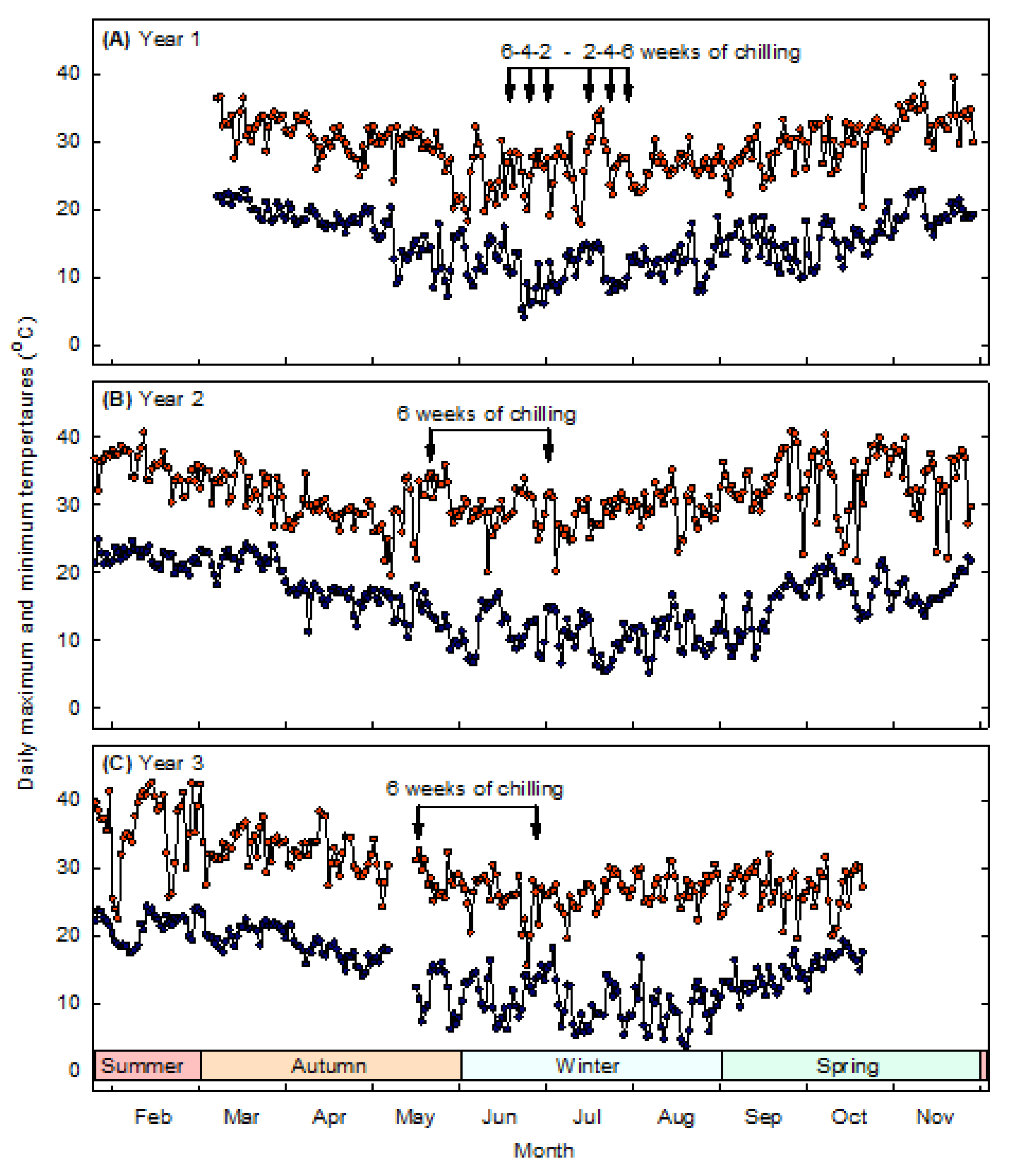
The current study was conducted under subtropical conditions where most peony plants received negligible natural chilling prior to artificial chilling. The plants were transferred from ambient temperatures of around 25 °C/10 °C (daily maximum/minimum) to 4 °C in the cold room and then returned to ambient temperatures of around 26 °C/12 °C. The threshold chilling requirement for peony plants may thus be much less than previously thought, with 2–6 weeks of artificial chilling at 4 °C being sufficient to promote shoot emergence and growth from plants that otherwise experienced warm ambient conditions. Sufficient chilling of peony plants mobilises stored carbohydrate from underground roots and transports soluble sugars to the renewal bud meristems [6,13]. Chilling the peony plants for 2–6 weeks at 4 °C may have been sufficient to mobilise sugars, allowing shoots to emerge from most plants when transferred to ambient temperatures of 26 °C/12 °C. Chilling peonies for 3–4 weeks at 0–4 °C also increases synthesis of gibberellins, which play a central role in releasing dormancy [14]. Chilling peonies for 3–5 weeks at 0–4 °C, prior to GA3 application, promotes dormancy release and increases stem height, leaf size and number of flowers [10]. However, the number of flower buds produced per plant after 6 weeks of chilling at 4 °C was low in the current study. The chilling treatment was, therefore, supplemented with cool-temperature pre-treatment and gibberellin treatment in the second year of the study in an attempt to alleviate chilling stress and enhance shoot emergence, shoot growth and flower bud production.
Pre-treating the peony plants from 20 °C to 8 °C over an 8-day period, prior to chilling at 4 °C, did not significantly affect shoot growth or flower bud production. However, pretreatment increased, from 91% to 100%, the percentage of gibberellin-treated plants that had emerged shoots. Pre-treatment at cool temperatures appears to alleviate plant stress during transfer of peony plants from average late-summer temperatures of 27 °C/19 °C in Japan to chilling at 4 °C [15]. Pre-treatment may, alternatively, have increased the number of effective chill-hours experienced by peony plants in the current study, although pretreatment only increased the percentage of plants with emerged shoots when it was coupled with subsequent GA3 application. These results suggest that shoot emergence is slightly susceptible to artificial-chilling injury in the subtropical autumn, but that subsequent growth of the emerged shoots in winter and spring is not affected by the sudden transfer from warm to cold conditions in the autumn. Shoot emergence and flowering have been shown to be accelerated when peony plants were pre-treated at 15 °C for 10 d in late summer in Japan prior to chilling at 4 °C [15]. Flower bud abortion decreases from 88% to 16% when peony plants are pre-treated at 10 °C for 14 d in South Korea in summer, when ambient field temperatures are 29 °C/9 °C, prior to chilling at 0 °C [16].
Gibberellin application more than doubled the number of emerged shoots on each peony plant. Gibberellin, when applied in combination with the cool-temperature pre-treatment in year 2, also increased the percentage of plants with emerged shoots, the total length of shoots on each plant, the number of leaves on each plant, and the number of flower buds on each plant. Gibberellin is considered a key regulator of shoot elongation in plants [17], but the effects of GA3 on peony shoot emergence and growth occurred, surprisingly, without significant effects on the length of individual shoots. GA3 increases the numbers of emerged shoots and leaves on peony plants in Poland [18] and increases shoot length of peony plants in Israel [11]. GA3 application also increases shoot length and leaf growth of chilled peonies in China [10]. The lack of a gibberellin effect on individual shoot length in the current study could have been the result of an unusual spike in temperature to 32 °C/14 °C during mid-winter. Peony shoots are shortest in New Zealand when temperatures rise to 25 °C/15 °C [5] and are shortest in Israel when night temperatures exceed 10 °C [8].
Application of GA3 to chilled peony plants may have decreased the threshold level of chilling required for shoot emergence and flower bud formation, resulting in almost all pre-treated plants having emerged shoots and tripling the number of flower buds. GA3 application increases the number of flowers 20-fold, and reduces the time to flowering of peonies in China [10]. GA3 application more than doubles the number of flowers produced for some cultivars in Israel [11], and it doubles flower production and reduces the time to flowering in Korea [19]. GA3 has also been used to break the dormancy of non-chilled peonies in the USA [4] and can partially or completely replace the chilling requirement for breaking bud dormancy in Israel and South Korea [20,21]. Chilling initiates gibberellin synthesis and transport in peony plants, which can assist in releasing renewal buds from dormancy by promoting cell division and shoot elongation [13]. Chilling and gibberellin promote the expression of genes that regulate the transition from vegetative to floral development in the apical meristem of renewal buds [22,23].
Most of the flower buds produced in the second year of the experiments aborted due to unusually high late-winter temperatures. Twenty-five flowers reached full bloom in Year 2, whereas 171 flowers reached full bloom in Year 3. Ambient temperatures in the tunnel house reached as high as 40 °C/19 °C during late winter and early spring of Year 2 but only as high as 31 °C/15 °C during the same period in Year 3. Moderate temperatures of 22 °C/10 °C allow flower bud development during the initial period after chilling in Israel, while temperatures of 28 °C/22 °C at later stages of development cause flower bud abortion [8]. Preventing flower bud abortion during a forced winter bloom in subtropical conditions may depend upon preventing exposure to unseasonally high temperatures during flower bud development. Options for preventing flower bud abortion could include placing plants into a controlled-environment chamber until anthesis or raising plants at higher altitudes that are likely to experience lower temperatures. Another reason the flower buds may have aborted was the age of the plants. Flower bud abortion on 1-year-old peony plants in the USA has been considered the result of limited carbohydrate supply to the developing buds [4]. Perennial peonies typically require 3 or 4 years to develop a sufficient root system to produce large numbers of flowers [9,24]. Commercial harvest generally occurs in the third year of peony growth, with full production in the fourth and fifth years when enough vegetative growth has been established to produce large numbers of renewal buds, stems and flowers [6].
5. Conclusions
Peony flower production in the subtropics proved possible using an optimal treatment combination of cool-temperature pre-treatment, artificial chilling, and gibberellin application. Pre-treating plants from 20 °C to 8 °C over an 8-day period in autumn, chilling them at 4 °C for 6 weeks in early winter, and treating them with 250 mL of 100 mg/L GA3, before returning them to subtropical winter conditions, provided the highest percentage of plants with shoot emergence, while also promoting high numbers of shoots, leaves and flower buds per plant. This treatment combination provided a median of 3 (0–7) flower buds per plant in the second year of production and a median of 8 (0–31) flower buds per plant in the third year of production. These results provide the foundation for commercial production of peony flowers in the subtropics, allowing market supply that is out-of-season with peony flower production in temperate climates.
Author Contributions
Conceptualization, K.C.B. and S.J.T.; methodology, K.C.B. and S.J.T.; investigation, K.C.B.; formal analysis, K.C.B., H.M.W. and S.J.T.; data curation, K.C.B.; writing—original draft preparation, K.C.B.; writing—review and editing, S.J.T. and H.M.W.; supervision, S.J.T. and H.M.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
A special thanks to Ian Darby, Tracey McMahon, Allison Morrison, Darren Morrow, Tarran Richards, Codie Murphy and Michael Nielsen for assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kamenetski, R.; Barzilay, A.; Cohen, M. Herbaceous peony for cut flower production: Flowering physiologation techniques and cultivation techniques. Acta Hortic. 2007, 755, 121–125. [Google Scholar] [CrossRef]
- Yom Din, G.; Cohen, M.; Kamenetsky, R. Database for herbaceous peony cultivated in warm climate regions: Effects of temperature on plant dormancy and growth. J. Hortic. 2015, 2, 3. [Google Scholar] [CrossRef]
- Byrne, T.; Halevy, A.H. Forcing herbaceous peonies. J. Am. Soc. Hortic. Sci. 1986, 111, 379–383. [Google Scholar]
- Evans, M.R.; Anderson, N.O.; Wilkins, H.F. Temperature and GA3 effects on emergence and flowering of potted Paeonia lactiflora. HortScience 1990, 25, 923–924. [Google Scholar] [CrossRef]
- Hall, A.J.; Catley, J.L.; Walton, E.F. The effect of forcing temperatures on peony shoot and flower development. Sci. Hortic. 2007, 113, 188–195. [Google Scholar] [CrossRef]
- Kamenetsky, R.; Dole, J. Herbaceous peony (Paeonia): Genetics, physiology and cut flower production. Floric. Ornam. Biotechnol. 2012, 6, 62–77. [Google Scholar]
- Fulton, T.A.; Hall, A.J.; Catley, J.L. Chilling requirements of Paeonia cultivars. Sci. Hortic. 2001, 89, 237–248. [Google Scholar] [CrossRef]
- Kamenetsky, R.; Barzilay, A.; Erez, A.; Halevy, A.H. Temperature requirements for floral development of herbaceous peony cv. ‘Sarah Bernhardt’. Sci. Hortic. 2003, 97, 309–320. [Google Scholar] [CrossRef]
- Rhie, Y.H.; Jung, H.H.; Kim, K.S. Chilling requirements for breaking dormancy and flowering in Paeonia lactiflora ‘Taebaek’ and ‘Mulsurae’. Hortic. Environ. Biotechnol. 2012, 53, 277–282. [Google Scholar] [CrossRef]
- Cheng, F.; Zhong, Y.; Long, F.; Yu, X.; Kamenetsky, R. Chinese herbaceous peonies: Cultivar selection for forcing culture and effects of chilling and gibberellin (GA3) on plant development. Isr. J. Plant Sci. 2009, 57, 357–367. [Google Scholar] [CrossRef]
- Halevy, A.H.; Levi, M.; Cohen, M.; Naor, V. Evaluation of methods for flowering advancement of herbaceous peonies. HortScience 2002, 37, 885–889. [Google Scholar] [CrossRef] [Green Version]
- Aoki, N. Effects of chilling period on the growth and cut-flower quality of forced herbaceous peony. Bull. Fac. Agric. Shimane Univ. 1991, 25, 149–154. [Google Scholar]
- Zhang, J.; Yang, L.; Teixeira da Silva, J.; Wei, D.; Yu, X. Biochemical and physiological studies on dormancy release in buds of field-grown Paeonia lactiflora. Acta Hortic. 2017, 1171, 99–110. [Google Scholar] [CrossRef]
- Gai, S.; Zhang, Y.; Liu, C.; Zhang, Y.; Zheng, G. Transcript profiling of Paeonia ostii during artificial chilling induced dormancy release identifies activation of GA pathway and carbohydrate metabolism. PLoS ONE 2013, 8, e55297. [Google Scholar] [CrossRef] [Green Version]
- Aoki, N. Effect of pre-chilling on the growth and development of flower bud and the flowering of forced herbaceous peony (Paeonia lactiflora Pall.). Bull. Fac. Agric. Shimane Univ. 1993, 27, 73–78. [Google Scholar]
- Park, J.H.; Rhie, Y.H.; Lee, S.Y.; Kim, K.S. Pre-chilling promotes flowering of Paeonia lactiflora ‘Taebaek’ without flower bud abortion. Hortic. Environ. Biotechnol. 2015, 56, 1–8. [Google Scholar] [CrossRef]
- Hedden, P.; Sponsel, V. A century of gibberellin research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [Green Version]
- Gabryszewska, E. Effects of temperature on the growth and dormancy of tissue-cultured herbaceous peony shoots. Acta Hortic. 2004, 725, 471–475. [Google Scholar] [CrossRef]
- Yeo, S.M.; Rhie, H.Y.; Lee, S.Y.; Jung, H.H.; Kim, K.S. Dormancy release and flowering of Paeonia lactiflora “Taebaek” by natural cumulative chilling and GA3 treatment. Hortic. Environ. Biotechnol. 2012, 53, 263–270. [Google Scholar] [CrossRef]
- Halevy, A.H.; Barzilay, A.; Kamenetsky, R. Flowering advancement in herbaceous peony. Acta Hortic. 2005, 673, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Cheng, F.; Yu, S.; Wang, Y. Effects of gibberellins and root powder on the forcing culture of tree peony. J. Beijing For. Univ. 2006, 28, 84–87. [Google Scholar]
- Mutasa-Göttgens, E.; Hedden, P. Gibberellin as a factor in floral regulatory networks. J. Exp. Bot. 2009, 60, 1979–1989. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Beruto, M.; Xue, J.; Zhu, F.; Liu, C.; Yan, Y.; Zhang, X. Molecular cloning and potential function prediction of homologous SOC1 genes in tree peony. Plant Cell Rep. 2015, 34, 1459–1471. [Google Scholar] [CrossRef]
- Walton, E.F.; McLaren, G.F.; Boldingh, H.L. Seasonal patterns of starch and sugar accumulation in herbaceous peony (Paeonia lactiflora Pall.). J. Hortic. Sci. Biotechnol. 2007, 82, 365–370. [Google Scholar] [CrossRef]
Figure 1.
Peony plants (A) in a polyethylene tunnel house, (B) with emerging shoots, (C) with elongated stems, and (D) with fully-elongated leaves and flower buds.
Figure 1.
Peony plants (A) in a polyethylene tunnel house, (B) with emerging shoots, (C) with elongated stems, and (D) with fully-elongated leaves and flower buds.

Figure 2.
Daily maximum and minimum temperatures experienced by peony plants in the tunnel house in (A) Year 1, (B) Year 2, and (C) Year 3 of the study. Arrows indicate the start and finish of 2, 4 or 6 weeks of chilling at 4 °C in Year 1, and 6 weeks of chilling at 4 °C in Years 2 and 3.
Figure 2.
Daily maximum and minimum temperatures experienced by peony plants in the tunnel house in (A) Year 1, (B) Year 2, and (C) Year 3 of the study. Arrows indicate the start and finish of 2, 4 or 6 weeks of chilling at 4 °C in Year 1, and 6 weeks of chilling at 4 °C in Years 2 and 3.

Figure 3.
Percentage of peony plants with emerged shoots in (A) Year 1, (B) Year 2 and (C) Year 3; the time after chilling to first shoot emergence in (D) Year 1, (E) Year 2 and (F) Year 3; and the number of shoots per emerged plant in (G) Year 1, (H) Year 2 and (I) Year 3. ‘0Ch’, ‘2Ch’, ‘4Ch’ and ‘6Ch’ = 0, 2, 4 or 6 weeks of chilling at 4 °C, respectively; ‘PC’ = pre-treatment from 20 °C to 8 °C prior to chilling; ‘GA’ = GA3 treatment after chilling. Means are presented in (A–C); medians (D–I) are presented with 25th and 75th percentiles (boxes), 10th and 90th percentiles (whiskers), and outliers (circles). Means or medians in the same year with different letters are significantly different (Binomial tests with Holm–Šidák corrections, or Kruskal–Wallis and Dunn’s tests with Bonferroni corrections; p < 0.05, n = 18–65).
Figure 3.
Percentage of peony plants with emerged shoots in (A) Year 1, (B) Year 2 and (C) Year 3; the time after chilling to first shoot emergence in (D) Year 1, (E) Year 2 and (F) Year 3; and the number of shoots per emerged plant in (G) Year 1, (H) Year 2 and (I) Year 3. ‘0Ch’, ‘2Ch’, ‘4Ch’ and ‘6Ch’ = 0, 2, 4 or 6 weeks of chilling at 4 °C, respectively; ‘PC’ = pre-treatment from 20 °C to 8 °C prior to chilling; ‘GA’ = GA3 treatment after chilling. Means are presented in (A–C); medians (D–I) are presented with 25th and 75th percentiles (boxes), 10th and 90th percentiles (whiskers), and outliers (circles). Means or medians in the same year with different letters are significantly different (Binomial tests with Holm–Šidák corrections, or Kruskal–Wallis and Dunn’s tests with Bonferroni corrections; p < 0.05, n = 18–65).

Figure 4.
Length of individual shoots on peony plants in (A) Year 1, (B) Year 2 and (C) Year 3; the total length of shoots per plant in (D) Year 1, (E) Year 2 and (F) Year 3; and the number of leaves per plant in (G) Year 1, (H) Year 2 and (I) Year 3. ‘0Ch’, ‘2Ch’, ‘4Ch’ and ‘6Ch’ = 0, 2, 4 or 6 weeks of chilling at 4 °C, respectively; ‘PC’ = pre-treatment from 20 °C to 8 °C prior to chilling; ‘GA’ = GA3 treatment after chilling. Medians are presented with 25th and 75th percentiles (boxes), 10th and 90th percentiles (whiskers), and outliers (circles). Medians in the same year with different letters are significantly different (Kruskal–Wallis and Dunn’s tests with Bonferroni corrections; p < 0.05, n = 18–65).
Figure 4.
Length of individual shoots on peony plants in (A) Year 1, (B) Year 2 and (C) Year 3; the total length of shoots per plant in (D) Year 1, (E) Year 2 and (F) Year 3; and the number of leaves per plant in (G) Year 1, (H) Year 2 and (I) Year 3. ‘0Ch’, ‘2Ch’, ‘4Ch’ and ‘6Ch’ = 0, 2, 4 or 6 weeks of chilling at 4 °C, respectively; ‘PC’ = pre-treatment from 20 °C to 8 °C prior to chilling; ‘GA’ = GA3 treatment after chilling. Medians are presented with 25th and 75th percentiles (boxes), 10th and 90th percentiles (whiskers), and outliers (circles). Medians in the same year with different letters are significantly different (Kruskal–Wallis and Dunn’s tests with Bonferroni corrections; p < 0.05, n = 18–65).

Figure 5.
Number of flower buds per peony plant in (A) Year 1, (B) Year 2 and (C) Year 3. ‘0Ch’, ‘2Ch’, ‘4Ch’ and ‘6Ch’ = 0, 2, 4 or 6 weeks of chilling at 4 °C, respectively; ‘PC’ = pre-treatment from 20 °C to 8 °C prior to chilling; ‘GA’ = GA3 treatment after chilling. Medians are presented with 25th and 75th percentiles (boxes), 10th and 90th percentiles (whiskers), and outliers (circles). Medians in the same year with different letters are significantly different (Kruskal–Wallis and Dunn’s tests with Bonferroni corrections; p < 0.05, n = 18–65).
Figure 5.
Number of flower buds per peony plant in (A) Year 1, (B) Year 2 and (C) Year 3. ‘0Ch’, ‘2Ch’, ‘4Ch’ and ‘6Ch’ = 0, 2, 4 or 6 weeks of chilling at 4 °C, respectively; ‘PC’ = pre-treatment from 20 °C to 8 °C prior to chilling; ‘GA’ = GA3 treatment after chilling. Medians are presented with 25th and 75th percentiles (boxes), 10th and 90th percentiles (whiskers), and outliers (circles). Medians in the same year with different letters are significantly different (Kruskal–Wallis and Dunn’s tests with Bonferroni corrections; p < 0.05, n = 18–65).

Figure 6.
Peony plants in full bloom under subtropical conditions after being pre-treated from 20 °C to 8 °C, chilled at 4 °C, and treated with GA3.
Figure 6.
Peony plants in full bloom under subtropical conditions after being pre-treated from 20 °C to 8 °C, chilled at 4 °C, and treated with GA3.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bogiatzis, K.C.; Wallace, H.M.; Trueman, S.J. Shoot Growth and Flower Bud Production of Peony Plants under Subtropical Conditions. Horticulturae 2021, 7, 476. https://doi.org/10.3390/horticulturae7110476
AMA Style
Bogiatzis KC, Wallace HM, Trueman SJ. Shoot Growth and Flower Bud Production of Peony Plants under Subtropical Conditions. Horticulturae. 2021; 7(11):476. https://doi.org/10.3390/horticulturae7110476
Chicago/Turabian StyleBogiatzis, Krista C., Helen M. Wallace, and Stephen J. Trueman. 2021. "Shoot Growth and Flower Bud Production of Peony Plants under Subtropical Conditions" Horticulturae 7, no. 11: 476. https://doi.org/10.3390/horticulturae7110476
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.