The Marine Fauna of New Zealand: Isopoda, Aegidae (Crustacea)
The Marine Fauna of New Zealand: Isopoda, Aegidae (Crustacea)
The Marine Fauna of New Zealand: Isopoda, Aegidae (Crustacea)

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
NATIONAL INSTITUTE OF<br />
WATER AND ATMOSPHERIC RESEARCH (NIWA)<br />
<strong>The</strong> <strong>Marine</strong> <strong>Fauna</strong> <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>:<br />
<strong>Isopoda</strong>, <strong>Aegidae</strong> (<strong>Crustacea</strong>)<br />
Niel L. Bruce<br />
<strong>Marine</strong> Biodiversity and Biosecurity<br />
National Institute <strong>of</strong> Water and Atmospheric Research Ltd<br />
Private Bag 14901, Kilbirnie<br />
Wellington, <strong>New</strong> <strong>Zealand</strong><br />
Present address:<br />
Museum <strong>of</strong> Tropical Queensland<br />
Queensland Museum and School <strong>of</strong> <strong>Marine</strong> and Tropical Biology<br />
James Cook University<br />
70–102 Flinders Street, Townsville 4810, Australia<br />
niel.bruce@qm.qld.gov.au<br />
NIWA Biodiversity Memoir 22<br />
2009
Cataloguing in Publication<br />
BRUCE, N. L.<br />
<strong>The</strong> marine fauna <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>: <strong>Isopoda</strong>, <strong>Aegidae</strong> (<strong>Crustacea</strong>) / by Niel L. Bruce—Wellington:<br />
NIWA (National Institute <strong>of</strong> Water and Atmospheric Research Ltd), 2009<br />
(NIWA Biodiversity Memoir, ISSN 74-0043; 22)<br />
ISBN 978–0–478–23284–4<br />
Series Editor: Dennis P. Gordon<br />
Copy edited and typeset by: Ge<strong>of</strong>f Gregory, Word <strong>The</strong>rapy, Paraparaumu<br />
Printed and bound by: Graphic Press & Packaging Ltd, Levin<br />
Received for publication—12 March 2008<br />
© NIWA Copyright 2009<br />
2
NATIONAL INSTITUTE OF<br />
WATER AND ATMOSPHERIC RESEARCH (NIWA)<br />
<strong>The</strong> <strong>Marine</strong> <strong>Fauna</strong> <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>:<br />
<strong>Isopoda</strong>, <strong>Aegidae</strong> (<strong>Crustacea</strong>)<br />
Niel L. Bruce<br />
<strong>Marine</strong> Biodiversity and Biosecurity<br />
National Institute <strong>of</strong> Water and Atmospheric Research Ltd<br />
Private Bag 14901, Kilbirnie<br />
Wellington, <strong>New</strong> <strong>Zealand</strong><br />
Present address:<br />
Museum <strong>of</strong> Tropical Queensland<br />
Queensland Museum and School <strong>of</strong> <strong>Marine</strong> and Tropical Biology<br />
James Cook University<br />
70-102 Flinders Street, Townsville 4810, Australia<br />
niel.bruce@qm.qld.gov.au<br />
AbstrAct<br />
<strong>The</strong> isopod family <strong>Aegidae</strong> <strong>of</strong> the <strong>New</strong> <strong>Zealand</strong> Exclusive Economic Zone is monographed. Six genera are present in the region:<br />
Aega Leach, 8 5 with eight species, Aegapheles gen. nov. with seven species, Aegiochus Bovallius, 885 with 6, Epulaega gen.<br />
nov. with two, Rocinela Leach, 8 8 with nine, and Syscenus Harger, 1880 with five species. Thirty-nine <strong>of</strong> the 45 species are<br />
named, including two new species <strong>of</strong> Aega, two new species <strong>of</strong> Aegapheles, nine new species <strong>of</strong> Aegiochus, six new species <strong>of</strong><br />
Rocinela, and one each <strong>of</strong> Epulaega gen. nov. and Syscenus; all but seven species are new records for the <strong>New</strong> <strong>Zealand</strong> marine<br />
fauna. Sixteen <strong>of</strong> the named species, approximately 40%, are endemic, but that figure is likely to drop, as many other large<br />
species are known to have extended distributions. Three species are removed from the <strong>New</strong> <strong>Zealand</strong> fauna: Aega novizealandiae<br />
Dana, 853 and Aega cyclops Haswell, 1881 are regarded as nomina dubia and or misidentifications; Rocinela orientalis Schioedte<br />
& Meinert, 879b is regarded as an uncorroborated record. <strong>The</strong> Barybrotidae is reinstated to family rank.<br />
A phylogenetic analysis <strong>of</strong> Aega was conducted using PAUP*, and a new generic classification is proposed, with Aegiochus<br />
Bovallius, 885 revalidated, the subgenus Rhamphion Brusca, 983 placed in synonymy with Aegiochus, and two new genera,<br />
Aegapheles gen. nov. and Epulaega gen. nov., described.<br />
To allow clear characterisation <strong>of</strong> certain <strong>New</strong> <strong>Zealand</strong> species, it was necessary to partially redescribe some Southern<br />
Ocean species: Aegiochus crozetensis (Kussakin & Vasina, 982), Aegiochus uschakovi (Kussakin, 967), and Aega punctulata Miers,<br />
1881; descriptive notes and figures are also provided for Aega angustata Whitelegge, 90 , Aegiochus plebeia (Hansen, 895) and<br />
Syscenus intermedius Richardson, 1910. Supplementary description and figures are given for the Antarctic species Aegiochus<br />
antarctica (Hodgson, 9 0) and Aegiochus glacialis (Tattersall, 92 ). Placed in synonymy are: Aega edwardsii Dollfus, 89 (= A.<br />
punctulata), Aega giganteoculata Nunomura, 988 (= Aegiochus vigilans (Haswell, 88 )), Aega koltuni Kussakin, 967 (= Aegiochus<br />
antarctica (Hodgson, 9 0)) and Syscenus pacificus Nunomura, 98 (= Syscenus latus Richardson, 909); Aega tumida Nunomura,<br />
988 is considered to be indistinguishable from Aegiochus spongiophila (Semper, 867). Species brought out <strong>of</strong> synonymy are:<br />
Aega punctulata Miers, 88 and Aega urotoma Barnard, 9 4.<br />
Keys are provided to the marine genera and to the named <strong>New</strong> <strong>Zealand</strong> species.<br />
Keywords: <strong>Isopoda</strong>, <strong>Aegidae</strong>, Aega, Aegapheles gen. nov., Aegiochus, Epulaega gen. nov., Rocinela, Syscenus, systematics, taxonomy,<br />
new genera, new species, phylogenetic analysis, <strong>New</strong> <strong>Zealand</strong> Exclusive Economic Zone, Southwest Pacific, Southern<br />
Ocean, Antarctic<br />
3
Frontispiece:<br />
Upper left: Aegapheles mahana sp. nov. Upper right: Aega monophthalma Johnston, 834.<br />
Lower: Aegapheles mahana sp. nov.<br />
4
cONtENts<br />
ABSTRACT .............................................................................................................................................................3<br />
INTRODUCTION .................................................................................................................................................7<br />
THE NEW ZEALAND AEGID FAUNA ............................................................................................................8<br />
MATERIAL AND METHODS ............................................................................................................................9<br />
MORPHOLOGy ..................................................................................................................................................<br />
FOSSIL AEGIDAE ............................................................................................................................................... 5<br />
PHyLOGENy ...................................................................................................................................................... 5<br />
TAxONOMy .......................................................................................................................................................25<br />
Suborder Cymothoida Wägele, 989 ..........................................................................................................25<br />
Revalidation and diagnosis <strong>of</strong> Barybrotidae Hansen, 890 .....................................................................25<br />
<strong>Aegidae</strong> White, 850 ......................................................................................................................................26<br />
Key to the marine genera <strong>of</strong> <strong>Aegidae</strong> ......................................................................................................26<br />
Genus Aega Leach, 8 5 .............................................................................................................................27<br />
Key to the <strong>New</strong> <strong>Zealand</strong> species <strong>of</strong> Aega .............................................................................................28<br />
Aega falklandica Kussakin, 967 (Figs 8– ) .........................................................................................28<br />
Aega komai Bruce, 996 (Figs 2, 3) .....................................................................................................34<br />
Aega monophthalma Johnston, 834 (Figs 4– 8) .................................................................................37<br />
Aega semicarinata Miers, 875 (Figs 9–22) ..........................................................................................44<br />
Aega stevelowei sp. nov. (Figs 23–26) .....................................................................................................50<br />
Aega urotoma Barnard, 9 4 (Figs 27–30) .............................................................................................55<br />
Aega whanui sp. nov. (Figs 3 –33) .........................................................................................................6<br />
Aega sp. .....................................................................................................................................................65<br />
Aegapheles gen. nov. ...................................................................................................................................65<br />
Key to the <strong>New</strong> <strong>Zealand</strong> species <strong>of</strong> Aegapheles ...................................................................................65<br />
Aegapheles alazon (Bruce, 2004) comb. nov. (Fig. 34) ..........................................................................66<br />
Aegapheles birubi (Bruce, 2004) comb. nov. (Fig. 35) ...........................................................................68<br />
Aegapheles copidis sp. nov. (Figs 36, 37) ................................................................................................70<br />
Aegapheles hamiota (Bruce, 2004) comb. nov. (Fig. 38) ........................................................................73<br />
Aegapheles mahana sp. nov. (Figs 39–4 ) ...............................................................................................75<br />
Aegapheles rickbruscai (Bruce, 2004) comb. nov. (Fig. 42) ...................................................................79<br />
Aegapheles umpara (Bruce, 2004) comb. nov. (Fig. 43) ........................................................................8<br />
Genus Aegiochus Bovallius, 885 ..............................................................................................................83<br />
Key to the <strong>New</strong> <strong>Zealand</strong> species <strong>of</strong> Aegiochus ....................................................................................84<br />
Aegiochus beri (Bruce, 983), comb. nov. (Figs 44, 45) ........................................................................85<br />
Aegiochus bertrandi sp. nov. (Figs 46–48) ..............................................................................................88<br />
Aegiochus coroo (Bruce, 983), comb. nov. (Figs 49– 0) ......................................................................93<br />
Aegiochus gordoni sp. nov. (Figs 52–55) ................................................................................................97<br />
Aegiochus insomnis sp. nov. (Figs 56–60) ............................................................................................ 03<br />
Aegiochus kakai sp. nov. (Figs 6 –64) .................................................................................................. 09<br />
Aegiochus kanohi sp. nov. (Figs 65–68) ................................................................................................ 5<br />
Aegiochus laevis (Studer, 884), comb. nov. (Figs 69, 70).................................................................. 20<br />
Aegiochus nohinohi sp. nov. (Figs 7 –74) ............................................................................................ 23<br />
Aegiochus piihuka sp. nov. (Figs 75–78) ............................................................................................... 29<br />
Aegiochus pushkini (Kussakin & Vasina, 982), comb. nov. (Figs 79–83) ....................................... 34<br />
Aegiochus riwha sp. nov. (Figs 84–87) ................................................................................................. 4<br />
Aegiochus tara sp. nov. (Figs 88–90) .................................................................................................... 46<br />
Aegiochus vigilans (Haswell, 88 ), comb. nov. (Fig. 9 ) .................................................................. 50<br />
Aegiochus sp. .......................................................................................................................................... 5<br />
5
Epulaega gen. nov...................................................................................................................................... 5<br />
Key to the <strong>New</strong> <strong>Zealand</strong> species <strong>of</strong> Epulaega .................................................................................... 52<br />
Epulaega derkoma sp. nov. (Figs 92–95) ............................................................................................... 52<br />
Epulaega fracta (Hale, 940), comb. nov. (Figs 96–99) ....................................................................... 56<br />
Genus Rocinela Leach, 8 8 ..................................................................................................................... 6<br />
Key to the <strong>New</strong> <strong>Zealand</strong> species <strong>of</strong> Rocinela ..................................................................................... 62<br />
Rocinela bonita sp. nov. (Figs 00– 04) ............................................................................................... 63<br />
Rocinela garricki Hurley, 957 (Figs 05– 09) .................................................................................... 69<br />
Rocinela leptopus sp. nov. (Figs 0– 3) ............................................................................................ 74<br />
Rocinela pakari sp. nov. (Figs 4– 8) ................................................................................................ 78<br />
Rocinela resima sp. nov. (Figs 9– 22) ............................................................................................... 84<br />
Rocinela runga sp. nov. (Figs 23– 25) ................................................................................................ 89<br />
Rocinela satagia sp. nov. (Figs 26– 29) .............................................................................................. 93<br />
Rocinela sp. ............................................................................................................................................. 98<br />
Genus Syscenus Harger, 880 .................................................................................................................. 98<br />
Key to the <strong>New</strong> <strong>Zealand</strong> species <strong>of</strong> Syscenus .................................................................................... 99<br />
Syscenus kapoo sp. nov. (Figs 30, 3 ) ............................................................................................... 99<br />
Syscenus latus Richardson, 909 (Figs 32– 34) ................................................................................202<br />
Syscenus moana Bruce, 2005 (Fig. 35) ................................................................................................206<br />
Syscenus springthorpei Bruce, 997 (Fig. 36) .....................................................................................208<br />
Syscenus sp. ............................................................................................................................................2 0<br />
Uncertain status or records ........................................................................................................................2 0<br />
Aega cyclops Haswell, 88 ...................................................................................................................2 0<br />
Aega novizealandiae Dana, 853, nomen dubium ..............................................................................2<br />
Rocinela orientalis Schioedte & Meinert, 879 ....................................................................................2<br />
Species included in the <strong>Aegidae</strong> ................................................................................................................2 2<br />
Aega Leach, 8 5 ....................................................................................................................................2 2<br />
Aegapheles gen. nov. ..............................................................................................................................2 3<br />
Aegiochus Bovallius, 885 .....................................................................................................................2 3<br />
Alitropus Milne Edwards, 840 ...........................................................................................................2 4<br />
Epulaega gen. nov. .................................................................................................................................2 4<br />
Rocinela Leach, 8 8 ..............................................................................................................................2 4<br />
Syscenus Harger, 880 ...........................................................................................................................2 5<br />
Xenuraega Tattersall, 909 ....................................................................................................................2 5<br />
Ross Sea and Antarctic Islands (Balleny Islands) species ......................................................................2 6<br />
Aegiochus antarctica (Hodgson, 9 0), comb. nov. (Fig. 37) ...........................................................2 6<br />
Aegiochus glacialis (Tattersall, 92 ), comb. nov. (Fig. 38) .............................................................2 9<br />
ACKNOWLEDGEMENTS ...............................................................................................................................222<br />
REFERENCES ....................................................................................................................................................223<br />
APPENDIx . Invalid Aegid names ...............................................................................................................23<br />
APPENDIx 2. Extra-limital species ................................................................................................................232<br />
Aega angustata Whitelegge, 90 (Figs 39, 40) ...............................................................................232<br />
Aega punctulata Miers, 88 (Fig. 4 ) ................................................................................................235<br />
Aegiochus crozetensis (Kussakin & Vasina, 982), comb. nov. (Fig. 42) ........................................237<br />
Aegiochus plebeia (Hansen, 897), comb. nov. (Figs 43, 44) ..........................................................238<br />
Aegiochus uschakovi (Kussakin, 967), comb. nov. (Fig. 45) ...........................................................24<br />
Syscenus intermedius Richardson, 9 0 (Fig. 46) .............................................................................24<br />
APPENDIx 3. Other material examined ........................................................................................................244<br />
APPENDIx 4. Matrices .....................................................................................................................................246<br />
INDEx .................................................................................................................................................................249<br />
6
<strong>The</strong> isopod fauna <strong>of</strong> <strong>New</strong> <strong>Zealand</strong> has received little<br />
attention over the previous two centuries (Bruce 200 ;<br />
Poore & Bruce in press), with only two isopod families<br />
receiving monographic or revisionary treatment, the<br />
Sphaeromatidae by Hurley and Jansen ( 977) and the<br />
Haploniscidae by Lincoln ( 985). Within the Cymothoida<br />
the <strong>Aegidae</strong> have perhaps received least attention.<br />
<strong>The</strong> only documentation following the earliest<br />
carcinological accounts <strong>of</strong> Dana ( 852), Miers ( 876a;<br />
876b) and Thomson and Chilton ( 886) was the record<br />
<strong>of</strong> a beach specimen from the Kermadec Islands by<br />
Chilton ( 9 ), the incidental mention <strong>of</strong> a species by<br />
Hale ( 926: 233, <strong>of</strong> Aega cyclops ‘in <strong>New</strong> <strong>Zealand</strong> area’),<br />
description <strong>of</strong> a single species <strong>of</strong> Rocinela by Hurley<br />
(1957), a misidentification by Stephenson (1980), and,<br />
most recently, popular accounts <strong>of</strong> the family (Bruce<br />
2002, 2003).<br />
Given the low number <strong>of</strong> previously recorded<br />
species (four) from <strong>New</strong> <strong>Zealand</strong>, and the relatively<br />
low number <strong>of</strong> marine species known from Australia<br />
(28) (excluding subantarctic island territories such as<br />
Macquarie Island—including Bruce et al. 2002; Bruce<br />
2004a and those reported here) and South Africa ( )<br />
it comes as a surprise that the <strong>New</strong> <strong>Zealand</strong> EEZ, with<br />
47 species, has the greatest number <strong>of</strong> aegid species <strong>of</strong><br />
any region <strong>of</strong> the world (Australia can be said to have<br />
several regions such as Southern, Indian and Pacific<br />
Oceans, though only two species have been recorded<br />
from other than eastern coasts). <strong>The</strong> East Pacific (see<br />
Brusca 983; Wetzer 990; Brusca & France 992) with<br />
5 species and North Atlantic (see Kussakin 979;<br />
Brand & Andres 2008; Bruce 993a) with 8 species are<br />
relatively well documented and the recorded diversity<br />
is probably close to actuality. Australian aegids have<br />
received some attention (Bruce 983; 988; 997a,b;<br />
2004a; Bruce et al. 2002) and my own examination <strong>of</strong><br />
museum collections in Australia has revealed numerous<br />
as yet undescribed species. In terms <strong>of</strong> area, <strong>New</strong><br />
<strong>Zealand</strong> has the greatest diversity <strong>of</strong> <strong>Aegidae</strong>.<br />
<strong>The</strong> area loosely termed the ‘southwestern Pacific’,<br />
stretching from eastern Papua <strong>New</strong> Guinea, through<br />
the island nations arc to <strong>New</strong> <strong>Zealand</strong> and westwards<br />
to the Australian coasts is the region <strong>of</strong> greatest known<br />
diversity for the family <strong>Aegidae</strong>. Including ‘known<br />
undescribed’ species, there are 72 aegid species* from<br />
* Trilles and Justine (2004) recorded three species <strong>of</strong> Aega<br />
from <strong>New</strong> Caledonia. Two <strong>of</strong> these are misidentifications,<br />
the third is not identified to species but is possibly one<br />
<strong>of</strong> the species described by Bruce (2004b). <strong>The</strong>se species<br />
are not included in the species totals given here.<br />
INtrOductION<br />
7<br />
this region, some 60% [the figures change as new<br />
records and species are discovered] <strong>of</strong> the species<br />
recorded worldwide. Museum collections that I have<br />
examined (e.g. USNM, Smithsonian Institution; <strong>The</strong><br />
Natural History Museum, London; Muséum national<br />
d’Histoire naturelle, Paris; and Zoologisk Museum,<br />
Copenhagen) do not indicate that other regions would<br />
have a diversity as great and as yet undocumented.<br />
Collections held at various Australian museums, and<br />
material collected around <strong>New</strong> Caledonia, indicate<br />
that many species remain to be described from the<br />
southwestern Pacific.<br />
symbIOsEs<br />
<strong>Aegidae</strong> are well-known associates <strong>of</strong> fishes, almost<br />
exclusively attaching temporarily to the external<br />
surfaces. A small number <strong>of</strong> species are associated<br />
with other invertebrates, notably sponges. Klitgaard<br />
( 995) found that Aegiochus ventrosa used only one<br />
<strong>of</strong> eleven examined species <strong>of</strong> sponge sampled in the<br />
northeastern Atlantic. Aegiochus lethrina, an associate<br />
<strong>of</strong> coral-reef fishes, has also been recorded from<br />
sponges (Bruce 983). <strong>The</strong>re is one record <strong>of</strong> an Aegiochus<br />
from the cloaca <strong>of</strong> an ascidian (Wetzer 990). In<br />
<strong>New</strong> <strong>Zealand</strong>, Aegiochus piihuka sp. nov. is associated<br />
with hexactinellid sponges, and Epulaega fracta and<br />
Aegiochus spongiophila have also been recorded from<br />
hexactinellids (Nunomura 988a). Records <strong>of</strong> aegids<br />
attached to squid (e.g. Bruce 996) are regarded as<br />
unconfirmed at present.<br />
mIcrOprEdAtOrs Or pArAsItEs?<br />
<strong>Aegidae</strong> are here regarded as micropredators rather<br />
than parasites (see Bruce 2003, 2004a; Brusca 983).<br />
Parasites are, variously defined, symbionts (e.g. Rohde<br />
982, 2005), and are widely regarded as having some<br />
manner <strong>of</strong> perceived deleterious or harmful effect<br />
on the host. Generally a permanent trophic adult association<br />
is noted between the parasite and the host<br />
individual. Aegids do not fulfil these criteria, and while<br />
attacking and feeding on their victims they rarely form<br />
a permanent association with their ‘host’, but instead<br />
detach following their feed.<br />
In a few instances it is known that a species forms<br />
a more long-term attachment (e.g. Syscenus—see Ross<br />
et al. 200 ). Others, such as Aegiochus lethrina, appear<br />
to be feeding on fish mucus within the ‘host’ nasal<br />
passages, rather than blood or tissue. Wägele ( 990)<br />
gave a brief and incidental description <strong>of</strong> the mode <strong>of</strong>
feeding <strong>of</strong> Aegiochus antarctica in aquaria feeding on<br />
provided prey species (namely North Atlantic plaice),<br />
noting that the isopod spent most <strong>of</strong> the time inactive<br />
in a burrow, emerging only to search for prey.<br />
<strong>The</strong>re is one reported instance <strong>of</strong> an aegid, Rocinela<br />
signata Schioedte and Meinert, 879a, attaching to the<br />
gills and inside <strong>of</strong> the mouth <strong>of</strong> the host (de Lima<br />
et al. 2005). Salmon under aquaculture have been<br />
reported as being attacked and killed by Rocinela belliceps<br />
by Novotny and Mahnken ( 97 ), and Nair and<br />
Nair (1983) reported that fish attacked by Alitropus in<br />
aquarium conditions became anaemic. Wing and Moles<br />
( 995) showed that under aquarium conditions Rocinela<br />
angustata preferentially attacks some prey species.<br />
<strong>The</strong>re are few data available on the feeding habits<br />
<strong>of</strong> aegid isopods; they are here considered to be micropredators,<br />
and the fish that they have been recorded<br />
from as prey.<br />
dIstrIbutION<br />
<strong>Aegidae</strong> are distributed throughout the world oceans,<br />
from the tropics to polar waters. Broadly, <strong>Aegidae</strong> are<br />
marine with a depth range from shallow or surface<br />
depths (such as species attaching to shallow-water<br />
coral-reef fishes) to a depth <strong>of</strong> 4609 metres, although<br />
most species (depth data are not available for a substantial<br />
number <strong>of</strong> species) are recorded from the<br />
continental shelf and rise at depths from less than 00<br />
metres to approximately 200 metres. Twenty species<br />
are known from depths in excess <strong>of</strong> 200 metres, and <strong>of</strong><br />
these six are from depths greater than 2000 metres.<br />
<strong>The</strong> large genera (Aega, Aegapheles, Aegiochus, Rocinela<br />
and Syscenus) are found throughout the world<br />
oceans. <strong>The</strong> genus Epulaega gen. nov. has an Indo-<br />
Pacific range with one species from South Africa, the<br />
remainder from the western Pacific. <strong>The</strong> monotypic<br />
genus Xenuraega is known only from the North Atlan-<br />
<strong>The</strong> two large genera Aega sensu lato and Rocinela<br />
have dominated the family and collections <strong>of</strong> aegids<br />
worldwide. Although Aega is here restructured to<br />
four genera, the <strong>New</strong> <strong>Zealand</strong> fauna has most species<br />
here recorded belonging to Rocinela and those genera<br />
that form the Aega clade. All <strong>of</strong> these genera may be<br />
relatively well represented at high latitudes. <strong>The</strong> genus<br />
Syscenus, mesopelagic fish micropredators, while<br />
possibly common (Ross et al. 200 ), is infrequently<br />
collected. Under the new classification presented here<br />
there are six genera occurring in <strong>New</strong> <strong>Zealand</strong> waters.<br />
<strong>The</strong> two remaining monotypic genera have not been<br />
recorded from <strong>New</strong> <strong>Zealand</strong> waters. Alitropus Milne<br />
Edwards, 840 is a tropical freshwater genus known<br />
thE NEw ZEAlANd AEgId FAuNA<br />
8<br />
tic. Alitropus, also monotypic, is restricted to freshwater<br />
habitats in the Indo-Malaysian region, extending at<br />
least to eastern Australia (Bruce 983). Two genera,<br />
Syscenus and Xenuraega, appear to be mesopelagic<br />
throughout their range.<br />
<strong>The</strong>re appear to be no obvious endemic groupings<br />
except within Aega where those species belonging to<br />
the two clades <strong>of</strong> species related to Aega angustata and<br />
to Aega antennata (Fig. 6, p. 9) are both restricted to the<br />
western Pacific, with both described and undescribed<br />
species occurring from Japan to southern Australia and<br />
eastwards to <strong>New</strong> Caledonia and <strong>New</strong> <strong>Zealand</strong>.<br />
Individual species may be widely distributed,<br />
such as Aega monophthalma, occurring in the Atlantic,<br />
Indian and Pacific Oceans, and ranging from cold highlatitude<br />
water to the tropics, or Aegapheles alazon, which<br />
occurs from South Africa to <strong>New</strong> <strong>Zealand</strong> and north<br />
to tropical and subtropical locations in both the Indian<br />
and Pacific Oceans. While many species have restricted<br />
ranges such as the Tasman Sea or southwestern Pacific,<br />
local endemism is generally lower than for those<br />
families <strong>of</strong> free-living isopods such as the Cirolanidae<br />
or Sphaeromatidae.<br />
In the genus Rocinela the distributional pattern<br />
is somewhat different from that <strong>of</strong> the genera Aega,<br />
Aegapheles, and Aegiochus in that no species occurs in<br />
all major oceans, and no species has both Northern<br />
and Southern Hemisphere ranges. Most species are<br />
regional endemics, being largely restricted to a region<br />
such as <strong>New</strong> <strong>Zealand</strong> (all <strong>New</strong> <strong>Zealand</strong> Rocinela are<br />
endemic, though it is probable that some extend<br />
towards <strong>New</strong> Caledonia), northern Pacific, or East<br />
Pacific, for example.<br />
While species-level endemism <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>’s<br />
<strong>Aegidae</strong> sits at 50% it is probable that this would<br />
drop lower with more complete documentation <strong>of</strong> the<br />
<strong>Aegidae</strong> <strong>of</strong> eastern Australia and southwestern Pacific<br />
island nations.<br />
from Indo–Australasia (Bruce 983; Ho & Tonguthai<br />
992); Xenuraega Tattersall, 909 is a blind, highly<br />
adapted bathypelagic genus, presently known only<br />
from the North Atlantic (Bruce 993a). Alitropus is<br />
absent from <strong>New</strong> <strong>Zealand</strong> waters, but Xenuraega is best<br />
considered to be <strong>of</strong> uncertain distribution, particularly<br />
as some pelagic and mesopelagic isopods do have<br />
worldwide distributions [e.g. Metacirolana caeca<br />
(Hansen, 9 6), Svavarsson & Bruce 2000], including<br />
some aegids such as Aega monophthalma Johnston,<br />
834; others, such as Aegapheles alazon Bruce, 2004 and<br />
several <strong>of</strong> the Southern Ocean species reported here,<br />
have extended Southern Hemisphere ranges.
mAtErIAl ExAmINEd<br />
Material examined includes that which was referred<br />
to in preparing descriptions (commenced in 2002).<br />
‘Also examined’ is used for comparative material <strong>of</strong><br />
other species. ‘Additional material’ includes specimens<br />
belonging to a species that was identified subsequent<br />
to the preparation <strong>of</strong> the description.<br />
gEOgrAphIc lImIts<br />
<strong>The</strong> defining area for inclusion in this monograph is<br />
that <strong>of</strong> the <strong>New</strong> <strong>Zealand</strong> chart area, NIWA Chart No<br />
73 (CANZ 997), extending beyond the recognised<br />
Exclusive Economic Zone (EEZ) (Fig. ). <strong>The</strong> boundaries<br />
<strong>of</strong> this region lie at approximately 24–57°S and<br />
57°E– 67°W, and as such potentially include records<br />
from the vicinity <strong>of</strong> the Australian territories <strong>of</strong> Lord<br />
Howe Island, Norfolk Island, and Macquarie Island.<br />
Aegids rarely occupy restricted coastal ranges, <strong>of</strong>ten<br />
being wide-ranging, so it is pertinent to include records<br />
from beyond territorial waters and the EEZ.<br />
dEscrIptIONs<br />
All descriptions were prepared using DELTA (Descriptive<br />
Language for Taxonomy: Dallwitz et al. 997).<br />
Separate data sets (suites <strong>of</strong> characters) are used for<br />
species within the Aega clade and the Rocinela–Syscenus<br />
clade and to diagnose the genera. Diagnoses are complementary<br />
to the description for higher taxa, and<br />
therefore the information is not repeated in the following<br />
description. Principal terms used in descriptions<br />
are shown in Figs 2 and 3.<br />
For integer numeric character states, the description<br />
may include a zero (0) rather than the more usual ‘without’<br />
or ‘none’; similarly for some real numeric characters<br />
it may read ‘ .0 times as long as’ rather than the simpler<br />
‘as long as’. Minor details qualifying a coded character<br />
state are retained within parentheses.<br />
Colour has not been included in the descriptions<br />
owing to post-mortem changes and subsequent fading<br />
in preserved specimens. Live colour in aegids is rarely<br />
observed or photographed so there are few comparative<br />
data. Eye colour in aegids can be red, black, dark<br />
brown, light brown or bronze (‘golden’). Eye colour<br />
is not always consistent within species. Some aegids<br />
have a noticeable white perimeter to each ommatidium,<br />
giving the eye a reticulate appearance, but in others the<br />
centre <strong>of</strong> the ommatidium is white.<br />
<strong>The</strong> uropodal endopod <strong>of</strong> all <strong>Aegidae</strong> (and most<br />
families <strong>of</strong> Cymothoida) has, on the lateral margin, a<br />
heavily plumose articulating seta that is set in a small<br />
mAtErIAl ANd mEthOds<br />
9<br />
notch, usually at a position about the distal one-third<br />
<strong>of</strong> the length <strong>of</strong> the lateral margin. This seta defines<br />
the proximal and distal portions <strong>of</strong> the endopod lateral<br />
margin, and counts <strong>of</strong> robust setae are given in relation<br />
to this point.<br />
drAwINgs ANd dIssEctIONs<br />
Unless otherwise stated, all drawings except for the<br />
mouthparts were made using a Leica M 2.5 stereo-<br />
microscope, using both reflected and transmitted light.<br />
All appendages were dissected from the right-hand<br />
side <strong>of</strong> the specimen unless otherwise stated. Appendages<br />
were drawn without being flattened, and while<br />
perspective has been kept as consistent as possible,<br />
allowances must be made for some differences in the<br />
drawings from measurements given. Mouthparts were<br />
dissected and mounted unstained in lactic acid (88%)<br />
and examined and drawn using a Zeiss Axioskop 2plus<br />
compound microscope. Mouthparts <strong>of</strong> some small<br />
species were remounted in ‘Aquamount Improved’<br />
Gurr, all other dissected appendages were placed into<br />
micro-vials and stored with the dissected specimen. All<br />
drawings were made using a camera lucida.<br />
Dissection <strong>of</strong> historical type material and fragile<br />
specimens was kept to a minimum (usually no dissection).<br />
Permission to dissect any material described<br />
wholly or in part by O. G. Kussakin was not granted,<br />
and the borrowed material was not accompanied by<br />
the dissected appendages.<br />
In order to maintain a reasonable brevity <strong>of</strong> text,<br />
some reduction <strong>of</strong> drawings and <strong>of</strong> description has<br />
been undertaken. In general, pereopods 2 and 3 are<br />
similar to each other as are pereopods 5–7. Pereopod 4<br />
is intermediate in form between anterior ( –3) and posterior<br />
(5–7) pereopods. For many species pereopods –3<br />
and pereopods 6 and 7 are illustrated, but pereopods<br />
3 and 6 are not described in detail. Similarly pleopods<br />
within genera are remarkably uniform, and for some<br />
species only pleopods and 2 are illustrated.<br />
<strong>The</strong> maxilliped palp is twisted obliquely and bent<br />
ventrally in relation to the plane <strong>of</strong> the base <strong>of</strong> the<br />
maxilliped. This makes it difficult to draw and to observe,<br />
in particular palp article 5 is <strong>of</strong>ten obscured. In<br />
small species the palp will flatten under a cover slip<br />
if cleared in lactic acid. In large species article 5 was<br />
observed directly and occasionally broken away from<br />
the palp itself. In most cases the number <strong>of</strong> robust setae<br />
is not critical in making a species identification and the<br />
accuracy <strong>of</strong> these counts, particularly for small setae,<br />
should not taken to be potentially indicative.
Figure 1. Map <strong>of</strong> the <strong>New</strong> <strong>Zealand</strong> Region (based on CANZ 977) showing boundary <strong>of</strong> the Exclusive Economic<br />
Zone (EEZ) and major place names.<br />
I do not consider the number <strong>of</strong> plumose marginal<br />
setae on the pleopods to be significant in differentiating<br />
species as it is likely to be size-dependent. For small<br />
species (< 0 mm), these differences may be informative,<br />
but the extent <strong>of</strong> the PMS (where the setae start on<br />
each margin) is more useful. For all species <strong>of</strong> a length<br />
greater than 5 mm, I have not given counts for the<br />
pleopod marginal setae.<br />
0<br />
mEAsurEmENts<br />
Whole specimens were measured in lateral view using<br />
a micrometer eyepiece, along the axis <strong>of</strong> the join<br />
between the coxae and pereonites. Owing to curvature<br />
<strong>of</strong> many specimens on fixation, dorsal views <strong>of</strong><br />
specimens are <strong>of</strong>ten foreshortened. Many aegids are<br />
large, between 3 cm and 6 cm, and may stretch or
end on preservation, rendering apparently precise<br />
measurements meaningless. <strong>The</strong>refore, lengths for<br />
specimens <strong>of</strong> 20 mm or more are given to the nearest<br />
millimetre. Pereopod measurements were made along<br />
the axis <strong>of</strong> the articles for the basis <strong>of</strong> pereopods –3<br />
and all articles for pereopod 7; for pereopods –3 the<br />
ischium, merus and carpus were measured along the<br />
inferior margin.<br />
tErmINOlOgy<br />
Words used in descriptions are shown in Figs 2 and 3.<br />
Setae, unless stated otherwise, are simple (following<br />
Watling 989).<br />
AbbrEvIAtIONs<br />
Institutional<br />
AK — Auckland Institute and Museum, Auckland<br />
AM —Australian Museum, Sydney<br />
BMNH — <strong>The</strong> Natural History Museum, London<br />
LACM — Natural History Museum <strong>of</strong> Los Angeles<br />
County, Los Angeles<br />
MNHN — Muséum national d’Histoire naturelle,<br />
Paris<br />
MTQ — Queensland Museum, Museum <strong>of</strong> Tropical<br />
Queensland, Townsville<br />
NMV — Museum Victoria, Melbourne<br />
NIWA – National Institute <strong>of</strong> Water and Atmospheric<br />
Research Ltd, Wellington<br />
body<br />
Body lacking processes with rare exception, such as the<br />
males <strong>of</strong> Aegiochus vigilans (Haswell, 88 ) (see Bruce<br />
983) and Aegiochus webberi (Nierstrasz, 93 ).<br />
rostral point<br />
Present in all genera. In Aega this is usually a distinct,<br />
acute anteriorly directed process, in Aegiochus it is<br />
ventrally and posteriorly bent and in Epulaega gen. nov.<br />
it is minute and in dorsal view the head may appear<br />
to lack a rostral point. In Rocinela it is a large flat and<br />
anteriorly rounded process. In Syscenus and Xenuraega<br />
ranges from moderate to small in size.<br />
Eyes<br />
Range in size from small (infrequent), cirolanid-like<br />
proportions as in Aegiochus laevis (Studer, 884) to<br />
huge, filling the entire head as in many species illustrated<br />
here. It is notable that in many species the eyes<br />
are distinctly dorsal, with ommatidia not extending<br />
to the ventral surface, and not lateral as in cirolanids<br />
and many other Cymothoida. In Aega the surface <strong>of</strong><br />
the eye is smooth, while in Rocinela the surface <strong>of</strong> each<br />
NMNZ – National Museum <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>, Te Papa<br />
Tongarewa, Wellington<br />
NTM — Museum and Art Gallery <strong>of</strong> the Northern<br />
Territory, Darwin<br />
QFS — Queensland Fisheries Service (now part <strong>of</strong> DPI,<br />
Brisbane)<br />
QM — Queensland Museum, Brisbane<br />
SAM — South Australian Museum, Adelaide<br />
SafM — South African Museum, Cape Town<br />
USNM — National Museum <strong>of</strong> Natural History, Smithsonian<br />
Institution, Washington DC<br />
ZIAS — Zoological Institute, Academy Sciences, Leningrad<br />
ZMA — Zoological Museum, Amsterdam<br />
ZMHA — Zoological Museum, Hamburg<br />
ZMUC — Zoologisk Museum, University <strong>of</strong> Copenhagen<br />
morphological<br />
BL— body length<br />
RS—robust seta/setae<br />
PMS—plumose marginal setae<br />
NAmEs<br />
mOrphOlOgy<br />
Names for new taxa other than place names and honorifics<br />
are derived from Biggs (1990) for Mäori names<br />
and Brown ( 956) for traditional classical names.<br />
Nomenclature for fishes has been sourced entirely<br />
from FishBase (Froese & Pauly 2002–07).<br />
ommatidium is distinctly rounded giving a nodular<br />
appearance to the eyes.<br />
pleon<br />
Relatively uniform throughout the family, all genera<br />
with five free (not fused) segments. Differences can<br />
be observed in the degree <strong>of</strong> prolongation <strong>of</strong> pleonite<br />
4 in Aega and the extent to which the posterolateral<br />
margins are acute.<br />
pleotelson<br />
Varies with regard to shape <strong>of</strong> the margins, setation<br />
and ornamentation.<br />
Antennule and antenna<br />
<strong>The</strong> antennule differs between genera in the degree <strong>of</strong><br />
flattening <strong>of</strong> peduncular articles 1 and 2, the relative<br />
extension <strong>of</strong> the distolateral angle <strong>of</strong> peduncular article<br />
2, and the relative proportions <strong>of</strong> peduncular article 3;<br />
the length <strong>of</strong> the flagellum may separate species. <strong>The</strong><br />
antennal peduncle is relatively uniform, with the first<br />
two articles always short; in Aega and related genera,<br />
peduncular article 3 is also relatively short; in Rocinela
Figure 2. Terms and positions used in descriptions: A, lateral view; B, dorsal head; C, ventral head; D, pleopod; E,<br />
pereopod ; F, pereopod 7; G, uropod.<br />
2
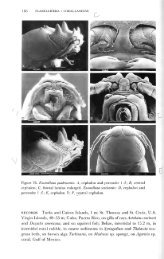
Figure 3. Terms used in descriptions for mouthparts;<br />
maxilliped palp article numbered.<br />
and Syscenus, this article is proportionally longer, about<br />
twice as long as the preceding article; flagellum length<br />
is variable, from longer than the body in Xenuraega to a<br />
little longer than the peduncle in some species.<br />
mouthparts<br />
<strong>The</strong> mandible is simple, with a narrow, distally acute<br />
uni- or bidentate incisor (occasionally weakly tridentate);<br />
the molar process is usually present as a small<br />
but distinct flat lobe, occasionally serrate (e.g. Aega<br />
vigilans, see Bruce 1983) and when small it is difficult<br />
to observe; the mandible palp is uniform throughout<br />
the family, but unusually seems to have the basal article<br />
arising from what appears to be a large articulated<br />
(non-cuticularised) area giving rise to the appearance<br />
<strong>of</strong> four distinct articles (as misinterpreted by Bruce<br />
983, 988). This area is considered to be part <strong>of</strong> the<br />
mandible.<br />
<strong>The</strong> maxillule is remarkably uniform, and consists<br />
<strong>of</strong> a short simple mesial lobe and the elongate lateral<br />
lobe which is provided with 5– 0 robust setae. <strong>The</strong>se<br />
setae vary from broad-based triangular in shape to<br />
slender, and may be hooked, hammer-head or falcate;<br />
they are always terminally acute. <strong>The</strong> mesial lobe is<br />
small and <strong>of</strong>ten lost in dissection, even from large<br />
specimens; this lobe appears to be absent from Rocinela,<br />
Syscenus, and Xenuraega, but present or absent in Aega,<br />
Aegiochus, and related genera.<br />
<strong>The</strong> maxilla is elongate and flattened, with a small<br />
distomesial lobe (the basal endite <strong>of</strong> Brandt & Poore<br />
2003). <strong>The</strong> distal margin is twisted and bent ventro-<br />
laterally so that illustrations made from slide-mounted<br />
preparations never show the true shape. Setation is<br />
uniform with the lateral lobe having 3–5 hooked robust<br />
setae, the mesial lobe with 2–4, one <strong>of</strong> which is<br />
usually straight.<br />
<strong>The</strong> maxilliped palp varies in the number <strong>of</strong> articles,<br />
these differences being diagnostic for different genera.<br />
<strong>The</strong> palp is not flat, being twisted and bent ventrally.<br />
In Aega and Aegapheles, maxilliped palp article 5 is difficult<br />
to observe by light microscopy (as evidenced by<br />
frequent errors <strong>of</strong> interpretation in the literature) as it<br />
is either largely or wholly concealed by article 4, or can<br />
be viewed only from the side. A maxilliped endite is<br />
present in most genera, and is usually small, usually<br />
provided with small simple setae, occasionally larger<br />
with long circumplumose setae (e.g. Aegiochus riwha<br />
sp. nov.) similar to those <strong>of</strong> cirolanids. Critical differences<br />
in the setation <strong>of</strong> maxilliped palp article 5 were<br />
observed by Brusca ( 983), who used these differences<br />
in support <strong>of</strong> his proposed subgenera Aega (Aega) and<br />
Aega (Rhamphion). <strong>The</strong> subgenus Aega was defined as<br />
having ‘stout recurved spines’ on maxilliped article 5<br />
with Rhamphion having ‘long, stout, simple, setae, but<br />
rarely recurved spines.’ However, these differences<br />
have not been found to be sustained on closer examination.<br />
Maxillipedal palp article 5 in some species <strong>of</strong> Aega<br />
and Aegapheles appears partially fused to article 4; in<br />
some species all the robust setae are elongate, in others<br />
article 5 has both elongate and hooked robust setae.<br />
3
pereopods<br />
<strong>The</strong>se are characteristically robust, as is so for most<br />
Cymothoida. <strong>The</strong> dactylus <strong>of</strong> the anterior pereopods<br />
(pereopods –3) is described as hooked or prehensile,<br />
and in most species the dactylus is strongly curved and<br />
.4 to .7 times as long as the propodus, but occasionally<br />
only as long as the propodus. In some species the<br />
dactylus is weakly prehensile (e.g. Epulaega derkoma<br />
sp. nov., Aegiochus riwha sp. nov. and Rocinela leptopus<br />
sp. nov.). <strong>The</strong> anterior pereopods generally have few<br />
slender and robust setae, the robust setae in some species<br />
being large. <strong>The</strong> propodus in both Aega and Rocinela<br />
may have a lobe or blade on the inferior margin,<br />
which in some species may be large. In Rocinela this<br />
lobe has prominent robust setae along the free margin.<br />
<strong>The</strong> posterior pereopods (pereopods 4–7) usually lack<br />
abundant slender setae and the inferior and distal margins<br />
are variously ornamented with robust setae.<br />
brood pouch<br />
Uniform throughout the family when details have been<br />
recorded, consisting <strong>of</strong> overlapping oostegites arising<br />
from coxae 2–5; without posterior pocket.<br />
penes<br />
Either sessile (i.e. opening flush with the surface <strong>of</strong><br />
sternite 7) or in the form <strong>of</strong> low tubercles, only occasionally<br />
(e.g. Aegiochus vigilans) in the form <strong>of</strong> flat lobes as<br />
seen in many cirolanids or sphaeromatids. <strong>The</strong> penial<br />
openings are usually separate, occasionally adjacent<br />
to each other, occasionally united.<br />
pleopods<br />
Remarkably uniform throughout the family, with<br />
useful differences evident in the shape <strong>of</strong> the rami <strong>of</strong><br />
pleopod , the extent to which the margins carry plumose<br />
setae and also the ornamentation <strong>of</strong> the peduncle;<br />
the margins <strong>of</strong> the rami are usually even or weakly<br />
serrate—in a few species there are prominent intersetal<br />
serrations, referred to as digitate in descriptions.<br />
<strong>The</strong> appendix masculina is basal in the Aega group <strong>of</strong><br />
genera, sub-basal in Rocinela, and usually simple and<br />
straight, <strong>of</strong>ten shorter than the endopod, but occasionally<br />
longer (notably Aegiochus vigilans); it is sometimes<br />
sinuate or armed with cuticular scales (e.g. Aegiochus<br />
tiaho sp. nov. and Aegiochus kakai sp. nov.).<br />
uropods<br />
Flat and lamellar in all genera except Xenuraega which<br />
has a filamentous exopod and the endopod reduced to<br />
a stub (Bruce 993a). In most species the plane <strong>of</strong> the<br />
exopod and endopod are about the same, the exopod<br />
with the lateral margin weakly tilted dorsally; in Rocinela<br />
and species <strong>of</strong> Aegapheles gen. nov. the plane <strong>of</strong><br />
the exopod can be strongly angled. Uropodal margins<br />
show a variety <strong>of</strong> setation patterns, with robust setae<br />
nearly always present on all margins in species <strong>of</strong> Aega,<br />
less evident in Rocinela and Syscenus.<br />
4<br />
sExuAl vArIAtION IN thE AEgIdAE<br />
In general, other than for the primary sexual characters<br />
(penial processes, appendix masculina, oostegites)<br />
there is remarkably little difference between males and<br />
females. In some species <strong>of</strong> Aega, females, particularly<br />
ovigerous females, may have wider body proportions<br />
than males, and the maxilla and maxilliped, in Aega,<br />
become covered in scale-setae, and the characteristic<br />
recurved or hooked robust setae are replaced by<br />
plumose setae; the characteristic shape <strong>of</strong> the male<br />
maxilliped article 5 is also not shown. For those species<br />
that have nodular or other such ornamentation it<br />
is more strongly developed in the male. <strong>The</strong> exception<br />
seems to be Rocinela, in which mature males may have<br />
a broader body shape, more setose maxilliped and,<br />
when it has been recorded, uropodal rami with dense<br />
marginal setae. Rocinela is also unusual that in some<br />
species eye size varies with maturity, small juveniles<br />
and mancas having proportionally larger eyes than<br />
do mature specimens. In some females <strong>of</strong> both Aega<br />
and Rocinela the robust setae <strong>of</strong> the anterior pereopods<br />
become more slender than in the males.<br />
sIblINg spEcIEs ‘FlOcKs’ wIthIN thE<br />
AEgIdAE<br />
Species <strong>of</strong> <strong>Aegidae</strong> have <strong>of</strong>ten, in the past, been differentiated<br />
using conspicuous morphological characters,<br />
and sibling species or ‘species swarms’ such as those<br />
<strong>of</strong> the ‘Cirolana parva-group’, or species <strong>of</strong> the sphaeromatid<br />
genus Oxinasphaera (e.g. Bruce 997b, 2004b),<br />
have not previously been reported for the family.<br />
Many species <strong>of</strong> <strong>Aegidae</strong> have been considered to be<br />
both variable and widely distributed. Recently, Bruce<br />
(2004a) showed that the supposedly globally distributed<br />
species Aegapheles deshaysiana (Milne Edwards,<br />
840) was a group <strong>of</strong> some 2 species, many <strong>of</strong> which<br />
proved, once described, to be distinctive, with only a<br />
few <strong>of</strong> those species being sibling species in the sense<br />
<strong>of</strong> being near identical.<br />
In describing species from the southwestern Pacific<br />
it has become increasingly apparent that groups <strong>of</strong><br />
closely similar species exist within the genera <strong>of</strong> the<br />
<strong>Aegidae</strong>. It is implicit that an increasing level <strong>of</strong> fine<br />
morphological character discrimination will come into<br />
use in order to separate these species. Examples in the<br />
present work include the species pair <strong>of</strong> Aegiochus beri<br />
(Bruce, 983) and Aegiochus riwha sp. nov., the closely<br />
similar species centring around Aegapheles alazon<br />
(Bruce, 2004), the sibling species related to Aegiochus<br />
coroo (Bruce, 983) and Aegiochus bertrandi sp. nov.,<br />
and species related to Aegiochus plebeia (Hansen, 897)<br />
and Aegiochus ventrosa (M. Sars, 859). <strong>The</strong>se are some<br />
examples, but there are more species groups <strong>of</strong> this sort.<br />
Elucidation <strong>of</strong> such complexes <strong>of</strong> species is confounded
y the fact that while some species do have regionally<br />
localised ranges, others may be found throughout the<br />
major oceans, and sibling species may also be at least<br />
partly geographically sympatric.<br />
Sibling species groups within the <strong>Aegidae</strong> have a<br />
characteristic near identical somatic morphology, antennules,<br />
antenna, frontal lamina and general appearance<br />
<strong>of</strong> the pleopods. Notwithstanding their overall<br />
similarity, these species can be discriminated and<br />
<strong>The</strong>re are no unambiguous records <strong>of</strong> <strong>Aegidae</strong> in the<br />
fossil record. Recently Polz (2005) described and placed<br />
Brunnaega Polz, 2005 and the sole species B. roeperi<br />
Polz, 2005 into the <strong>Aegidae</strong>. <strong>The</strong> basis for assigning<br />
the specimen to the <strong>Aegidae</strong> appeared to be that the<br />
fossil specimen did not fit the diagnosis <strong>of</strong> Palaega<br />
Woodward, 870 (Cirolanidae), but no explanation was<br />
given as to why the family <strong>Aegidae</strong> was considered<br />
more appropriate than the Cirolanidae, Corallanidae or<br />
Tridentellidae. Most recently described fossil isopods<br />
<strong>of</strong> the Cymothoida that lack an obviously spinose pleo-<br />
ANAlysIs OF AegA<br />
<strong>The</strong> <strong>Aegidae</strong> is one <strong>of</strong> the large group <strong>of</strong> families now<br />
placed in the recognised paraphyletic Cymothooidea<br />
<strong>of</strong> Brandt and Poore (2003), this superfamily including<br />
those families generally known to associate with<br />
or parasitise fish during at least one phase <strong>of</strong> their<br />
life history (the Anuropidae being an exception). <strong>The</strong><br />
relationship <strong>of</strong> the <strong>Aegidae</strong> to the other cymothooid<br />
families is not clear, some analyses (e.g. Brandt & Poore<br />
2003) placing the family as the sister group to the Cymothoidae<br />
plus ‘Epicaridea’, while molecular analyses<br />
suggest the <strong>Aegidae</strong> could be the sister group to the<br />
Cirolanidae (Dreyer & Wägele 2002) or to the Cirolanidae<br />
and Corallanidae (Dreyer & Wägele 200 ). Wägele<br />
( 989) also questioned the monophyly <strong>of</strong> the <strong>Aegidae</strong>,<br />
as the morphology <strong>of</strong> the maxilliped palp in the genera<br />
Rocinela and Syscenus is much the same as that <strong>of</strong> the<br />
Cymothoidae. <strong>The</strong> close relationship between the Cirolanoidea<br />
and several families <strong>of</strong> the Cymothooidea<br />
is emphasised by the several species <strong>of</strong> <strong>Aegidae</strong>, Corallanidae<br />
and Tridentellidae that retain the tridentate<br />
mandible incisor, one <strong>of</strong> the two identified apomorphic<br />
states for the Cirolanoidea. It is difficult to homologise<br />
lost or reduced morphological character states, and at<br />
FOssIl AEgIdAE<br />
phylOgENy<br />
characterised using morphological criteria. Consistent<br />
differences are to be found in: the details <strong>of</strong> pereopod<br />
proportions and setation; details, sometimes subtle, in<br />
the shape, proportions and setation (size and pattern)<br />
<strong>of</strong> the uropods; and the shape and setation <strong>of</strong> the posterior<br />
margins <strong>of</strong> the pleotelson. While some <strong>of</strong> these<br />
characters are <strong>of</strong> a finer resolution than previously<br />
used, they are usually found to be highly consistent<br />
and species-specific once identified.<br />
telson posterior margin have been placed in Cirolana<br />
(see Weider & Feldmann 992 for a detailed discussion),<br />
and consequently most cymothooid genera are<br />
placed in the Cirolanidae (e.g. Feldmann & Goolaerts<br />
2005; Wieder & Feldmann 992), or as ‘family uncertain’<br />
(Brandt et al. 999). Brunnaega is better placed in<br />
the Cirolanidae as it agrees well with the form <strong>of</strong> both<br />
fossil and extant species <strong>of</strong> that family. At present the<br />
<strong>Aegidae</strong> is considered to be not known from the fossil<br />
record.<br />
present I regard the relationships <strong>of</strong> families within the<br />
Cymothooidea and Cirolanoidea as equivocal.<br />
Until this present revision, Aega was a large genus,<br />
comprising some 00 species including new species<br />
described here. In the course <strong>of</strong> preparing this monograph<br />
it was apparent that there were several ‘species<br />
groups’ within Aega. <strong>The</strong>re is a large group <strong>of</strong> species<br />
related to Aegapheles deshaysiana (see Bruce 2004a) and<br />
a group <strong>of</strong> species related to Aega angustata and Aega<br />
komai; other perceived groups were those species with<br />
digitate pleopod margins (among other characters)<br />
such as Aegiochus coroo (Bruce, 983).<br />
Morphological observations suggested that there<br />
would be at least one major division within Aega sensu<br />
lato [as recognised by Brusca ( 983) when he established<br />
two subgenera], but it was subjectively entirely<br />
unclear to what extent other groups within Aega could<br />
be identified as monophyletic. <strong>The</strong> monophyly <strong>of</strong> Aega<br />
was assumed, although somewhat uncertain as most <strong>of</strong><br />
the distinguishing character states usually used to define<br />
or key the genus, such as the 5-articled maxilliped<br />
palp, would generally be regarded as plesiomorphic, as<br />
derived reductions <strong>of</strong> the number <strong>of</strong> articles to three or<br />
5
two are known in the superfamily Cymothooidea only<br />
from the parasitic Cymothoidae, ‘Epicaridea’ and the<br />
presumably more derived genera <strong>of</strong> the <strong>Aegidae</strong> such<br />
as Rocinela and Syscenus. <strong>The</strong> sole unique apomorphy<br />
upholding the monophyly <strong>of</strong> Aega (sensu lato) was<br />
maxilliped palp articles 3 and 4 with large recurved<br />
(i.e. strongly hooked) robust setae.<br />
Fifty-seven species were included in the analysis,<br />
being all those fully described for the purposes <strong>of</strong> this<br />
monograph and those described by Bruce (2004a) and<br />
yu and Bruce (2006). Specimens <strong>of</strong> Aega antennata,<br />
A. maxima, A. psora, A. serripes, Aegiochus arctica,<br />
Aegiochus plebeia, Aegiochus ventrosa, and Epulaega<br />
nodosa were examined and coded directly to the data<br />
set. Species coded from the literature were: Aegiochus<br />
francoisae (Wetzer, 990), Aegiochus lethrina (Bruce,<br />
983) (and one specimen), Aegiochus leptonica (Bruce,<br />
988); and Aega falcata Kensley and Chan, 200 . All<br />
other species were considered inadequately described<br />
for purposes <strong>of</strong> this analysis.<br />
Outgroup<br />
A preliminary analysis <strong>of</strong> the genera <strong>of</strong> <strong>Aegidae</strong>, using<br />
key characters as character states, was run using the<br />
phylogenetic analysis program PAUP* 4.0b 0 (Sw<strong>of</strong>ford<br />
2004) in order to assess potential sister-group<br />
relationships. Results indicated that Aega sensu lato<br />
was the sister group to all other genera, with Rocinela<br />
+ Alitropus the sister group to Syscenus + Xenuraega, a<br />
plausible working hypothesis. <strong>The</strong> final analysis was<br />
executed with Tridentella as the outgroup, because<br />
that family is the sister group to the clade <strong>Aegidae</strong>–<br />
Cymothoidae–‘Epicaridea’ <strong>of</strong> Brandt and Poore (2003).<br />
A particular coding difficulty was that within the<br />
<strong>Aegidae</strong> only Aega (sensu lato) has a 5-articled maxilliped<br />
palp, and so the different maxilliped states for<br />
palp articles 4 and 5 could not be coded against any<br />
aegid genus.<br />
cladistic analysis<br />
<strong>The</strong> data set used was derived from the descriptive<br />
DELTA character set, and modified to code all<br />
characters as unordered (i.e. reversible); multistate<br />
characters were treated as polymorphic. Most proportional<br />
characters were omitted; details <strong>of</strong> robust<br />
setae on the posterior pereopods were largely omitted.<br />
Nexus files were generated using DELTA. <strong>The</strong> data set<br />
consisted <strong>of</strong> 58 taxa (including single outgroup taxon)<br />
and 75 characters. <strong>The</strong> analyses used PAUP* (version<br />
4.0b. 0, Sw<strong>of</strong>ford 2004). A heuristic search was run using<br />
the treespace search method (hs addseq=random<br />
nchuck=3 chuckscore= nreps=500 randomize=trees).<br />
Resolution <strong>of</strong> the resultant trees was achieved through<br />
the use <strong>of</strong> the ‘reweight’ using the same constraints.<br />
6<br />
Parsimony jacknifing method in PAUP* was used to<br />
assess relative support for major clades.<br />
Characters are largely discussed in the section<br />
‘Morphology’ (p. 10), and specific states given in the<br />
character list.<br />
results<br />
A total 5,042 equally parsimonious trees <strong>of</strong> tree length<br />
608 was obtained, with a consistency index <strong>of</strong> 0.2 22,<br />
homoplasy index <strong>of</strong> 0.7878 and retention index <strong>of</strong><br />
0.66 7. <strong>The</strong> strict consensus tree (Fig. 4) and majority<br />
rule tree (Fig. 5) show two major clades, both with<br />
further resolution. <strong>The</strong> large number <strong>of</strong> trees generated<br />
is indicative <strong>of</strong> a high level <strong>of</strong> instability in the higher<br />
clades. <strong>The</strong> basal clades have a high level <strong>of</strong> stability<br />
with the three basal clades in the strict consensus (Fig.<br />
4) tree having jacknife support values <strong>of</strong> 98% (Aega +<br />
Aegapheles) and 83% (Aegiochus) 70% (Epulaega) respectively.<br />
<strong>The</strong> 50% majority rule tree (Fig. 4) shows that<br />
the major groupings are maintained, with significant<br />
further structure. Use <strong>of</strong> the reweight method <strong>of</strong> PAUP*<br />
resulted in a single fully resolved tree (Fig. 6), and the<br />
discussion <strong>of</strong> the clades focuses on the basal branches,<br />
the more terminal branches showing considerable<br />
instability.<br />
discussion <strong>of</strong> clades<br />
<strong>The</strong>re is a basal division (clades and 2) <strong>of</strong> all species<br />
<strong>of</strong> Aega sensu lato that confirms the division recognised<br />
by Brusca ( 983). <strong>The</strong>se two clades each split into<br />
two clades that are here recognised as Aega (clade 4),<br />
Aegapheles gen. nov. (clade 3), Aegiochus (clade 5) and<br />
Epulaega gen. nov. (clade 6).<br />
clade 1 is supported by four apomorphic states:<br />
the rostrum projecting anteriorly (Ch 3.2), antennule<br />
peduncle dorsoventrally flattened and expanded variously<br />
from weakly to strongly (Ch 8.2.3.4), maxilliped<br />
palp article 5 is wide (Ch 37.3) and pleopod endopod<br />
is subtruncate (Ch 53.2—with homoplasious occurrence<br />
in Aegiochus vigilans and Epulaega fracta). Most<br />
species in this clade lack a mandibular molar process<br />
(Ch 29.2) and have a relatively short antennule flagellum<br />
(Ch 2 .2).<br />
clade 2 has three defining apomorphies: the distal<br />
margin <strong>of</strong> the maxillule has three large robust setae and<br />
several small robust setae (Ch 30.3) with one homoplasious<br />
occurrence in Aega magnifica) and the principal<br />
robust setae are broad-based (Ch 3 .2), and the<br />
uropodal rami are acute (Ch 67.3); most species have a<br />
straight lateral margin <strong>of</strong> the uropodal exopod.<br />
clade 3 comprises the species here placed in the<br />
Aegapheles gen. nov., the defining apomorphies being<br />
the elongate point to the pleotelson apex (Ch 6.4)<br />
which also extends beyond the distal extremity <strong>of</strong> the<br />
uropodal rami. <strong>The</strong> uropodal rami are not coplanar (Ch
Figure 4. Clades in Aega: Strict consensus tree.<br />
7
64.2) with the plane <strong>of</strong> the exopod held at an oblique<br />
angle to the endopod (unique within clade , but with<br />
homoplasious occurrence with Rocinela). <strong>The</strong> uropodal<br />
endopod distolateral margin has 4 to 9 robust setae (Ch<br />
73.3). In addition the uropodal endopod lateral margin<br />
is excised, varying from weakly to strongly or ‘falcate’<br />
Figure 5. Clades in Aega: 50% majority rule; jacknife values.<br />
8<br />
(Ch 69 2.3), but this character state occurs in clade<br />
(see discussion for clade ).<br />
clade 4 comprises the species here considered<br />
to belong to Aega sensu lato, the characterising<br />
apomorphies being the frontal lamina posterior<br />
margin not clearly defined (Ch 26.2) and having sub-
parallel lateral margins (Ch 28.2) or widest posteriorly<br />
(Ch 28.4). <strong>The</strong> former has some homoplasious<br />
occurrence in Aegapheles, namely in<br />
A. excisa, A. mahana sp. nov. and A. umpara. <strong>The</strong> latter<br />
character is reversed in the clade Aega antennata–<br />
Aega falcata. Clade 4 divides into three clades (clades<br />
8, 9 and 0), and these are discussed in detail below;<br />
see also ‘general remarks’ (below).<br />
clade 5 constitutes Aegiochus sensu lato, and is<br />
upheld by a single apomorphic character state—a<br />
serrate pleotelson posterior margin (Ch 5.3). Additional<br />
states are the carpus <strong>of</strong> pereopods 2 and 3 with<br />
or 2 large robust setae (Ch 46.2), and most species<br />
have an acute appendix masculina (Ch 58.2) although<br />
there are five apparent reversals, possibly owing to the<br />
males not being fully mature.<br />
Within clade 5 there is the single-species clade<br />
<strong>of</strong> Aegiochus vigilans, sister clade to Aegiochus. This<br />
clade could warrant the establishment <strong>of</strong> a monotypic<br />
genus for A. vigilans. <strong>The</strong> species shows the apparent<br />
retention <strong>of</strong> some cirolanid-like characters, such as<br />
the morphology <strong>of</strong> the frontal lamina and clypeus, a<br />
tridentate mandibular incisor and a relatively large molar<br />
process that has marginal teeth. <strong>The</strong> mature males<br />
develop three large processes, one being the rostrum,<br />
two arising from the anterior margin <strong>of</strong> pereonite .<br />
This character in conjunction with the extremely long<br />
appendix masculina set on a posteromesial lobe are two<br />
unique states within the family, but it is far from certain<br />
that they be considered as <strong>of</strong> generic level (rather than<br />
species-level). Cephalic and pereopodal processes in<br />
related families such as the Cirolanidae and Corallanidae<br />
have not generally proved to be <strong>of</strong> generic merit,<br />
and the appendix masculina is <strong>of</strong>ten <strong>of</strong> variable length<br />
within genera; these two characters are considered too<br />
weak to use as reliable generic apomorphies within<br />
the family <strong>Aegidae</strong>. <strong>The</strong>re are numerous undescribed<br />
species <strong>of</strong> <strong>Aegidae</strong>, and the resolution <strong>of</strong> both major<br />
clades could change with further data; at this point a<br />
new genus is not proposed.<br />
clade 6 is Epulaega gen. nov., which is upheld by<br />
maxilliped palp article 5 being minute (Ch 37.4) and<br />
fused penial processes (Ch 49.3).<br />
clade 7 is Aegiochus sensu strictu, excluding<br />
A. vigilans, and is upheld by the rostrum separating the<br />
antennule bases in dorsal view (Ch 5.2) and maxilliped<br />
palp article 5 is subrectangular and longer than wide<br />
(Ch 37.2). In this clade the rostrum is either ventrally<br />
directed (Ch 3. —A. bertrandi–A. coroo clade) or ventrally<br />
directed and posteriorly folded (Ch 3.3—all other<br />
species). All species within this clade have one small<br />
robust seta on the inferior margin <strong>of</strong> the merus.<br />
clade 8 (and 9 and 10) includes the species here<br />
considered as Aega sensu strictu (excluding the species<br />
<strong>of</strong> clade ). <strong>The</strong> clade is upheld by one state, that <strong>of</strong><br />
antennule peduncle article 3 being less than half as<br />
wide as article 2 (Ch 20.2). Clade 9 lacks explicit apomorphies,<br />
but within this group there are two welldefined<br />
species pairs, each <strong>of</strong> which represent several<br />
more described and undescribed species. <strong>The</strong> clade<br />
A. angustata–A. komai is highly distinctive, and supported<br />
by several apomorphic states, these being the<br />
distal longitudinal carina on the pleotelson (Ch 2.3),<br />
the deeply serrate pleotelson posterior margin (Ch<br />
5.4), pereopod merus thickened (with one homoplasious<br />
occurrence in A. falcata) (Ch 39.2) and deeply<br />
serrate uropod margins (Ch 74.3); all species also have<br />
short, flat penial lobes, although this is not unique.<br />
One other named species (Aega d<strong>of</strong>leini) and several<br />
undescribed species belong to this clade on the basis<br />
<strong>of</strong> these recognised apomorphies. <strong>The</strong> clade A. antennata–A.<br />
falcata is supported by the unique antennule<br />
morphology, with peduncle article being strongly<br />
anteriorly produced (Ch 9.2) and the rostrum not<br />
separating antennule bases (Ch 5. ) (the antennule<br />
bases are divergent). <strong>The</strong>re are at least a further three<br />
undescribed species that belong to this clade. <strong>The</strong> sister<br />
clade to the A. antennata–A. falcata clade is the A.<br />
semicarinata–urotoma–angustata–komai clade, supported<br />
by the very large robust setae that oppose the dactylus<br />
<strong>of</strong> pereopod 2 or 2 and 3 (Ch 47.2).<br />
clade 10 is weakly characterised, with only homoplasious<br />
states (Ch 2.2, 8.3, and 58.2).<br />
clade 11 is potentially unstable—in all trees except<br />
for the reweighted tree (Fig. 6) clade clades with<br />
clade 3 (Aegapheles; see Figs 3–5). <strong>The</strong> character state<br />
that is shared with clade 3 is the falcate or excised lateral<br />
margin <strong>of</strong> the uropodal endopod. <strong>The</strong> species in<br />
clade lack the produced pleotelson apex (Ch 6.4),<br />
the posterior margin forming a caudomedial point<br />
(Ch 6.2); the uropodal rami are coplanar (Ch 64. );<br />
and there are few robust setae on the uropodal endopod<br />
lateral margin (Ch 72. , 73. ). In the reweighted<br />
tree these species clade as the sister group to Aega.<br />
On present data I regard the generic placement <strong>of</strong> the<br />
species in clade as equivocal.<br />
general remarks<br />
<strong>The</strong>re is strong support for the basal clades, and for<br />
the genera here recognised. This analysis can be regarded<br />
only as a first assessment, based on a relatively<br />
limited data set. It is possible that with description <strong>of</strong><br />
additional species within clade 4 (Aega sensu lato) a<br />
greater clarification <strong>of</strong> relationships within that clade<br />
may be achieved. In particular, the highly distinctive<br />
clades A. antennata–A. falcata and A. angusta–A. komai<br />
each have several undescribed species. Further data<br />
may allow more confident resolution <strong>of</strong> the position<br />
<strong>of</strong> clade . Further resolution <strong>of</strong> clade 5 may also allow<br />
for the establishment <strong>of</strong> a new genus for Aegiochus<br />
vigilans (no similar but undescribed species exist to my<br />
knowledge), although the unity <strong>of</strong> clade 7 (Aegiochus)<br />
9
Figure 6. Clades in Aega: successively weighted; type species in bold.<br />
20
CharaCter List for AegA<br />
body characters<br />
. Dorsal surfaces: . smooth or polished; 2. punctuate;<br />
3. heavily pitted.<br />
2. Lateral margins: . ovate; 2. subparallel.<br />
3. Rostral point: . ventrally directed, not projecting, not folded;<br />
2. projecting anteriorly, not ventrally folded; 3. folded<br />
ventrally and posteriorly.<br />
4. Rostral point: . minute; 2. prominent.<br />
5. Rostral point: . does not separate antennule bases (in dorsal<br />
view); 2. separates bases (in dorsal view).<br />
6. Eyes: . small (separated by more than 40% width <strong>of</strong><br />
head); 2. large, not medially united; 3. large, medially<br />
united.<br />
7. Pereonite 1 and coxae 2–3 each with posteroventral angle:<br />
. without produced point; 2. with small distinct<br />
produced point.<br />
8. Coxae 5–7, posterior margins: . convex; 2. straight;<br />
3. concave; 4. sinuate.<br />
9. Coxae 5–7, posterolateral angle: . acute (less than 45°);<br />
2. blunt (more than 45°); 3. rounded.<br />
0. Pleonite 4 posterolateral margins: . not extending to<br />
posterior margin <strong>of</strong> pleonite 5; 2. extending to but not<br />
beyond posterior margin <strong>of</strong> pleonite 5; 3. extending<br />
clearly beyond posterior margin <strong>of</strong> pleonite 5.<br />
. Pleonite 5 posterolateral angles: . overlapped by lateral<br />
margins <strong>of</strong> pleonite 4; 2. free, not overlapped by lateral<br />
margins <strong>of</strong> pleonite 4.<br />
2. Pleotelson dorsal surface: . with longitudinal carina<br />
only distally; 2. without longitudinal carina; 3. with<br />
longitudinal carina.<br />
3. Pleotelson dorsal surface: . without submedian depressions;<br />
2. anteriorly with 2 submedian depressions;<br />
3. posteriorly with 2 submedian depressions.<br />
4. Pleotelson lateral margins: . convex; 2. straight; 3. sinuate.<br />
5. Pleotelson marginal ornamentation: . smooth; 2. crenulated;<br />
3. serrate; 4. deeply serrate.<br />
6. Pleotelson posterior margin: . evenly rounded; 2. converging<br />
to caudomedial point; 3. sub-truncate (including<br />
emarginate); 4. with elongate medial point; 5. with<br />
median excision.<br />
7. Pleotelson, maximal robust setae: . without RS; 2. with 2<br />
to 6 RS; 3. with 7 to 0 RS; 4. with to 4 RS; 5. with<br />
5 or more RS.<br />
Antennule and antenna<br />
8. Antennule peduncle articles 1 and 2: . slender, cylindrical,<br />
article 2 without distal lobe; 2. flattened, article 2 without<br />
anterodistal lobe or weak lobe; 3. flattened, article<br />
2 anterodistal lobe not extending beyond mid-point <strong>of</strong><br />
article 3; 4. flattened, article 2 anterodistal lobe extending<br />
to end <strong>of</strong> article 3.<br />
9. Antennule peduncle articles 1 and 2: . not anteriorly produced;<br />
2. anteriorly produced.<br />
20. Antennule peduncle article 3: . more than half as wide as<br />
article 2; 2. less than half as wide as article 2.<br />
2 . Antennule flagellum: . ‘long’, extending posteriorly<br />
beyond head; 2. ‘short’, not extending posteriorly<br />
beyond head.<br />
2<br />
22. Antenna peduncle article 2 inferior surface: . without<br />
longitudinal suture; 2. with indistinct groove; 3. with<br />
distinct longitudinal suture.<br />
23. Antenna peduncle article 4: . without deep longitudinal<br />
groove; 2. with deep longitudinal groove.<br />
24. Antenna peduncle article 5: 1. not markedly wider or flatter<br />
than article 4; 2. flattened and expanded.<br />
25. Antenna flagellum: . ‘long’, extending beyond pereonite<br />
2; 2. ‘short’, not extending beyond pereonite 2.<br />
Frontal lamina and mouthparts<br />
26. Frontal lamina: . slender, reduced or absent; 2. posterior<br />
margin not clearly defined; 3. posterior margin clearly<br />
defined; 4. posterior margin free, forming a projecting<br />
‘blade’.<br />
27. Frontal lamina posterior margin: . posteriorly abutting<br />
clypeus; 2. not abutting clypeus; 2. with narrow posterior<br />
stem.<br />
28. Frontal lamina lateral margins: . diverging towards anterior;<br />
2. sub-parallel; 3. narrowing posteriorly; 4. widest<br />
posteriorly.<br />
29. Mandible molar process: 1. present, small distinct flat lobe;<br />
2. absent.<br />
30. Maxillule with: . several (6–8) distally hooked robust<br />
setae progressively increasing in size laterally;<br />
2. large and several (3–5) small straight<br />
or weakly hooked robust setae; 3. 3 large and<br />
several (3–5) small robust setae.<br />
3 . Maxillule principal RS: . narrow-based, slender, distally<br />
hooked; 2. wide-based, broad, distally acute or weakly<br />
hooked.<br />
32. Maxilla mesial lobe setae: . robust seta; 2. 2 robust setae;<br />
3. 3 robust setae; 4. 4 robust setae; 5. 5 robust setae.<br />
33. Maxilla mesial lobe setae: . simple; 2. both simple and serrate;<br />
3. serrate or plumose.<br />
34. Maxilliped article 3 robust setae: . narrow-based, elongate,<br />
straight or weakly curved; 2. broad-based, hooked.<br />
35. Maxilliped palp article 4 hooked RS: . all large or becoming<br />
progressively larger distally; 2. penultimate RS<br />
distinctly smaller than adjacent RS.<br />
36. Maxilliped palp article 5: . articulating with article 4; 2.<br />
partly fused to article 4; 3. wholly fused to article 4.<br />
37. Maxilliped palp article 5 shape: . longer than wide, distally<br />
rounded ; 2. longer than wide, sub-rectangular<br />
; 3. wider than long, distally<br />
convex; 4. small subcircular lobe.<br />
38. Maxilliped palp article 5 robust setae: . serrate (or simple<br />
and serrate), elongate, appearing flexible; 2. simple,<br />
stiff, weakly curved or straight.<br />
pereopods<br />
39. Pereopod 1 merus inferior margin: . not convex and thickened;<br />
2. convex and thickened.<br />
40. Pereopod 1 merus inferior margin: . with robust setae; 2.<br />
without robust setae.<br />
4 . Pereopods 2 and 3, merus inferior margin: . with large RS;<br />
2. with small RS; 3. RS absent.<br />
42. Pereopod 1 carpus inferodistal angle: . with RS; 2. without<br />
RS.<br />
43. Pereopod 1 (2 and 3) propodus inferior margin (palm): . with<br />
or more RS; 2. without RS.
44. Pereopod 1 propodal palm: . simple, without blade or distal<br />
lobe; 2. with small digitate distal lobe (no RS; rounded<br />
in sections); 3. with inferodistal margin produced (with<br />
RS); 4. with flat blade or broad lobe.<br />
45. Pereopod 2 merus inferior margin RS set as: . two (or three)<br />
discontinuous groups; 2. single row or rows.<br />
46. Pereopods 2 and 3 carpus inferodistal angle: . without RS or<br />
single small RS; 2. with or 2 large RS.<br />
47. Pereopod 2 or 3 propodus: . without large club-shaped<br />
distal robust seta; 2. with large club-shaped distal<br />
robust seta.<br />
48. Pereopods 5–7 inferior margins <strong>of</strong> ischium–carpus: . with<br />
short robust setae; 2. with long acute robust setae.<br />
penes<br />
49. Penial processes or openings: . set apart; 2. mutually adjacent;<br />
3. fused or united.<br />
50. Penes: 1. low tubercles; 2. opening flush with surface <strong>of</strong><br />
sternite 7; 3. short lobes.<br />
pleopods<br />
5 . Pleopod 1 exopod distally: . broadly rounded; 2. narrowly<br />
rounded, mesial margin weakly to strongly<br />
oblique.<br />
52. Pleopod 1 exopod mesial margin with PMS: . on distal onethird;<br />
2. on distal half; 3. on distal two-thirds; 4. on<br />
entire margin.<br />
53. Pleopod 1 endopod distally: . rounded; 2. subtruncate.<br />
54. Pleopod 1 endopod lateral margin with PMS from: . on distal<br />
margin only; 2. distal one-third; 3. distal half.<br />
55. Pleopod 1 endopod mesial margin with PMS on: . distal<br />
one-third; 2. distal half; 3. distal two-thirds; 4. entire<br />
margin; 5. distal margin only.<br />
56. Pleopod 2 appendix masculina: . with straight margins; 2.<br />
basally swollen.<br />
57. Pleopod 2 appendix masculina: . extending to or beyond<br />
distal margin <strong>of</strong> ramus; 2. not extending to distal<br />
margin <strong>of</strong> ramus.<br />
58. Pleopod 2 appendix masculina: . distally bluntly or narrowly<br />
rounded; 2. distally acute; 3. distally obliquely<br />
truncate.<br />
59. Appendix masculina: . without acute cuticular scales; 2.<br />
with acute cuticular scales.<br />
60. Exopods <strong>of</strong> pleopods 1–3 each with distolateral margin: . not<br />
digitate; 2. digitate.<br />
6 . Endopods <strong>of</strong> pleopods 3–5 each: . without distolateral point;<br />
2. with distolateral point.<br />
62. Pleopods 2–5 peduncle distolateral margin: . without prominent<br />
acute RS; 2. with prominent acute RS.<br />
uropods<br />
63. Uropod peduncle posterior lobe about: . ‘short’ one-third as<br />
long as endopod; 2. one-half to two-thirds as long as<br />
endopod; 3. ‘long’ two-thirds or longer than endopod<br />
length.<br />
64. Uropod rami: . with endopod and exopod co-planar;<br />
2. not co-planar, exopod at angle <strong>of</strong> about 35° to<br />
endopod.<br />
22<br />
65. Uropod rami: . extending to pleotelson apex; 2. not<br />
extending beyond pleotelson; 3. extending beyond<br />
pleotelson.<br />
66. Uropod rami marginal setae: . in single tier; 2. in two or<br />
three tiers; 3. dense, in several tiers.<br />
67. Uropod rami apices: . narrowly rounded; 2. broadly<br />
rounded; 3. acute.<br />
68. Uropod rami apically: 1. not bifid; 2. bifid.<br />
69. Uropod endopod lateral margin: . without prominent excision;<br />
2. falcate; 3. with prominent excision.<br />
70. Uropod endopod lateral proximal margin: . convex; 2.<br />
straight.<br />
7 . Uropod endopod lateral distal margin: . convex; 2. straight;<br />
3. concave.<br />
72. Uropod endopod proximal lateral margin with: . 0 or robust<br />
setae; 2. 2 to 6 (or more) robust setae.<br />
73. Uropod endopod distal lateral margin with: . 0 or robust<br />
setae; 2. 2 or 3 robust setae; 3. 4 to 9 robust setae.<br />
74. Uropod endopod mesial margin: . even, weakly or strongly<br />
convex; 2. sinuate; 3. deeply serrate.<br />
75. Uropod exopod: . not extending to end <strong>of</strong> endopod; 2.<br />
extending to end <strong>of</strong> endopod; 3. extending beyond<br />
end <strong>of</strong> endopod.<br />
ANAlysIs OF AEgIdAE<br />
<strong>The</strong> <strong>Aegidae</strong> White, 850, is a long established family,<br />
the unity <strong>of</strong> which has rarely been questioned. Brandt<br />
and Poore (2003, p. 898) rightly mention that ‘though<br />
these families are relatively easily recognisable, undisputed<br />
synapomorphies are not revealed in the literature’. Wägele<br />
( 989, fig. 93) suggested that the family might be<br />
paraphyletic, referring to a ‘Gruppe Aega’ (consisting<br />
solely <strong>of</strong> the genus Aega), that being the sister group<br />
to a clade containing ‘Gruppe Rocinela’ together<br />
with the Cymothoidae and the Epicaridea. Brusca<br />
and Wilson ( 99 ) disagreed with that interpretation,<br />
which had been based on the reduction <strong>of</strong> the articles<br />
<strong>of</strong> the maxilliped palp, considering such reductions<br />
as a common homoplasious adaptation to parasitism.<br />
Brusca and Wilson’s analysis and matrix equally failed<br />
to identify synapomorphies to uphold the <strong>Aegidae</strong> (I<br />
accept that this was not their intention), the <strong>Aegidae</strong><br />
coding identically to the Cymothoidae in that analysis.<br />
In the more recent analysis <strong>of</strong> Brandt and Poore (2003),<br />
the only apomorphic state identified that separates<br />
the <strong>Aegidae</strong> from the Cymothoidae is the presence<br />
<strong>of</strong> marginal setae on both rami <strong>of</strong> pleopods 3 and 4;<br />
this is a relatively weak character given that this is the<br />
state for the large families Cirolanidae, Corallanidae<br />
and also the Tridentellidae, and also that the <strong>Aegidae</strong><br />
is polymorphic for that character, with many species<br />
having the setae on endopods <strong>of</strong> pleopods 3 and 4<br />
either reduced or absent. Loss <strong>of</strong> marginal setae on<br />
pleopods 3 and 4 is a highly homoplasious character<br />
in the Cymothoidae associated with both freshwater<br />
habitats (Cirolanidae) and commensal or symbiotic life<br />
history (other families).
Traditionally the <strong>Aegidae</strong> have been characterised<br />
in keys and diagnoses as having ‘prehensile’ pereopods<br />
–3, or pereopods –3 with hooked dactylus, and having<br />
‘hooks’ or hooked ‘spines’ on the maxilliped palp<br />
(e.g. Bruce 993b; Kensley & Schotte 989; Wetzer &<br />
Brusca 997), although Bruce ( 993b) also referred<br />
to the bilobed maxilla with a small mesial lobe. This<br />
character, the maxilla being a simple broad plate with<br />
a distomesial lobe, is unique to the <strong>Aegidae</strong> and Cymothoidae,<br />
with a single homoplasious occurrence in<br />
the sphaeromatoid genus Paravireia Chilton, 925 (see<br />
Brökeland et al. 200 ). <strong>The</strong> <strong>Aegidae</strong>, Cymothoidae and<br />
Tridentellidae also share an elongate maxillule that is<br />
terminated by prominent, flat, incisory robust setae,<br />
these <strong>of</strong>ten being referred to as hooked though that is<br />
rarely the case.<br />
What then does uphold the monophyly <strong>of</strong> the<br />
<strong>Aegidae</strong>? <strong>The</strong> Cymothoidae have prehensile dactyli<br />
on pereopods –7, but ambulatory pereopods 4–7<br />
would generally be regarded as the plesiomorphic<br />
state within the Cymothoidae. A principal uniting<br />
character remains the maxilliped palp <strong>of</strong> five to three<br />
articles, with articles 2–4 provided with prominent<br />
and usually recurved robust setae. <strong>The</strong> Cymothoidae<br />
have maxilliped palp articles and 2 indistinguishably<br />
fused, and the axis <strong>of</strong> palp article 2 is strongly oblique<br />
to article . In contrast the <strong>Aegidae</strong> have between 2<br />
and 5 maxilliped palp articles, with Syscenus being<br />
polymorphic with 2 or 3 maxilliped palp articles (see<br />
figures in Bruce 2005).<br />
Eye size was not a character considered in previous<br />
analyses (Wägele 989, Brusca & Wilson 99 , Brandt<br />
& Poore 2003), but the <strong>Aegidae</strong> with few exceptions<br />
have large eyes, in Aega and Aegiochus these <strong>of</strong>ten being<br />
imperceptibly united medially. Eye reduction or loss<br />
is a common convergent state among parasites, cave<br />
and groundwater crustaceans, and deep-sea fauna. Enlargement<br />
<strong>of</strong> the eyes is, in contrast, rare and cannot be<br />
dismissed as a convergent or homoplasious character<br />
state. Although this condition occurs in some species<br />
<strong>of</strong> Corallanidae (see Delaney 989) and Tridentellidae,<br />
it is most highly developed in the <strong>Aegidae</strong>, with those<br />
species with the smallest eyes having eyes considerably<br />
larger than, for example, those <strong>of</strong> cirolanids or<br />
sphaeromatids.<br />
<strong>The</strong> character states that support the monophyly<br />
<strong>of</strong> the <strong>Aegidae</strong> are therefore the unique large eye size,<br />
in conjunction with a styliform maxillule with mesial<br />
robust setae (only terminal robust setae in the Cymothoidae)<br />
and the maxilla having one basal endite. <strong>The</strong><br />
characters <strong>of</strong> ‘prehensile’ pereopods –3 and hooked<br />
robust setae on the maxilliped palp are accurate but not<br />
unique to the <strong>Aegidae</strong> (both states effectively occurring<br />
in the Cymothoidae).<br />
23<br />
results<br />
A heuristic search was conducted, all characters unordered.<br />
<strong>The</strong> data set consisted <strong>of</strong> nine taxa (including<br />
single outgroup taxon) and 30 characters. A single fully<br />
resolved tree (Fig. 7) resulted.<br />
Epulaega presents as the sister group to the remainder<br />
<strong>of</strong> the <strong>Aegidae</strong>, the genus being upheld by the<br />
vestigial maxilliped palp article 5 (Ch 23.3) and the<br />
autapomorphic fused penial processes; the remaining<br />
genera are defined by having a dorsal rostrum (Ch 3.2),<br />
and separate into two clades, the Aegiochus–Aega–Aegapheles<br />
clade, which is supported by a large and acute<br />
rostrum (Ch 5.3) and the Alitropus—Xenuraega clade,<br />
upheld by numerous apomorphic states, notably the<br />
unicuspid (or absent) mandible incisor (Ch 7.3), lack<br />
<strong>of</strong> maxilliped endite (Ch 20.2), maxilliped palp with<br />
articles and 2 indivisibly fused, consisting <strong>of</strong> two or<br />
three articles (Ch 2 .2/3) with the major terminal article<br />
oblique to the axis <strong>of</strong> the maxilliped basal article,<br />
sub-basal appendix masculina (Ch 26.2), endopods <strong>of</strong><br />
pleopods 3 and 4 smaller than exopods (Ch 27.2) and<br />
uropod rami distally rounded (Ch 29.2).<br />
Figure 7. Cladogram <strong>of</strong> the genera <strong>of</strong> <strong>Aegidae</strong> with<br />
Tridentalla as the outgroup.
<strong>The</strong> Aega–Aegapheles clade is upheld by the flattened<br />
antennule peduncle article (Ch 2.2), frontal lamina<br />
wide and posteriorly separate from the clypeus (Ch<br />
6.2), and the endopods <strong>of</strong> pleopods 3 and 4 with few<br />
setae at the distomesial angle only (Ch 28.2). Aegiochus<br />
is characterised by the quadrate maxilliped palp article<br />
5 (Ch 23.2) and the rostrum clearly separating the<br />
antennule bases (Ch 0.2).<br />
<strong>The</strong> Rocinela–Syscenus–Xenuraega clade is supported<br />
by antennule peduncle articles and 2 elongate (Ch<br />
.2), antenna peduncle article 5 longest (Ch 3.2),<br />
and the maxillule lacking small mesial robust setae<br />
(Ch 8.2). <strong>The</strong> clade Syscenus–Xenuraega is defined by<br />
the head not being laterally overlapped by pereonite<br />
(Ch 2.2), the pleon distinctly narrower than the pereon<br />
and pleonite 5 with the lateral margins entirely free<br />
(Ch 7.2).<br />
CharaCter List for aegidae<br />
. Body: . dorsally vaulted; 2. dorsally depressed.<br />
2. Head: . overlapped laterally by anterior angles <strong>of</strong> pereonite<br />
; 2. not overlapped laterally by anterior angles<br />
<strong>of</strong> pereonite .<br />
3. Rostral point size: . small, scarcely or not visible in dorsal<br />
view; 2. large, prominent in dorsal view.<br />
4. Rostral point position: . anteroventral; 2. dorsal.<br />
5. Rostral point apex shape: . narrowly rounded; 2. broadly<br />
rounded; 3. acute.<br />
6. Coxae 5–7: . as long or longer than respective pereonite;<br />
2. shorter than respective pereonite.<br />
7. Pleon: . not distinctly narrower than pereonite 7; 2. distinctly<br />
narrower than pereonite 7.<br />
8. Pleonite 5: . lateral margins largely or wholly overlapped<br />
by pleonite 4; 2. lateral margins free.<br />
9. Eyes: . normal in size ; 2. large, occupying<br />
50% or more <strong>of</strong> head; 3. absent.<br />
0. Antennules, peduncle article 1: . close set or together;<br />
2. separated by rostrum.<br />
. Antennule peduncle: , articles and 2 short ; 2, articles and 2 elongate .<br />
24<br />
2. Antennule peduncle articles 1 and 2: . cylindrical; 2. dorsoventrally<br />
flattened.<br />
3. Antennal peduncle: . articles –3 short, 4 and 5 longest;<br />
2. articles and 2 short, 5 longest.<br />
4. Antenna peduncle articles 4 or 4 and 5: . without long<br />
plumose setae; 2. with long plumose setae.<br />
5. Frontal lamina: . wide < 3 H long as wide to wider than<br />
long>; 2. slender, elongate.<br />
6. Frontal lamina: . posterior margin wide, against<br />
clypeus; 2. posteriorly wide, separate from clypeus;<br />
3. posteriorly narrow, forming stem.<br />
7. Mandible incisor: . tricuspid; 2. biscuspid; 3. unicuspid;<br />
4. lacking incisor.<br />
8. Maxillule: . with small and large mesial robust setae;<br />
2. without small mesial robust setae.<br />
9. Maxillule distal setae: . slender only; 2. broad-based triangular<br />
.<br />
20. Maxilliped endite: . present; 2. absent.<br />
2 . Maxilliped palp articles: . 5-articled; 2. 3-articled;<br />
3. 2-articled.<br />
22. Maxilliped palp articles 3 and 4: . without hooked RS;<br />
2. with hooked RS.<br />
23. Maxilliped palp article 5: . rectangular, longer than wide;<br />
2. distally rounded, wider than long; 3. vestigial, short<br />
lobe.<br />
24. Maxilliped palp: . article present; 2. article indivisibly<br />
fused.<br />
25. Penial processes: 1. flat lobes; 2. flush .<br />
26. Pleopod 2 appendix masculina: . inserted basally; 2. inserted<br />
sub-basally.<br />
27. Pleopods 3 and 4 endopods: . same size as exopod; 2. smaller<br />
than exopod.<br />
28. Pleopods 3 and 4 endopods: . with PMS; 2. with few PMS<br />
at distolateral angle only; 3. without PMS.<br />
29. Uropodal rami: . with distinct apex; 2. rounded, without<br />
distinct apex.<br />
30. Uropod rami : . coplanar; 2. exopod at oblique<br />
angle to endopod.
subOrdEr cymOthOIdA wägElE,<br />
1989<br />
Brandt and Poore (2003) provided a new classification<br />
for the non-asellotan isopods (the ‘former Flabellifera’)<br />
based on a thorough character analysis, and recognised<br />
the subordinal separation <strong>of</strong>, among others, the Cymothoida<br />
Wägele, 989 from the Sphaeromatidea Wägele,<br />
989, as had earlier been proposed by Wägele ( 989).<br />
That classification is followed here.<br />
rEvAlIdAtION ANd dIAgNOsIs tO<br />
bArybrOtIdAE hANsEN, 1890<br />
barybrotidae Hansen, 890<br />
Barybrotidae Hansen, 890: 66.– Monod, 934: 0.<br />
diagnosis: Body evenly vaulted. Eyes dorso-lateral,<br />
large. Antennae and antennule well developed; division<br />
between peduncle and flagellum distinct; flagellae<br />
multi-articulate. Antennule shorter than antenna.<br />
Frontal lamina present, abutting clypeus; clypeus and<br />
labrum present. Mouthparts forming buccal cone.<br />
Mandible incisor broad, incisor tridentate; molar process<br />
present, lamellar; lacinia mobilis and spine row<br />
absent, represented by or 2 setae. Maxillule styliform,<br />
with flattened terminal RS. Maxilla a simple minute<br />
lobe, lacking RS. Maxilliped endite absent; palp with<br />
4 articles, article 2 elongate, about 2.9 times proximal<br />
width, articles 2–4 with hooked RS. Pereopods robust;<br />
pereopods –3 with prehensile dactylus, about as<br />
long or longer than propodus; superior distal angles<br />
<strong>of</strong> ischium and merus strongly produced and setose.<br />
Pereopods 4–7 ‘natatory’, with flattened basis, with superior<br />
and inferior margins provided with continuous<br />
row <strong>of</strong> long plumose setae. Pleon with 5 free pleonites<br />
plus pleotelson. Pleopod rami lamellar, without ridges<br />
or folding, with plumose marginal setae on both rami<br />
<strong>of</strong> pleopods and 2, setation reduced or absent on<br />
endopods <strong>of</strong> pleopods 3 and 4; pleopod 5 endopod<br />
without setae.<br />
Composition: <strong>The</strong> family has one monotypic genus<br />
Barybrotes Schioedte & Meinert, 879a, the type species<br />
<strong>of</strong> which is Barybrotes indus Schioedte & Meinert,<br />
879a; other named species are junior synonyms <strong>of</strong><br />
the type species.<br />
tAxONOmy<br />
25<br />
remarks: <strong>The</strong>re are several character states that prevent<br />
Barybrotes Schioedte & Meinert, 879a, being placed<br />
in the <strong>Aegidae</strong>, and that require the reinstatement <strong>of</strong><br />
Hansen’s ( 890) family. Prime among these is that the<br />
mouthparts, while reduced and probably used to feed<br />
from fish prey, do not show homologous character<br />
states with that <strong>of</strong> the <strong>Aegidae</strong>, nor the Corallanidae<br />
and Tridentellidae. In particular the maxilla is a minute<br />
single lobe lacking robust setae (similar to that seen in<br />
the Corallanidae), not wide and flat with a distomesial<br />
basal endite, and both maxilla lobes with hooked robust<br />
setae as occurs in all <strong>Aegidae</strong> and also Cymothoidae;<br />
the maxilliped is <strong>of</strong> a different form to that <strong>of</strong> the<br />
<strong>Aegidae</strong>, notably with only four palp articles, with<br />
article 2 elongate; and the mandible incisor retains<br />
the cirolanid form, being wide and tridentate, though<br />
somewhat narrower than seen in Cirolanidae. In the<br />
past the genus has been referred to the nominate family<br />
(e.g. Richardson 9 0; Thielemann 9 0; Monod 934)<br />
or subfamily (Nierstrasz 93 ), to the Corallanidae<br />
(Barnard 936) and more recently to the <strong>Aegidae</strong> (Pillai<br />
954, 967; Brandt & Poore 2003; Kensley et al. 2007). I<br />
have been unable to discover any published justification<br />
for placing Barybrotes in the <strong>Aegidae</strong>.<br />
<strong>The</strong>re are numerous character states that strongly<br />
suggest that Barybrotes has evolved from a Natatolanalike<br />
cirolanid ancestor (Natatolana Bruce, 98 ; see<br />
Keable 2006), including the proportions <strong>of</strong> the peduncular<br />
articles <strong>of</strong> the antennule (articles and 2 short,<br />
3 long) and antenna (articles 3 and 4 subequal in<br />
length), presence <strong>of</strong> a prominent pappose robust seta<br />
at the distal margin <strong>of</strong> antennular peduncle article 2,<br />
flagellum <strong>of</strong> the antennule with short (‘ring-like’) articles<br />
that may form a callynophore in males, elongate<br />
frontal lamina, wide and tridentate mandible incisor<br />
(though narrower than in the Cirolanidae), pereopods<br />
–3 with the superior distal angles <strong>of</strong> the ischium and<br />
merus produced and provided with long slender setae,<br />
pereopods 5–7 with a flattened basis provided with<br />
long plumose setae on superior and anterior margins<br />
and along the mid-lateral margin. All these character<br />
states are typical <strong>of</strong> Natatolana.<br />
<strong>The</strong> diagnosis is based on an examined series <strong>of</strong><br />
specimens from the Zoological Museum, Natural History<br />
Museum <strong>of</strong> Denmark, listed in Appendix 3.<br />
distribution: Indian Ocean from East Africa (present<br />
material) to Thailand; in the Pacific from Vietnam,<br />
Indonesia and Philippines.
<strong>Aegidae</strong> White, 850<br />
<strong>Aegidae</strong> White, 850: 68.– Dana, 852 : 304; 853: 765.–<br />
Hansen, 890: 3 5, 405.– Richardson, 905a: 66.–<br />
Menzies & Glynn, 968: 44.– Menzies & George, 972:<br />
9.9.– Kussakin, 979: 23 .– Brusca, 980: 229; 983:<br />
6.– Menzies & Kruczynski, 983: 6 .– Kensley & Schotte,<br />
989: 5.– Bruce, 993: 54.– Wetzer & Brusca, 997:<br />
30.– Roman & Dalens, 999: 228.– Bruce, Lew Ton<br />
& Poore, 2002: 59.– Keable, Poore & Wilson, 2002:<br />
unpaginated.<br />
Aeginae.– Menzies, 962: 7.<br />
diagnosis: Eyes large, <strong>of</strong>ten medially united. Mouthparts<br />
forming buccal cone; maxillule styliform, with<br />
terminal and mesial robust setae; maxilla with single<br />
distomesial basal endite; maxilliped palp with conspicuous<br />
recurved (‘hooked’) RS. Pereopods –3 robust,<br />
with dactylus as long as or longer than propodus,<br />
usually strongly recurved.<br />
desCription: Body evenly vaulted or dorsally depressed.<br />
Eyes lateral or dorso-lateral, usually large, sometimes<br />
contiguous or nearly so; occasionally absent. Antennae<br />
and antennule well developed; division between<br />
peduncle and flagellum distinct; flagellae multi-articulate;<br />
antennule shorter than antenna, peduncle<br />
4-articled; antennal peduncle 5-articled. Frontal lamina<br />
present, varied in shape, occasionally absent, usually<br />
not abutting clypeus; clypeus and labrum present,<br />
<strong>of</strong>ten indistinct. Mouthparts forming buccal cone.<br />
Mandible incisor narrow, small molar process present,<br />
occasionally absent, lamellar and triangular when<br />
present; lacinia mobilis and spine row absent. Maxillule<br />
styliform, with flattened terminal RS, may be distally<br />
hooked; mesial lobe present or absent. Maxilla with<br />
small distomesial basal endite joined to larger mesial<br />
lobe; each lobe with or more broad, usually apically<br />
curved (hooked) RS. Maxilliped endite present (Aega<br />
group <strong>of</strong> genera and Rocinela) or absent (Syscenus and<br />
Xenuraega); palp with 3–5 articles, at least articles 3 and<br />
4 with large hooked RS. Pereopods robust; pereopods<br />
–3 with strongly curved dactylus (i.e. prehensile),<br />
about as long or longer than propodus (occasionally<br />
weakly curved or shorter than propodus); with few<br />
slender setae. Pereopods 4–7 ambulatory, articles not<br />
compressed or flattened, basis without long plumose<br />
marginal setae; ischium to propodus inferior and distal<br />
margins with RS. Pleopod rami lamellar, without<br />
ridges or folding, with plumose marginal setae on both<br />
rami <strong>of</strong> pleopods and 2, setation variously reduced<br />
or absent on endopods <strong>of</strong> pleopods 3–5; pleopod 5<br />
endopod without setae.<br />
remarks: <strong>The</strong>re are few unique character states that can<br />
be used to define the <strong>Aegidae</strong> (see ‘Analysis’ p. 22).<br />
<strong>The</strong>re is a pagination error in this publication, with page<br />
304 printed as 204.<br />
26<br />
Within the Cymothoida the styliform maxillule, with<br />
terminal robust setae is a character shared with both the<br />
Tridentellidae and Cymothoidae, and a maxillule that<br />
has mesial (i.e. subterminal) as well as terminal robust<br />
setae is shared only with the Tridentellidae. <strong>The</strong> maxilla<br />
with a single distomesial endite is a character shared<br />
only with the Cymothoidae. <strong>The</strong> Tridentellidae have<br />
ambulatory pereopods –3 and lack hooked robust<br />
setae on the maxilliped, and are further characterised<br />
by having an elongate maxilliped endite. <strong>The</strong> <strong>Aegidae</strong><br />
is the only family in which all but a very few species<br />
have greatly enlarged eyes, a state that is in general<br />
rare within the <strong>Isopoda</strong>. <strong>The</strong>re are some Corallanidae<br />
that have large eyes, but in most species the eyes are<br />
similar in size to those <strong>of</strong> the Cirolanidae. <strong>The</strong> Corallanidae<br />
lack hooked robust setae on the maxilliped,<br />
pereopods –3 are ambulatory, and the family has the<br />
unique character states <strong>of</strong> strongly hooked maxillule<br />
and vestigial maxilla with no endites. <strong>The</strong> <strong>Aegidae</strong>,<br />
lack <strong>of</strong> wholly unique characters notwithstanding, can<br />
be readily identified by the combination <strong>of</strong> characters<br />
listed in the diagnosis.<br />
Key to the marine genera <strong>of</strong> aegidae<br />
A key to all genera was provided by Bruce ( 993a),<br />
which included the only estuarine and freshwater genus<br />
Alitropus (known only from tropical Australia and<br />
Asia). Regional keys have been given to the East Pacific<br />
by Brusca ( 983), the Caribbean (Kensley & Schotte<br />
989), and to northern cold-water seas by Kussakin<br />
( 979, in Russian).<br />
Although the marine genus Xenuraega Tattersall,<br />
909 has not been recorded from <strong>New</strong> <strong>Zealand</strong> it is<br />
included in this key. <strong>The</strong> true extent <strong>of</strong> the distribution<br />
<strong>of</strong> this genus is far from certain, but mesopelagic and<br />
pelagic isopods <strong>of</strong>ten have extensive distributions,<br />
sometimes in all oceans (e.g. Metacirolana caeca, see<br />
Svavarsson and Bruce 2000, and Aega monophthalma<br />
herein). For this reason it is considered entirely possible<br />
that Xenuraega could be taken in <strong>New</strong> <strong>Zealand</strong><br />
waters.<br />
Pleonite abruptly narrower than pereonite 7;<br />
pleonite 5 lateral margins entirely free; eyes usually<br />
absent ...............................................................2<br />
– Pleonite not abruptly narrower than pereonite 7;<br />
pleonite 5 lateral margins partly or entirely overlapped<br />
by pleonite 4; eyes present, <strong>of</strong>ten large .3<br />
2. Frontal lamina present; maxilliped palp 3-articled;<br />
both uropod rami lamellar ........Syscenus [p. 98]<br />
– Frontal lamina absent; maxilliped palp 2-articled;<br />
uropodal rami with endopod stub-like, exopod<br />
filamentous................................Xenuraega [p. 2 5]
3. Body dorsally compressed; frontal lamina slender,<br />
shield-shaped or lanceolate; rostrum anteriorly<br />
widely rounded or truncate; maxilliped palp 3articled<br />
...........................................Rocinela [p. 6 ]<br />
– Body dorsally moderately to strongly vaulted;<br />
frontal lamina wide; rostrum narrowly rounded<br />
or acute; maxilliped palp 5-articled .....................4<br />
4. Rostrum anteriorly directed, acute; frontal lamina<br />
ventrally flat, antennule peduncle articles 1 and<br />
2 flattened; maxilliped palp article 5 wider than<br />
long, partly or entirely fused to 4 ........................5<br />
– Frontal lamina with free posterior margin and/or<br />
posteriorly narrow; antennule peduncle articles<br />
1 and 2 not flattened or expanded .......................6<br />
5. Uropod rami co-planar; uropodal rami to or beyond<br />
pleotelson apex; uropodal endopod lateral<br />
margin without distinct excision; pleotelson lacking<br />
distinct, usually produced point ............ Aega<br />
– Plane <strong>of</strong> uropod endopod at oblique angle to<br />
exopod, uropodal rami not extending to pleotelson<br />
apex; uropodal endopod lateral margin<br />
usually distinct excision; pleotelson apex forming<br />
distinct, usually produced point ............................<br />
......................................................Aegapheles [p. 65]<br />
6. Rostrum bent ventrally or ventrally and posteriorly;<br />
maxilliped palp article 5 longer than wide,<br />
not fused to article 4 ...................Aegiochus [p. 83]<br />
– Rostrum minute, not projecting, not visible in<br />
dorsal view; maxilliped palp article 5 minute, less<br />
than 0.3 width <strong>of</strong> article 4...........Epulaega [p. 5 ]<br />
Genus Aega Leach, 8 5<br />
Æga Leach, 8 5: 369; 8 8: 549; Desmarest, 825.– Milne<br />
Edwards, 840: 238.– Dana, 852: 304*; 853: 747.–<br />
Gosse, 855: 34.– Harger, 880: 383.– Haswell, 882:<br />
284.– Bate & Westwood, 86 – 868: 276.– Miers, 876b:<br />
08.– Schioedte & Meinert, 879b: 334.– Hansen, 890:<br />
3 6.– Sars, 897: 58.– Richardson, 905a: 67.– Stebbing,<br />
905: 20.– Hodgson, 9 0: 7.– Stephensen, 948: 36.<br />
Aega.– Gerstaecker, 882: 227.– Barnard, 9 4: 36 ; 936:<br />
57.– Hale, 925: 68.– Wahrberg, 930: 8.– Nierstrasz<br />
& Schuurmans Stekhoven Jr, 930: e74.– Gurjanova,<br />
933: 429; 936: 70.– Holthuis, 956: 4 .– Menzies, 962:<br />
7.– Schultz, 969: 89.– Menzies & George, 972:<br />
7.– Kensley, 978: 56.– Kussakin, 979: 23 .– Brusca,<br />
983: 7.– Menzies & Kruczynski, 983: 62.– Bruce, 983:<br />
757; 996: 29.– Brusca & Iverson, 985: 40.– Kensley<br />
& Schotte, 989: 6.– Bruce, Lew Ton & Poore, 2002:<br />
60.<br />
Pterelas Guérin-Méneville, 836: VII.– Dana, 852: 204; Dana,<br />
853: 748.<br />
Æegacylla Dana, 854: 76.<br />
Aega (Aega).– Brusca, 983: 0.<br />
* <strong>The</strong>re is a pagination error in this publication, with page<br />
304 printed as 204.<br />
27<br />
type speCies: Oniscus psora Linnaeus, 758 (= Aega psora<br />
(Linnaeus, 758); original orthography was Oniscus<br />
Pforá; by subsequent designation, Menzies ( 962). Aega<br />
emarginata (Leach, 8 5) is a junior synonym. Aega affinis<br />
Milne Edwards, 840 was regarded as a synonym<br />
<strong>of</strong> A. psora by Kussakin ( 979).<br />
diagnosis: Body moderately to strongly dorsally<br />
vaulted. Rostral point acute, anteriorly produced between<br />
antennule peduncles. Eyes present, <strong>of</strong>ten large,<br />
usually separate. Pleon not distinctly narrower than<br />
pereonite 7, pleonite not abruptly narrower than<br />
pleonite 2. Antennule peduncle articles 1 and 2 flattened,<br />
<strong>of</strong>ten expanded with anterodistal angle <strong>of</strong> article<br />
2 forming lobe, article 3 less than 0.3 H width <strong>of</strong> article<br />
2. Frontal lamina wide, posterior margin not clearly<br />
defined, lateral margins usually straight. Maxilliped<br />
palp 5-articled; article 5 wide, <strong>of</strong>ten fused to article<br />
4, distal margin convex, with slender setae; endite<br />
present. Coxae 5–7 as long as or longer than respective<br />
pereonite. Pereopods –3 merus inferior margin with<br />
large robust setae, usually set as one or more rows.<br />
desCription: Pleon not abruptly narrower than pereon;<br />
pleonites all visible, not posteriorly widest, pleonite 5<br />
laterally overlapped by pleonite 4; pleonites 3–5 posteriorly<br />
produced to an acute point. Pleotelson large,<br />
about as long as longer pleon, usually with PMS and<br />
RS.<br />
Mandible with uni- or bicuspid incisor; molar<br />
process present, reduced or absent. Maxillule with 5–8<br />
elongate, flat, narrow-based terminal and mesial RS.<br />
Maxilliped 5-articled, article wider than long, articles<br />
3 and 4 each with 2–6 stout recurved RS, article 5 with<br />
2–7 occasionally hooked RS; endite present, usually<br />
with –2 terminal setae.<br />
remarks: Under the revised concept Aega sensu strictu<br />
contains those species with a prominent, acute and<br />
anteriorly projecting rostrum, the antennule peduncle<br />
with articles and 2 strongly dorsoventrally<br />
compressed, sometimes with an anterolateral lobe, a<br />
slender peduncle article 3 (less than one-third as wide<br />
as article 2), and the uropod peduncle with an elongate<br />
mesial lobe that stretches most <strong>of</strong> the length <strong>of</strong> the uropodal<br />
endopod. Species within Aega sensu strictu lack<br />
a falcate uropodal endopod, although this is weakly<br />
expressed in the type species; most species have matte,<br />
punctate or pitted dorsal body surfaces.<br />
Three species, A. magnifica (Dana, 854), A. maxima<br />
Hansen, 897 and A. sheni yu & Bruce, 2006 lack the<br />
slender antennule peduncle article 3 and have a clearly<br />
falcate uropodal endopod, and approach some Aegapheles<br />
in the appearance <strong>of</strong> the antennule and uropodal<br />
endopod. <strong>The</strong>se species are here regarded as incertae<br />
sedis (see discussion <strong>of</strong> clades, p. 6– 9).
Aega antennata Richardson, 9 0 and A. falcata<br />
Kensley & Chan, 200 are immediately distinguished<br />
from all other species (and all other <strong>Aegidae</strong>) by having<br />
antennule peduncle article strongly anteriorly<br />
produced. <strong>The</strong>re are several other undescribed species<br />
similar to these two species that also have this unique<br />
character state.<br />
Thirty-six named species are included in the genus,<br />
those below, and those listed under ‘Species included<br />
...’ (p.2 2). <strong>The</strong> genus is represented in all oceans<br />
from shallow waters to a depths <strong>of</strong> 2 48 metres (Aega<br />
maxima).<br />
etymoLogy: <strong>The</strong> name could be derived from Greek<br />
mythology (e.g. Aega being described as a nursemaid<br />
to Zeus, and variously as the daughter <strong>of</strong> Olenos, <strong>of</strong><br />
King Melisseus <strong>of</strong> Crete and <strong>of</strong> Helios). Alternatively<br />
the name could be derived from the Greek aeigis or the<br />
Latin aegis, meaning shield or cover. Another possibility<br />
is that the name was in allusion to the relatively<br />
large eyes <strong>of</strong> many species and is derived from Middle<br />
or Old English ēage. Leach ( 8 5) gave no clues<br />
as to his choice <strong>of</strong> name, and as it seems not to relate<br />
directly to mythological history, geographical location<br />
nor morphological attributes, the basis for his choice<br />
remains a mystery.<br />
Key to the new <strong>Zealand</strong> species <strong>of</strong> Aega<br />
Eyes large, medially united ..................................2<br />
– Eyes separate ..........................................................3<br />
2. Posterior margins <strong>of</strong> pereonites 6 and 7 and pleonites<br />
nodular; surfaces heavily pitted; pleotelson<br />
dorsally with distinct median longitudinal carina,<br />
posterior margin with distinct apical point; margins<br />
<strong>of</strong> pleotelson and uropods with conspicuous<br />
acute RS ....................... Aega monophthalma (p. 37)<br />
– Posterior margins <strong>of</strong> pereonites 6 and 7 and pleonites<br />
smooth; surfaces finely setose; pleotelson<br />
dorsally without median carina, posterior margin<br />
without distinct apical point; margins <strong>of</strong> pleotelson<br />
and uropods RS small ......................................<br />
..............................................Aega stevelowei (p. 50)<br />
3. Pereopod 2 or 2 and 3 propodus with large clubshaped<br />
RS opposite base <strong>of</strong> dactylus; uropodal<br />
endopod lateral margin even ...............................4<br />
– Pereopod 2 or 2 and 3 propodus without large<br />
club-shaped RS opposite base <strong>of</strong> dactylus; uropodal<br />
endopod lateral margin falcate .................6<br />
4. Pleotelson and uropods deeply serrate; body<br />
elongate, more than 3 times as long as greatest<br />
width ...........................................Aega komai (p. 34)<br />
– Pleotelson and uropods not deeply serrate; body<br />
less than 3 times as long as greatest width .........5<br />
28<br />
5. Pleotelson dorsally with two sub-median depressions,<br />
posterior margin strongly concave; eyes<br />
narrowly separated ( by ~9% width <strong>of</strong> head) ......<br />
..........................................Aega semicarinata (p. 44)<br />
– Pleotelson dorsally without depression, posterior<br />
margin subtruncate; eyes widely separated (by<br />
~29% width <strong>of</strong> head) .............Aega urotoma (p. 55)<br />
6. Body very wide ( .6 times as long as greatest<br />
width), dorsal surfaces distinctly pitted; eyes<br />
small, widely separated (by ~38% width <strong>of</strong><br />
head) pereopods –3 dactylus about as long as<br />
propodus ................................. Aega whanui (p. 6 )<br />
– Body wide ( .8 times as long as greatest width),<br />
dorsal surfaces smooth; eyes large, narrowly<br />
separated ( by ~ 0% width <strong>of</strong> head) pereopods<br />
–3 dactylus about .5 as long as propodus .........<br />
.............................................Aega falklandica (p. 28)<br />
Aega falklandica Kussakin, 967 (Figs 8– )<br />
Aega falklandica Kussakin, 1967: 227, figs 3, 4.– Kensley, 1980:<br />
159; 2001: 227.– Branch, Griffiths, Kensley & Sieg, 1991:<br />
12, fig. (not numbered).<br />
Aega (Aega) falklandica.– Brusca, 983: .<br />
materiaL examined: Holotype <strong>of</strong> Aega falklandica: ♀<br />
(non-ovig. 3 mm), <strong>New</strong> Island, Falkland Islands, 2<br />
April 959, 0 m, coll. Slava. Zool Inst, Acad. Science,<br />
Leningrad RAN /46405.<br />
Non-type. ♂ (3 mm), Macquarie Ridge, 54°30–28’S,<br />
59°00’E, 5 February 967, Cr 27, stn 975, 443–549 m,<br />
coll. RV Eltanin (USNM 099250).<br />
Also examined: Holotype <strong>of</strong> Aega maxima Hansen,<br />
897, ♀ (non-ovig. 54 mm), <strong>of</strong>f Cocos Island, <strong>of</strong>f Panama,<br />
East Pacific, 26 February 1891, Albatross stn 3362,<br />
2056 m [as 25 fms] (USNM 20727).<br />
desCription: Body .8 times as long as greatest width,<br />
dorsal surfaces smooth and sparsely punctate, widest<br />
at pereonite 5, lateral margins ovate. Eyes large,<br />
not medially united, separated by about 0% width<br />
<strong>of</strong> head; each eye made up <strong>of</strong> ~27 transverse rows <strong>of</strong><br />
ommatidia, each row with ~ 5– 7 ommatidia; eye<br />
colour dark brown. Pereonite 1 and coxae 2–3 each with<br />
posteroventral angle with small distinct produced<br />
point. Coxae 5–7 with entire oblique carina; posterior<br />
margins sinuate, posterolateral angle acute (less than<br />
45°). Pleon with pleonite visible in dorsal view; pleonite<br />
4 with posterolateral margins extending clearly<br />
beyond posterior margin <strong>of</strong> pleonite 5; pleonite 5 with<br />
posterolateral angles overlapped by lateral margins <strong>of</strong><br />
pleonite 4. Pleotelson 0.8 times as long as anterior width,<br />
dorsal surface without longitudinal carina; lateral margins<br />
sinuate, smooth, posterior margin converging to<br />
caudomedial point, with 6–8 RS.
Figure 8. Aega falklandica Kussakin, 967. Holotype; all appendages drawn in situ (Leningrad, RN /46405). A, dorsal view,<br />
holotype; B, lateral view; C, head; D, frons; E, pereopod , distal articles; F, pereopod 2, distal articles; G, pleotelson posterior<br />
margin apex; H, uropod endopod, ventral view; I, left uropod, dorsal view.<br />
29
Figure 9. Aega falklandica Kussakin, 967. Eltanin specimen (USNM 099250). A, lateral view; B, antenna peduncle;<br />
C, antennule; D, mandible; E, mandible palp article 3; F, maxillule; G, maxillule apex; H, maxilla; I, maxilla apex;<br />
J, maxilliped; K, maxilliped palp; L, maxilliped palp article 5 (Leica); peduncle, dorsal view.<br />
30
Figure 10. Aega falklandica Kussakin, 967. Eltanin specimen (USNM 099250). A–D, pereopods , 2, 6 and 7 respectively;<br />
E, pereopod ischium, mesial surface; F, sternite 7 showing penial papillae.<br />
Antennule peduncle article 2 anterodistal lobe not<br />
extending beyond mid-point <strong>of</strong> article 3; articles 3 and<br />
4 0.5 times as long as combined lengths <strong>of</strong> articles and<br />
2, article 3 2.6 times as long as wide; flagellum with 12<br />
articles, extending to posterior margin <strong>of</strong> eye. Antenna<br />
peduncle article 2 inferior surface without distinct longitudinal<br />
suture; article 4 .6 times as long as wide, 0.9<br />
times as long as combined lengths <strong>of</strong> articles –3, with<br />
deep longitudinal groove, inferior margin plumose<br />
seta, and 0 short slender setae; article 5 not markedly<br />
wider or flatter than article 4, 1.1 times as long as article<br />
4, 2.2 times as long as wide, inferior margin with 6<br />
palmate setae; flagellum with 17 articles, extending to<br />
middle <strong>of</strong> pereonite .<br />
3<br />
Frontal lamina flat, as wide as long, lateral margins<br />
converging posteriorly, anterior margin rounded, with<br />
small median point, posterior margin not abutting<br />
clypeus.<br />
Mandible molar process present, minute; palp article<br />
2 with 7 distolateral setae (3 large biserrate, remainder<br />
smaller, simple), palp article 3 with 27 setae. Maxillule<br />
with 8 terminal RS (falcate). Maxilla mesial lobe with<br />
5 RS (3 stout, 2 distally biserrate); lateral lobe with 3<br />
RS. Maxilliped endite with 0 apical setae; palp article 2<br />
with 6 RS (1 hooked; with further fine marginal setae);<br />
article 3 with 6 recurved RS (5 hooked, long straight);<br />
article 4 with 7 hooked RS (5 large, 2 small); article 5
articulating with article 4, distally convex, with 6 RS<br />
(5 straight, curved).<br />
Pereopod 1 basis .9 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 2 RS (and simple<br />
seta); merus inferior margin with RS (or 2), set as<br />
distal group, superior distal angle with 0 RS (2 slender<br />
setae); carpus 0.5 as long as merus, inferior margin with<br />
0 RS; propodus .3 times as long as proximal width,<br />
inferior margin with 0 RS, propodal palm with small<br />
distal lobe, dactylus abruptly hooked, .5 as long as<br />
propodus. Pereopod 2 ischium inferior margin with<br />
RS, superior distal margin with 3 RS (and slender<br />
seta); merus inferior margin with 6 RS (set as 4 + 2), set<br />
as two groups, superior distal margin with 0 acute RS<br />
(4 slender setae); carpus longer than that <strong>of</strong> pereopod<br />
, with inferodistal lobe, inferodistal angle with 0 RS,<br />
propodus without large club-shaped distal RS. Pereopod<br />
3 similar to pereopod 2 (7 or 9 RS); propodus without<br />
large club-shaped distal RS. Pereopods 5–7 inferior<br />
margins <strong>of</strong> ischium–carpus with short RS. Pereopod 6<br />
similar to pereopod 7 (slightly larger, inferior margins<br />
with more RS). Pereopod 7 basis 3.3 times as long as<br />
greatest width, inferior margins with 5 palmate setae<br />
(or more); ischium 0.5 as long as basis, inferior margin<br />
with 9 RS (set as , 2, 3 and 3), superior distal angle with<br />
6 RS, inferior distal angle with 7 RS; merus 0.8 as long<br />
as ischium, .9 times as long as wide, inferior margin<br />
with 9 RS (set as , 4 and 4), superior distal angle with<br />
0 RS (and 2 slender setae), inferior distal angle with<br />
8 RS; carpus 0.8 as long as ischium, 2.6 times as long<br />
as wide, inferior margin with 7 RS (set as 3 and 4),<br />
superior distal angle with 0 RS, inferior distal angle<br />
with 0 RS; propodus 0.6 as long as ischium, 3.0 as<br />
long as wide, inferior margin with 5 RS (set as , 2 and<br />
2), superior distal angle with 2 slender setae, inferior<br />
distal angle with 3 RS.<br />
Penes low tubercles; penial openings separated by<br />
4% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod .9 times as long as wide, distally<br />
narrowly rounded, mesial margin weakly oblique,<br />
lateral margin straight, mesial margin strongly convex,<br />
with PMS on distal two-thirds; endopod 2.5 times<br />
as long as wide, distally subtruncate, lateral margin<br />
strongly concave, with PMS on distal margin only, mesial<br />
margin with PMS on distal half; peduncle .9 times<br />
as wide as long, mesial margin with 8 coupling hooks.<br />
Pleopod 2 appendix masculina with straight margins, 0.8<br />
times as long as endopod, distally narrowly rounded<br />
(with small apical point). Exopods <strong>of</strong> pleopods –3<br />
each with distolateral margin not digitate; endopods<br />
<strong>of</strong> pleopods 3–5 each with distolateral point; pleopods<br />
2–4 peduncle distolateral margin without prominent<br />
acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about three-quarters as long as endo-<br />
32<br />
pod. Uropod rami with endopod and exopod weakly<br />
oblique, rami extending to pleotelson apex, marginal<br />
setae in single tier (dense), apices narrowly rounded.<br />
Endopod apically not bifid, lateral margin proximally<br />
convex, with prominent excision (shallow), positioned<br />
about three-quarters along ramus, proximal lateral<br />
margin with RS, distal lateral margin with 3 RS,<br />
mesial margin weakly convex, with 6 RS. Exopod not<br />
extending to end <strong>of</strong> endopod, 3.3 times as long as<br />
greatest width, apically not bifid; lateral margin weakly<br />
convex, with 2 RS; mesial margin sinuate, proximally<br />
concave, with 3 RS.<br />
femaLe: Similar to the male, but for the sexual characters;<br />
no ovigerous females present.<br />
size: Present material 3 mm.<br />
Variation: <strong>The</strong> two specimens differ in a number <strong>of</strong><br />
details, though without more material it is impossible<br />
to say whether this is regional variation or potentially<br />
specific differences. <strong>The</strong> robust setae on the merus <strong>of</strong><br />
pereopods –3 present a constant pattern, although the<br />
number <strong>of</strong> robust setae varied with the holotype having<br />
only RS on the pereopod merus and the <strong>New</strong><br />
<strong>Zealand</strong> specimen having 2, the merus <strong>of</strong> pereopod 2<br />
has 4+2 but pereopod 3 merus had 5+2 (holotype) or<br />
6+3 (<strong>New</strong> <strong>Zealand</strong>).<br />
<strong>The</strong> shape and proportions <strong>of</strong> the uropod are the<br />
same between the two specimens but there is a difference<br />
in the number <strong>of</strong> robust setae, notably on the<br />
uropodal endopod lateral margin with the holotype<br />
having a pattern <strong>of</strong> +3, the <strong>New</strong> <strong>Zealand</strong> specimen<br />
+2. Both specimens had somewhat damaged uropods<br />
so these numbers may be an artefact <strong>of</strong> that damage.<br />
remarks: Aega falklandica can be identified by the antennule<br />
peduncle articles 1 and 2 being flattened and<br />
expanded, uropodal endopod lateral margin being<br />
medially indented with the anterior portion conspicuously<br />
convex, by the short propodus with a simple<br />
palm and small distal lobe, and by the pattern and<br />
number <strong>of</strong> robust setae on pereopods –3. Similar species<br />
include Aega magnifica which is readily separated<br />
by pereopods –3 having a conspicuous blade on the<br />
palm <strong>of</strong> the propodus.<br />
<strong>The</strong>re are two similar Pacific species: Aega acuminata<br />
Hansen 897 and Aega maxima Hansen, 897. <strong>The</strong><br />
former has far smaller eyes than A. falklandica, the propodal<br />
lobe on pereopod is larger and the uropodal<br />
exopod is proportionally longer, extending just beyond<br />
the apex <strong>of</strong> the endopod (Brusca 983; Hansen 897).<br />
<strong>The</strong> principle differences between A. falklandica and<br />
A. maxima are, in A. maxima, slightly smaller eyes, the<br />
palm <strong>of</strong> pereopods –3 without any trace <strong>of</strong> a distal<br />
lobe, and the uropodal endopod distal margin appear-
Figure 11. Aega falklandica Kussakin, 967. Eltanin specimen (USNM 099250). A–E, pleopods –5 respectively; F, pleopod<br />
peduncle mesial margin; G, uropod; H, uropod exopod, ventral view.<br />
ing distinctly truncate and with only a weak excision on<br />
the lateral margin. <strong>The</strong> only known specimen <strong>of</strong> Aega<br />
maxima was taken at a depth 2 48 metres <strong>of</strong>f Cocos<br />
Island, <strong>of</strong>f Pacific Panama (Hansen 1897), considerably<br />
deeper that the holotype <strong>of</strong> A. falklandica ( 0 m) or the<br />
<strong>New</strong> <strong>Zealand</strong> specimen (maximum depth <strong>of</strong> 549 m).<br />
Kensley’s records are from 85 to 270 m.<br />
33<br />
prey: No records.<br />
distribution: Falkland Islands, South Atlantic, Marion<br />
Island, southern Indian Ocean and <strong>of</strong>f southwestern<br />
<strong>New</strong> <strong>Zealand</strong>. At depths <strong>of</strong> 0 m (Falkland Islands)<br />
otherwise 85–270 m (Marion Islands) and 549 metres<br />
(<strong>New</strong> <strong>Zealand</strong>).
Aega komai Bruce, 996 (Figs 2, 3)<br />
Aega komai Bruce, 1996: 129, figs 1–4.<br />
Aega angustata.— Stephenson, 1980: 153, figs 1–5.<br />
[misidentification, not Aega angustata Whitelegge,<br />
90 ].<br />
materiaL: ♀ (non-ovig., 26 mm), <strong>of</strong>f Taranaki Bight<br />
region, 39°02.5’S, 73°55.5’E, 2 March 990, 86 m, on<br />
spiny dogfish snout, coll. J.B. Jones (NMNZ Cr.12000).<br />
2 ♂ ( 6.5, 8.0 mm), between Fannel Island and Barrier<br />
Island, Hauraki Gulf, 22 November 976, 86–97 m,<br />
Squalus blainvillei, <strong>of</strong>f skin behind pectoral fins, coll.<br />
RV Ikatere (AK 4855).<br />
Additional material: ♀ (non-ovig., 23 mm), Taiwan,<br />
22° 8.6’N, 9° 4.8’E, 28 July 2000, stn CP , 262 m,<br />
coll. Bouchet, Richer de Forges and Chan (MNHN<br />
Is.5860). ♂ (29 mm), <strong>of</strong>f Great Barrier Island, North<br />
Island, January 2006, old longline gear at ~500 m, coll.<br />
Steve Lowe (NIWA 23777).<br />
Also examined: Holotype <strong>of</strong> Aega angustata Whitelegge,<br />
90 . ♂ ( 4.3 mm), 5.5–6.5 km <strong>of</strong>f Wattamolla, NSW,<br />
34° 0’S, 5 ° ’E, 22 March 898, stn 57, 08 m, coll.<br />
E.R. Waite on HMCS <strong>The</strong>tis (AM G2 60).<br />
desCription <strong>of</strong> new zeaLand speCimens: Body 3.4 times<br />
as long as greatest width, dorsal surfaces polished<br />
in appearance, widest at pereonite 5, lateral margins<br />
subparallel. Rostral point projecting anteriorly, not<br />
ventrally folded. Eyes large, not medially united, separated<br />
by about 36% width <strong>of</strong> head; each eye made up<br />
<strong>of</strong> ~ 6 transverse rows <strong>of</strong> ommatidia, each row with ~9<br />
ommatidia; eye colour pale brown. Pereonite 1 and coxae<br />
2–3 each with posteroventral angle with small distinct<br />
produced point (ventral); coxae 5–7 with incomplete<br />
oblique carina. Pleon with pleonite visible in dorsal<br />
view; pleonite 4 with posterolateral margins extending<br />
to but not beyond posterior margin <strong>of</strong> pleonite 5; pleonite<br />
5 with posterolateral angles free, not overlapped<br />
by lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.7 times as<br />
long as anterior width, dorsal surface with longitudinal<br />
carina on distal third; lateral margins weakly convex,<br />
deeply serrate (with 13–15 flat marginal spines), posterior<br />
margin subtruncate, with 3– 5 RS.<br />
Antennule peduncle article 2 anterodistal lobe extending<br />
to end <strong>of</strong> article 3; article 3 0.3 times as long as<br />
combined lengths <strong>of</strong> articles and 2, 3.0 as long as wide;<br />
flagellum with 6 articles, extending to mid-point <strong>of</strong><br />
eye. Antenna peduncle article 2 inferior surface without<br />
distinct longitudinal suture; article 4 0.8 times as long as<br />
wide, 0.8 times as long as combined lengths <strong>of</strong> articles<br />
–3, with deep longitudinal groove, inferior margin 0<br />
plumose setae, and 0 short simple setae; article 5 flattened<br />
and expanded, 2.4 times as long as article 4, .7<br />
times as long as wide, inferior margin with 2 pappose<br />
setae, anterodistal angle with cluster <strong>of</strong> 5 short simple<br />
34<br />
setae; flagellum with 9 articles, extending to posterior<br />
<strong>of</strong> pereonite .<br />
Frontal lamina flat, as wide as long, lateral margins<br />
converging posteriorly, anterior margin rounded,<br />
forming median angle, posterior margin not abutting<br />
clypeus.<br />
Pereopod 1 basis .4 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin<br />
with 0 RS, superior distal margin with 2 RS (acute);<br />
merus inferior margin convex and thickened, with 0<br />
RS, superior distal angle with 2 RS; carpus 0.9 as long<br />
as merus; inferior margin with 0 RS; propodus .4<br />
times as long as proximal width, inferior margin with<br />
0 RS, propodal palm with small distal lobe (concave),<br />
dactylus abruptly hooked, .0 as long as propodus.<br />
Pereopod 2 ischium inferior margin with RS (stout),<br />
superior distal margin with 2 RS; merus inferior margin<br />
with 6 RS (and 2 slender setae), set as two groups (ill-<br />
defined), superior distal margin with 2 acute RS; carpus<br />
longer than that <strong>of</strong> pereopod , with inferodistal lobe,<br />
inferodistal angle with RS, propodus without large<br />
club-shaped distal robust seta. Pereopod 3 not similar to<br />
pereopod 2 (dactylus slender and claw-like); propodus<br />
with large club-shaped distal robust seta. Pereopod 6<br />
similar to pereopod 7 (but more robust with longer RS<br />
on inferior margins). Pereopod 7 basis 3.2 times as long<br />
as greatest width, inferior margins with 6 palmate setae<br />
(many missing); ischium 0.7 as long as basis, inferior<br />
margin with 9 RS (set loosely as , 4 and 4), superior<br />
distal angle with 5 RS, inferior distal angle with 0<br />
RS; merus 0.7 as long as ischium, 2.7 times as long as<br />
wide, inferior margin with 9 RS (set loosely as , 4 and<br />
4), superior distal angle with 6 RS, inferior distal angle<br />
with 6 RS; carpus 0.9 as long as ischium, 5.3 times as<br />
long as wide, inferior margin with 9 RS (set as 2, 2, , ,<br />
2 and ), superior distal angle with 3 RS, inferior distal<br />
angle with 7 RS; propodus 0.8 as long as ischium, 7.4<br />
times as long as wide, inferior margin with 5 RS (set<br />
as , , 2 and ), superior distal angle with slender<br />
setae, inferior distal angle with 4 RS.<br />
Penes low tubercles; penial openings separated by<br />
0% <strong>of</strong> sternal width.<br />
Uropod peduncle ventrolateral margin with 2 RS<br />
(and ~5 plumose setae), posterior lobe about threequarters<br />
as long as endopod. Uropod rami extending<br />
beyond pleotelson (lateral and mesial margins deeply<br />
serrate), marginal setae in single tier, apices acute.<br />
Endopod apically not bifid, lateral margin straight<br />
(deeply serrate), without prominent excision, proximal<br />
lateral margin with 5 RS (margin not divided, with 5<br />
prominent flat spines), mesial margin straight (deeply<br />
serrate), with 2 RS (and 2 prominent flat spines). Exopod<br />
not extending to end <strong>of</strong> endopod, 3.2 times as long<br />
as greatest width, apically not bifid; lateral margin<br />
straight (deeply serrate), with 5 RS (and 5 prominent<br />
flat spines); mesial margin straight, with 1 RS.
Figure 12. Aega komai Bruce, 996. NMNZ female, except F and I. A, dorsal view; B, lateral view; C, head; D, frons;<br />
E, pleonites, lateral view; F, penial process; G, pleotelson and uropods; H, apex <strong>of</strong> pleotelson; I, sternite 7; J, antenna<br />
peduncle, showing deep groove.<br />
35
Figure 13. Aega komai Bruce, 996. NMNZ female. A–D, pereopods –3 and 7 respectively (pereopods 2 and 3 basis<br />
omitted); E, pereopod 2, mesial surface <strong>of</strong> ischium; F, dactylus, pereopod 3; G, antennule; H, antenna; I, uropod; J, uropodal<br />
exopod, ventral view.<br />
36
size: Female 26 mm, two males 6.5 and 8 mm; male<br />
holotype (Japan) 20.5 mm.<br />
Variation: <strong>The</strong>re are only three specimens, and therefore<br />
the details here are <strong>of</strong> range only. Pleotelson RS<br />
3– 5, with parallel variation in the spines. Uropod<br />
endopod mesial margin 2 RS ( once), lateral margin<br />
always 5 RS; uropod exopod mesial margin , lateral<br />
margin 6–8 RS.<br />
Pereopod always without RS; inferior margin <strong>of</strong><br />
merus <strong>of</strong> pereopods 2 and 3 each with 5–7 RS (these<br />
robust setae are difficult to observe without dissection,<br />
so the range may be narrower or greater than<br />
given here).<br />
<strong>The</strong> material from <strong>New</strong> <strong>Zealand</strong> agrees well with<br />
the description <strong>of</strong> the holotype, but there are a number<br />
<strong>of</strong> small differences. <strong>The</strong> lateral margin <strong>of</strong> the uropodal<br />
exopod in the holotype has a shorter proportion<br />
<strong>of</strong> the lateral margin serrate (56% <strong>of</strong> the length <strong>of</strong> the<br />
ramus) than the <strong>New</strong> <strong>Zealand</strong> material (64% <strong>of</strong> length<br />
<strong>of</strong> ramus). <strong>The</strong> holotype has partly damaged uropods,<br />
and without additional material is not possible to be<br />
certain if this is a consistent difference between the two<br />
populations. <strong>The</strong> proportions and setation <strong>of</strong> pereopod<br />
7 also vary slightly, suggesting the possibility that these<br />
are separate populations.<br />
remarks: Aega komai is can be identified by the following<br />
combination <strong>of</strong> characters: elongate body, antennule<br />
peduncle articles and 2 strongly compressed and<br />
expanded, antenna article 5 conspicuously flattened,<br />
deeply serrate uropod rami which extend beyond the<br />
posterior margin <strong>of</strong> the pleotelson and a deeply serrate<br />
subtruncate pleotelson posterior margin.<br />
<strong>The</strong>re are three other similar species: Aega angustata<br />
Whitelegge, 90 , Aega d<strong>of</strong>leini Thielemann, 920,<br />
and an undescribed species from southern Australian<br />
waters. Aega angustata is readily distinguished by the<br />
produced pleotelson posterior margin, the uropod rami<br />
falling well short <strong>of</strong> the pleotelson posterior margin and<br />
the uropod exopod lateral margin not being serrate and<br />
provided with prominent robust setae on both margins.<br />
Aega d<strong>of</strong>leini has a produced pleotelson margin, with<br />
uropodal rami extending to the pleotelson apex; and<br />
the pleotelson and uropodal rami are figured as being<br />
weakly and irregularly serrate. <strong>The</strong> as-yet-undescribed<br />
species from southern Australia has the uropods extending<br />
to the pleotelson apex and the posterior margins<br />
<strong>of</strong> the pleotelson are distinctly angled and with<br />
smaller serrations and spines than A. komai.<br />
prey: <strong>The</strong> holotype was recorded from the mantle <strong>of</strong><br />
the squid Loligo bleekeri Keferstein, although this may<br />
be a capture artefact. Squalus blainvillei (Risso, 827),<br />
Squalidae; dogfish and longnose spurdog (UK usage),<br />
grey-spiny or spiny dogfish (<strong>New</strong> <strong>Zealand</strong> usage).<br />
37<br />
distribution: Previously recorded from Japan. In <strong>New</strong><br />
<strong>Zealand</strong> from Taranaki Bight and Hauraki Gulf, western<br />
and northeastern North Island respectively; also<br />
Taiwan; at depths <strong>of</strong> 86–262 metres.<br />
Aega monophthalma Johnston, 834 (Figs 4– 8)<br />
Æga monophthalma Johnston, 1834: 233, fig. 43a–b.– Milne<br />
Edwards, 840: 244.– Lütken, 859: 75.– Bate &<br />
Westwood, 1867*: 286, figure.– Sars, 1897: 62, pl. 26, fig.<br />
.– Norman, 904: 434; 905a: 94; 905b: 3.– Hansen,<br />
1916: 171.– Stephensen, 1948: 38, fig. 7 (8–9).<br />
Rocinela monophthalma.– White, 1850: 80; 1857: 253, pl. 14, fig.<br />
7.– Gosse, 1855: 134, fig. 233.<br />
Æga monopthalma.– Schioedte & Meinert, 879b: 365<br />
(lapsus).<br />
Aega monophthalma.– Gerstaecker, 882: 254.– Barnard, 9 4:<br />
362, pl. 3 B– Nierstrasz & Schuurmans Stekhoven, 930:<br />
77, fig. 14.– Nierstrasz, 93 : 83– Gurjanova, 933:<br />
430.– Stephensen, 1937: 7, 17.– Kussakin, 1979: 235: figs<br />
04, 05.– Ellis, 98 : 23.– Hemmingsen & MacKenzie,<br />
996: 37; 200 : 9.– Bruce, 200 : 2, photo.– Bruce,<br />
Lew Ton & Poore, 2002: 6 .– Tracey et al., 2005: 07,<br />
colour fig.<br />
Aega monopthalma.– Moreira & Sadowsky, 979: 08.— Treat,<br />
1980: 912, fig. 1 (lapsus).<br />
Aega monopthalmus.– Kensley, 1978: 57, fig. 24G–H; 2001:<br />
227.<br />
Aega (Aega) monophthalma.– Brusca, 983: .<br />
type LoCaLity: “Berwick on Tweed” (Johnston 834),<br />
Berwick Bay, Northumberland. Johnston had two specimens<br />
and two species, and stated that he described the<br />
larger specimen which is now in <strong>The</strong> Natural History<br />
Museum, London (holotype, BMNH 979:299: ). <strong>The</strong><br />
smaller specimen is Aega stroemii Lütken, 859 [= A.<br />
bicarinata Rathke, 837, not A. bicarinata Leach, 8 8,<br />
according to Brusca ( 983)].<br />
materiaL examined: Holotype, ♂ (50 mm), Berwick on<br />
Tweed, Northumberland, on large codfish, White MS<br />
cat. No. 972a, coll. G. Johnston (BMNH 979.299. )<br />
[penes close set but apart and not projecting.].<br />
Non-type. <strong>New</strong> <strong>Zealand</strong>: ♂ (49 mm), Chatham Rise,<br />
42°45.68’S, 79°59.33’W, 2 April 200 , 920–77 m, coll.<br />
RV Tangaroa (NIWA 23755). ♀ (non-ovig. 62 mm), <strong>New</strong><br />
<strong>Zealand</strong>, in fish pound after fish processed, Fisheries<br />
Research Division stn CO2/ 02/88 (NIWA 23756).<br />
♀ (ovig. 63 mm), South Norfolk Ridge, 33°22.6 ’S,<br />
70° 2.70’E, June 2003, 5 4–540 m, coll. NORFANZ,<br />
RV Tangaroa (NIWA 23757). Immature (28 mm),<br />
manca ( 8.5 mm), North Norfolk Ridge, 28°5 .2 ’S,<br />
67°42.53’E, 5 May 2003, 690–8 2 m, coll. NORFANZ,<br />
RV Tangaroa (NIWA 23758, 23759). Australia: ♂ (40<br />
mm), ♀ (48 mm), 758–84 m, east <strong>of</strong> Kiama, NSW,<br />
34°42–38’S, 5 ° 6– 8’E, 3 December 987, 760–855 m,<br />
* See Holthuis ( 977) for details <strong>of</strong> the dates <strong>of</strong> publication<br />
<strong>of</strong> Bate and Westwood’s book.
Figure 14. Aega monophthalma Johnston, 834. NIWA 23755. A, dorsal view; B, lateral view; C, head; D, frons; E, anterior<br />
view <strong>of</strong> frontal lamina; F, penial openings; G, antennule, dorsal view; H, antennule, ventral view; I, antenna, dorsal view;<br />
J, antenna, ventral view; K, pleotelson and uropods.<br />
38
Figure 15. Aega monophthalma Johnston, 834. NIWA 23755. A, mandible; B, mandible palp article 3; C, maxillule;<br />
D, maxillule apex; E, maxilla; F, maxilla apex; G, maxilliped; H, maxilliped palp articles 4 and 5 (Leica); I, maxilliped palp<br />
articles –5.<br />
39
coll. FRV Kapala (AM P43978). & (56 mm), <strong>of</strong>f Broken<br />
Bay, NSW, 33°26–29’S, 52°06–04’E, 5 July 980, 440<br />
m, coll. FRV Kapala (AM P3 9 8).<br />
Additional material. <strong>New</strong> <strong>Zealand</strong> region: ♀ (nonovig<br />
72 mm), <strong>of</strong>f Great Barrier Island, North Island,<br />
24 April 2004, from Hyperoglyphe antarctica, longline<br />
at ~500 m, coll. Steve Lowe (NIWA 23760). ♀ (nonovig<br />
47 mm), <strong>of</strong>f Great Barrier Island, North Island,<br />
October–November 2004, from Hyperoglyphe antarctica,<br />
longline at ~500 m, coll. Steve Lowe (NIWA 2376 ).<br />
♀ (non-ovig 63 mm), vicinity <strong>of</strong> West Norfolk Rise,<br />
33°4 ’S, 67° 4’E, 600 m, FV Jacquiline, stn 69, coll.<br />
D. Smith (NMNZ Cr. 335). ♀ (non-ovig 73 mm),<br />
Chatham Rise, 44°32. 9–34.04’S, 75°27.95–27.94’W,<br />
3 November 2005, 880– 022 m, commercial trawl,<br />
stn SWA050 /70, coll. P. McMillan (NIWA 23762).<br />
<strong>New</strong> caledonia: Manca (24 mm), 23° 9’S, 68°00’E, 2<br />
October 992, BERyx , stn. CP60, 590–600 m, coll. B.<br />
Richer de Forges (MNHN Is.5861).<br />
desCription: Body .9 times as long as greatest width,<br />
dorsal surfaces punctate (coarsely pitted, posterior<br />
margins <strong>of</strong> pleonites with coarse granules approaching<br />
nodular), widest at pereonite 6, lateral margins weakly<br />
ovate. Rostral point projecting anteriorly, not ventrally<br />
folded. Eyes large, medially united, anterior clear field<br />
8% length <strong>of</strong> head, posterior clear field 33% length<br />
<strong>of</strong> head; each eye made up <strong>of</strong> ~36 transverse rows <strong>of</strong><br />
ommatidia, each row with ~ 8 ommatidia; eye colour<br />
black. Pereonite 1 and coxae 2–3 each with postero-<br />
ventral angle right-angled (small produced point;<br />
coxae 3 and 4 posteriorly rounded). Coxae 5–7 with<br />
entire oblique carina; posterior margins straight, posterolateral<br />
angle rounded. Pleon with pleonite largely<br />
concealed by pereonite 7; pleonite 4 with posterolateral<br />
margins extending to but not beyond posterior margin<br />
<strong>of</strong> pleonite 5; pleonite 5 with posterolateral angles<br />
overlapped by lateral margins <strong>of</strong> pleonite 4. Pleotelson<br />
0.6 times as long as anterior width (with deep reticulated<br />
pits), dorsal surface with longitudinal carina and<br />
with 2 sub-medial depressions; lateral margins weakly<br />
convex, smooth, posterior margin subtruncate or with<br />
distinct short median point (somewhat sinuate), with<br />
44–48 RS.<br />
Antennule peduncle article 2 anterodistal lobe extending<br />
to end <strong>of</strong> article 3; articles 3 and 4 0.25 times<br />
as long as combined lengths <strong>of</strong> articles and 2, article<br />
3 2.3 times as long as wide; flagellum with 12 articles,<br />
extending to mid-point <strong>of</strong> eye. Antenna peduncle article<br />
2 inferior surface with distinct longitudinal suture;<br />
article 4 .5 times as long as wide, 0.8 times as long<br />
as combined lengths <strong>of</strong> articles –3, with deep longitudinal<br />
groove, inferior margin plumose setae, and<br />
2 short simple setae; article 5 not markedly wider or<br />
flatter than article 4, 1.5 times as long as article 4, 2.3<br />
times as long as wide, inferior margin with palmate<br />
40<br />
seta (distal), anterodistal angle with cluster <strong>of</strong> five short<br />
simple setae; flagellum with 22 articles, extending to<br />
posterior <strong>of</strong> pereonite .<br />
Frontal lamina flat, longer than greatest width,<br />
rectangular (lateral margins weakly concave, ridged),<br />
anterior margin with median point (downwardly directed,<br />
anteriorly recessed process), with prominent<br />
median point, posterior margin abutting clypeus.<br />
Mandible molar process present, minute; palp article<br />
2 with 8 distolateral setae. Maxillule with 8 terminal<br />
and subterminal RS. Maxilla mesial lobe with 4 RS (2<br />
hooked, 2 weakly curved); lateral lobe with 4 RS. Maxilliped<br />
endite with 2 apical setae (long weakly CP); palp<br />
article 2 with 8 RS (small stiff setae/slender RS); article<br />
3 with 6 recurved RS (and simple straight RS); article<br />
4 with 7 hooked RS (4 large, 3 small); article 5 partly<br />
fused to article 4, distally convex, with 5 RS (partly<br />
fused with article 4; all setae short and simple).<br />
Pereopod 1 basis .9 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 2 RS (and 2 slender<br />
simple setae); merus inferior margin with 3 RS, set as<br />
two groups (<strong>of</strong> , 2 and distal simple setae), superior<br />
distal angle with 0 RS (2 simple setae); carpus 0.6 as<br />
long as merus, inferior margin with 0 RS; propodus .<br />
times as long as proximal width, inferior margin with 0<br />
RS (distally with 2 small simple setae), propodal palm<br />
with small distal lobe, dactylus smoothly curved, .5 as<br />
long as propodus. Pereopod 2 ischium inferior margin<br />
with RS, superior distal margin with 2 RS (and 2<br />
simple setae); merus inferior margin with 5 RS (set as<br />
3 + 2 setae and distal simple seta), set as two groups,<br />
superior distal margin with acute RS; carpus similar<br />
in size to that <strong>of</strong> pereopod , inferodistal angle with<br />
RS, propodus without large club-shaped distal RS.<br />
Pereopod 3 similar to pereopod 2 (but longer, ischium<br />
inferior distal angle with 2 RS; dactylus markedly more<br />
slender than that <strong>of</strong> pereopods and 2); propodus without<br />
large club-shaped distal RS. Pereopods 5–7 inferior<br />
margins <strong>of</strong> ischium–carpus with short RS. Pereopod 6<br />
similar to pereopod 7 (with fewer RS on inferior margins<br />
<strong>of</strong> ischium–propodus). Pereopod 7 basis 3.2 times<br />
as long as greatest width, inferior margins with 0 palmate<br />
setae; ischium 0.6 as long as basis, inferior margin<br />
with 6 RS (set as , 2 and 3), superior distal angle with 6<br />
RS, inferior distal angle with 6 RS; merus 0.9 as long as<br />
ischium, 2. times as long as wide, inferior margin with<br />
6 RS (set as , 3 and 2), superior distal angle with<br />
RS, inferior distal angle with 9 RS; carpus 0.9 as long as<br />
ischium, 2.6 times as long as wide, inferior margin with<br />
5 RS (set as and 4), superior distal angle with 9 RS,<br />
inferior distal angle with 0 RS; propodus 0.6 as long<br />
as ischium, 3.5 times as long as wide, inferior margin<br />
with 4 RS (set as and 4), superior distal angle with 4<br />
slender setae ( acute RS and 2 simple and palmate<br />
setae), inferior distal angle with 3 RS.
Figure 16. Aega monophthalma Johnston, 834. NIWA 23755. A–E, pereopods –3, 6, 7 respectively; F and G, pereopod and<br />
2, ischium superior distal angle, mesial side.<br />
4
Figure 17. Aega monophthalma Johnston, 834. NIWA 23755. A–D, pleopods –3, 5 respectively; E, appendix masculina apex;<br />
F, uropod.<br />
Penes low tubercles; penial openings separated by<br />
7% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod .9 times as long as wide, distally<br />
narrowly rounded, mesial margin weakly oblique,<br />
lateral margin straight, mesial margin strongly convex<br />
(finely crenulate with minute simple setae present),<br />
with PMS on distal one-third; endopod 2.2 times as<br />
long as wide, distally subtruncate, lateral margin<br />
42<br />
strongly concave, with PMS on distal one-third, mesial<br />
margin with PMS on distal margin only; peduncle 2<br />
times as wide as long, mesial margin with coupling<br />
hooks. Pleopod 2 appendix masculina with straight margins,<br />
0.9 times as long as endopod, distally acute (with<br />
narrowed apical point; basally with lateral groove).<br />
Exopods <strong>of</strong> pleopods –3 each with distolateral margin<br />
not digitate; endopods <strong>of</strong> pleopods 3–5 each with dis-
Figure 18. Aega monophthalma Johnston, 834. NIWA 23755. A, uropod exopod, ventral view; B, uropod exopod, apex; C,<br />
uropod endopod, apex.<br />
tolateral point; pleopods 2–4 peduncle distolateral margin<br />
with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 3 RS<br />
(and continuous PMS), posterior lobe about as long as<br />
endopod. Uropod rami extending to pleotelson apex,<br />
marginal setae dense, in several tiers, apices broadly<br />
rounded. Endopod apically not bifid, lateral margin<br />
proximally convex, without prominent excision, proximal<br />
lateral margin with 3 RS, distal lateral margin<br />
with 2 RS, mesial margin strongly convex or concave,<br />
with 7– 0 RS. Exopod extending to end <strong>of</strong> endopod,<br />
2.9 times as long as greatest width, apically not bifid;<br />
lateral margin convex, with 7– 8 RS; with 3–4 RS.<br />
femaLe: Eyes narrowly separated; ovigerous female<br />
ocular surface depressed, dorsal body surfaces not as<br />
nodular as male; uropodal margins lacking prominent<br />
RS; non-ovigerous female similar to male with the<br />
exception <strong>of</strong> sexual characteristics.<br />
size: Specimens from the southwestern Pacific: males<br />
from 40 to 49 mm, females 48 to 73 mm; single manca<br />
8.5 mm.<br />
43<br />
Variation: Only a small number (5) <strong>of</strong> specimens were<br />
available at the time <strong>of</strong> writing the description for this<br />
species, and the fact that the uropod and pleotelson<br />
apices are mostly damaged means that it is not possible<br />
to precisely detail the variation present. <strong>The</strong> pleotelson<br />
has from about 44 to 48 (22+22 to 24+24) robust setae.<br />
<strong>The</strong> uropod endopod lateral margin has 2– 7 RS, the<br />
mesial margin 7– 0 RS; the exopod lateral margin has<br />
6– 8 RS, the mesial margin 3–5 RS. <strong>The</strong> robust setae<br />
on the merus <strong>of</strong> pereopods –3 are constant: pereopod<br />
merus with +2, pereopod 2 merus with 3+2 and<br />
pereopod 3 merus to 4+2.<br />
<strong>The</strong> extent to which the antennule peduncle articles<br />
and 2 are produced varies, with the large specimen<br />
from <strong>of</strong>f Great Barrier Island being less strongly produced;<br />
in some specimens the dorsal pitting is weaker<br />
than in others, and the robust setae on the uropods and<br />
pleotelson are not always as prominent as illustrated;<br />
uropod apices are also frequently damaged and regrowth<br />
may appear more rounded that in undamaged<br />
specimens.<br />
remarks: This spectacular and large isopod, at the time<br />
<strong>of</strong> first description only the seventh in the genus, is im-
mediately recognisable by the highly textured dorsal<br />
surface, heavily ‘spined’ pleotelson and uropods, the<br />
subtruncate pleotelson posterior margin with a welldefined<br />
median point, the pleotelson with a prominent<br />
longitudinal ridge, the antennule peduncle articles<br />
being flattened and expanded, the huge eyes which<br />
appear to be medially united and the characteristic<br />
shape <strong>of</strong> the frontal lamina. Juvenile specimens are not<br />
as nodular, and can be identified by the characteristic<br />
shape <strong>of</strong> the frontal lamina, and the shape, ornamentation<br />
and setation <strong>of</strong> the pleotelson and uropodal rami.<br />
<strong>The</strong> ovigerous female is slightly wider in body shape,<br />
and the prominent robust setae are missing from the<br />
margins <strong>of</strong> the pleotelson and uropods; the frontal<br />
lamina is the same as in the male, and the appendages<br />
are otherwise similar. Although the distribution is vast,<br />
there is no doubt that all the material identified here is<br />
the one species, and furthermore such a distribution is<br />
not unique (e.g. Metacirolana caeca (Hansen, 9 6), see<br />
Svavarsson & Bruce 2000), possible influenced by the<br />
Great Global Conveyer currents (e.g. see Manighetti<br />
200 ).<br />
A rather similar Aega sp. collected east <strong>of</strong> Heron<br />
Island, Queensland (NTM, unregistered, see p. 244) can<br />
be distinguished by the far smaller antennule peduncle<br />
articles 1 and 2, flat frontal lamina, more anteriorly<br />
rounded head, more and larger robust setae on pereopods<br />
–3, lack <strong>of</strong> large robust setae on the pleotelson<br />
and uropods, more rounded uropodal exopod, and by<br />
the different nature <strong>of</strong> the robust pitting on the pereon<br />
and pleotelson.<br />
prey: <strong>The</strong>re are no recent prey identifications. Gadus<br />
morrhua and Scymnum microcephalum (= Scymno microcephalo),<br />
Somniosus microcephalus (= Greenland<br />
shark, Dalatiidae) (Schioedte & Meinert 879b); Centrophorus<br />
squamosus (gulper shark, Centrophoridae)<br />
(Kussakin 979); Hyperoglyphe antarctica [bluenose and<br />
matiri (<strong>New</strong> <strong>Zealand</strong>) or Antarctic butterfish, Centro-<br />
lophidae].<br />
distribution: North Atlantic, South Africa and southwestern<br />
Pacific. Localities: Schioedte and Meinert<br />
( 879b) cite Iceland, Bergen, Floroe, Lodshagen and<br />
Farsun (all Norway), German Sea (= German Bight?)<br />
and Herne, Skagerak. South Africa (Barnard, 9 4);<br />
Bahamas (Treat 980); Hansen ( 9 6) cites the Færoe<br />
Islands and Jutland (Denmark) as regional records;<br />
Moray Firth and Shetland, Scotland (Norman 904).<br />
Present material is from the Chatham Rise, <strong>New</strong> <strong>Zealand</strong>,<br />
<strong>New</strong> Caledonia and from southeastern Australia<br />
<strong>of</strong>f the mid- and southern <strong>New</strong> South Wales coast.<br />
Previously recorded at depths <strong>of</strong> 460–730 m, present<br />
material at depths between 440 and 022 metres.<br />
44<br />
Aega semicarinata Miers, 875 (Figs 9–22)<br />
Æga semicarinata Miers, 1875: 115.– 1879: 201, pl. 11, figs.<br />
1–1d.– Dollfus, 1891: 57, pl. 8, figs 2–2a.– Bouvier, 1911:<br />
39, pl. 2, fig. 1.<br />
Aega semicarinata.– Barnard, 9 6: 06.– Nierstrasz, 93 :<br />
83.– Hale, 937: 9.– Barnard, 940: 40 .– Stephensen,<br />
1947: 23.– Menzies, 1962: 118, fig. 38A–C.– Moreira<br />
& Sadowsky, 1979: 109.– Kensley, 1978: 57, fig. 24I–J;<br />
980b: 59; 200 : 227.– Kussakin & Vasina, 982: 264.–<br />
Branch, Griffiths, Kensley & Sieg, 1991: 26.– Bruce, Lew<br />
Ton & Poore, 2002: 62.<br />
Æga semicarinatus.– Stebbing, 920: 334.<br />
Aega bicavata Nordenstam, 1930, 547, fig. 11, Pl. 20, fig. 11.<br />
Aega (Aega) bicavata.– Brusca, 983: 0.<br />
Aega (Aega) semicarinata.– Brusca, 983: .<br />
type LoCaLity: Kerguelen, southern Indian Ocean (Miers<br />
875).<br />
types: At the then British Museum <strong>of</strong> Natural History,<br />
London, according to Miers ( 875). Not listed by Ellis<br />
( 98 ), though one unregistered specimen, labelled as<br />
‘type’ is held at <strong>The</strong> Natural History Museum, London.<br />
<strong>The</strong> type information states: ‘outside label destroyed at<br />
Godstone’ and the only other data is ‘HMS Sylvia’.<br />
materiaL examined: Holotype(?), ♀ (non-ovig 27 mm),<br />
‘outside label destroyed at Godstone’, HMS Sylvia<br />
(BMNH).<br />
<strong>New</strong> <strong>Zealand</strong> material: ♀ (74 mm ovig, previously<br />
dissected), I685, vicinity <strong>of</strong> Bounty Plateau, 48° 9.50–<br />
7.20’S, 79°29.50–40’W, 6 March 979, dredged, 722<br />
m and ♂ (35 mm), stn Z3, labels in tube: “Z3, 40F, A,<br />
8/63”; “?John Graham, Oamaru, “♂ <strong>of</strong> giant I685”;<br />
therefore presumably same data as I685 (NIWA 2377<br />
♀, 23772 ♂). ♂ (38 mm), Chatham Rise, 42°43.95’S,<br />
79°53.9 ’W, 8 April 200 , 076–990 m, coll. S. O’Shea<br />
on RV Tangaroa (NIWA 23773). ♂ (3 mm), Chatham<br />
Rise, 43°49.605’S, 78°29.284’E, 6 October 200 , 454 m,<br />
Agassiz trawl, coll. RV Tangaroa (NIWA 23774). ♂ (3<br />
mm), North Otago, 27.5 m, 962, coll. John Graham<br />
(NMNZ Cr. 20 6). ♀ (~4–5 cm, ovig, broken, two<br />
pieces), 44°4 .35’S, 72°34.0’E, 390–360 m, RV James<br />
Cook (NMNZ Cr.4969).<br />
Additional material: ♀ (63 mm, non-ovig), Juan<br />
Fernandez Islands, Chile, 920, A 463 (LACM 20-<br />
2. ). 2♀ (44, 38 mm), <strong>of</strong>f Table Bay, South Africa<br />
(BMNH 93 . . 0. 8-20, part). ♂ (39 mm), Chatham<br />
Rise, 42.7 60–7 08°S, 80.0390–0297°E, 28 May 2006,<br />
935– 2 0 m, coll. RV Tangaroa (NIWA 25658).<br />
desCription: Body 2.6 times as long as greatest width,<br />
dorsal surfaces punctate, widest at pereonite 6, lateral<br />
margins subparallel. Rostral point projecting anteriorly,<br />
not ventrally folded. Eyes large, not medially<br />
united, separated by about 9% width <strong>of</strong> head; each<br />
eye made up <strong>of</strong> ~36 transverse rows <strong>of</strong> ommatidia,
Figure 19. Aega semicarinata Miers, 875. NIWA 23773. A, dorsal view; B, lateral view; C, head; D, frons; E, pleotelson posterior<br />
margin; F, penial openings; G, antenna; H, antennule; I, antenna peduncle, dorsal view.<br />
each row with ~ 8 ommatidia; eye colour dark brown.<br />
Pereonite 1 and coxae 2–3 each with posteroventral angle<br />
rounded. Coxae 5–7 with entire oblique carina (raised,<br />
forming distinct ridge); posterior margins convex,<br />
posterolateral angle blunt (more than 45°). Pleon with<br />
pleonite visible in dorsal view; pleonite 4 with posterolateral<br />
margins extending to but not beyond posterior<br />
margin <strong>of</strong> pleonite 5; pleonite 5 with posterolateral<br />
45<br />
angles overlapped by lateral margins <strong>of</strong> pleonite 4.<br />
Pleotelson 0.7 times as long as anterior width, dorsal<br />
surface with 2 sub-median depressions (and posterior<br />
median depression); lateral margins straight, crenulate,<br />
posterior margin emarginate, with 0 RS.<br />
Antennule peduncle article 2 anterodistal lobe not<br />
extending beyond mid-point <strong>of</strong> article 3; articles 3<br />
and 4 0.4 times as long as combined lengths <strong>of</strong> articles
Figure 20. Aega semicarinata Miers, 875. NIWA 23773. A, mandible; B, mandible palp article 3; C, maxillule; D, maxillule<br />
apex; E, maxilla; F, maxilla apex; G, maxilliped; H, maxilliped articles 2–5; I, maxilliped article 5 (Leica).<br />
1 and 2, article 3 2.8 times as long as wide; flagellum<br />
with 9 articles, extending to mid-point <strong>of</strong> eye. Antenna<br />
peduncle article 2 inferior surface without distinct<br />
longitudinal suture; article 4 .2 times as long as wide<br />
(dorsally with wide longitudinal depression), 0.8 times<br />
as long as combined lengths <strong>of</strong> articles –3, with deep<br />
longitudinal groove, inferior margin plumose setae,<br />
and 0 short simple setae; article 5 not markedly wider<br />
or flatter than article 4, 1.5 times as long as article 4, 2.7<br />
times as long as wide, inferior margin with 0 palmate<br />
setae, anterodistal angle with cluster <strong>of</strong> 5 short simple<br />
setae; flagellum with 20 articles, extending to posterior<br />
<strong>of</strong> pereonite .<br />
46<br />
Frontal lamina flat, longer than greatest width, lateral<br />
margins converging posteriorly, anterior margin<br />
rounded, without small median point, posterior margin<br />
abutting clypeus.<br />
Mandible molar process absent; palp article 2 with<br />
8 distolateral setae, palp article 3 with 20 setae (proximally<br />
smooth, distally finely serrate). Maxillule with<br />
8 terminal and subterminal RS (proximal 3 falcate).<br />
Maxilla mesial lobe with 3 RS ( hooked 2 straight);<br />
lateral lobe with 4 RS (large). Maxilliped endite with<br />
0 apical setae; palp article 2 with 3 RS (with further<br />
fine marginal setae); article 3 with 5 recurved RS (and<br />
slender); article 4 with 5 hooked RS; article 5 partly
Figure 21. Aega semicarinata Miers, 875. NIWA 23773. A–E, pereopods –3, 6 and 7, respectively.<br />
fused to article 4, distally convex, with 7 RS (straight,<br />
2 lateralmost curved).<br />
Pereopod 1 basis 2.2 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with RS (acute); merus<br />
inferior margin with 4 RS, set as two groups (<strong>of</strong> and<br />
3), superior distal angle with RS (small, acute); carpus<br />
0.7 as long as merus, inferior margin with 0 RS;<br />
propodus .8 times as long as proximal width, inferior<br />
margin with 0 RS, propodal palm with small distal lobe,<br />
dactylus smoothly curved, .2 as long as propodus.<br />
Pereopod 2 ischium inferior margin with 0 RS, superior<br />
distal margin with 2 RS (acute); merus inferior margin<br />
with 6 RS (set as 4 and 2), set as two groups, superior<br />
distal margin with 2 acute RS (short); carpus similar in<br />
size to that <strong>of</strong> pereopod , inferodistal angle with 0 RS,<br />
propodus without large club-shaped distal RS. Pereopod<br />
3 not similar to pereopod 2; propodus with large<br />
club-shaped distal RS. Pereopods 5–7 inferior margins<br />
<strong>of</strong> ischium–carpus with short RS. Pereopod 6 similar to<br />
47<br />
pereopod 7 (but larger and more robust, basis 2.8 times<br />
as long as wide). Pereopod 7 basis 3.4 times as long as<br />
greatest width, inferior margins with palmate setae;<br />
ischium 0.5 as long as basis, inferior margin with 3 RS<br />
(set singly), superior distal angle with 3 RS, inferior<br />
distal angle with 4 RS; merus 0.9 as long as ischium, 2<br />
times as long as wide, inferior margin with 6 RS (set<br />
as , 2 and 3), superior distal angle with 6 RS, inferior<br />
distal angle with 5 RS; carpus . as long as ischium,<br />
3.0 as long as wide, inferior margin with 5 RS (set as ,<br />
2 and 3), superior distal angle with 8 RS, inferior distal<br />
angle with 3 RS; propodus 0.9 as long as ischium, 3.8<br />
times as long as wide, inferior margin with 3 RS (set<br />
as and 2), superior distal angle with slender setae<br />
(palmate), inferior distal angle with 3 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by 0% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod .4 times as long as wide, distally<br />
broadly rounded, lateral margin weakly convex, mesial<br />
margin strongly convex, with PMS on distal half; endo-
Figure 22. Aega semicarinata Miers, 875. NIWA 23773. except H. A–D, pleopods –3, 5 respectively; E, uropod; F, uropod<br />
exopod, ventral view; G, uropod apices (exopod to right); H, uropod, NMNZ Cr.4969.<br />
48
pod 2.2 times as long as wide, distally subtruncate,<br />
lateral margin strongly concave, with PMS on distal<br />
one-third, mesial margin with PMS on distal half;<br />
peduncle .6 times as wide as long, mesial margin<br />
with 0 coupling hooks. Pleopod 2 appendix masculina<br />
with straight margins, 0.73 times as long as endopod,<br />
distally bluntly rounded. Exopods <strong>of</strong> pleopods –3<br />
each with distolateral margin not digitate; endopods<br />
<strong>of</strong> pleopods 3–5 each with distolateral point (minute);<br />
pleopods 2–4 peduncle distolateral margin with prominent<br />
acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about two-thirds as long as endopod.<br />
Uropod rami not extending beyond pleotelson, marginal<br />
setae dense, in several tiers, apices broadly rounded.<br />
Endopod apically not bifid, lateral margin straight, without<br />
prominent excision, proximal lateral margin with<br />
0 RS, distal lateral margin with 2 RS, mesial margin<br />
weakly convex, with 5 RS. Exopod extending beyond<br />
end <strong>of</strong> endopod (slightly), 2.7 times as long as greatest<br />
width, apically not bifid; lateral margin weakly convex,<br />
with 2 RS; mesial margin straight or convex (distally<br />
convex), with 7 RS.<br />
femaLe: Pereopod 3 lacks the distal robust seta on<br />
the propodus. Oostegites arising from the coxae <strong>of</strong><br />
pereonites –5. Eggs are large, 2.8–3.5 mm diameter.<br />
Present data indicate that females may grow to a far<br />
larger size than do the males.<br />
size: Up to 75 mm for the largest female examined here<br />
making it the largest aegid species. Males are smaller,<br />
present material measuring from 27 to 38 mm.<br />
Variation: <strong>The</strong> small number (five entire) <strong>of</strong> specimens,<br />
the fact that the uropod apices are mostly damaged<br />
with the robust setae rubbed <strong>of</strong>f and the presence <strong>of</strong><br />
a dense fringe <strong>of</strong> plumose setae means that it is not<br />
possible to precisely detail the variation present. <strong>The</strong><br />
robust setae on the merus <strong>of</strong> pereopods –3 present a<br />
constant pattern <strong>of</strong> two rows, although the number<br />
<strong>of</strong> robust setae varies: pereopod merus with +2 to<br />
+3, pereopods 2 and 3 merus with 4+2 to 5+2, one<br />
pereopod 3 with 3+2. Pereopod 3 has a large robust<br />
seta adjacent to the base <strong>of</strong> the dactylus on males<br />
NIWA23773, 23772 but this seta is absent or reduced<br />
in females and absent in the male NMNZ Cr.9269. It is<br />
not possible to say whether or not the absence <strong>of</strong> this<br />
robust seta is due to damage, although the presence or<br />
absence <strong>of</strong> such a prominent character would generally<br />
be considered to be significant.<br />
<strong>The</strong>re is some variation in the shape <strong>of</strong> the uropod<br />
endopod with some specimens having a distinctly oblique<br />
mesial margin (Fig. 22E) in others it is subtruncate<br />
(Fig. 22H). <strong>The</strong> robust setae vary: uropod exopod lateral<br />
margin 0– 2, mesial margin 5–8; endopod lateral<br />
margin 2–3, mesial margin 5–8.<br />
49<br />
<strong>The</strong> specimen from Juan Fernandez agrees well<br />
with the description presented here with the exception<br />
that the frontal lamina is shorter and wider, the<br />
mesial margin <strong>of</strong> the uropodal exopod is slightly more<br />
convex than illustrated and the RS on pereopod 2 form<br />
a single row rather than two groups. <strong>The</strong> pleotelson<br />
indentation is a little shallower but this may be due to<br />
damage as it is clearly eroded and rubbed. Counts for<br />
the RS on the merus <strong>of</strong> pereopods and 2: P : +3, +2;<br />
P2: 5+2 (both). <strong>The</strong> uropod endopod mesial margins<br />
both have 9 RS, slightly higher than for <strong>New</strong> <strong>Zealand</strong><br />
specimens.<br />
remarks: Aega semicarinata, one <strong>of</strong> the largest species<br />
<strong>of</strong> <strong>Aegidae</strong>, may be identified by the widely excavate<br />
and crenulated posterior margin <strong>of</strong> the pleotelson, the<br />
dorsal surface <strong>of</strong> which has two shallow submedian<br />
depressions and one median posterior depression.<br />
<strong>The</strong>se depressions can give the impression <strong>of</strong> a weakly<br />
defined longitudinal ridge, presumably after which<br />
Miers named the species. <strong>The</strong> moderately expanded<br />
antennule, large but separate eyes, coxae prominent in<br />
dorsal view, simple propodus on pereopods –3 and<br />
pereopod 3 propodus usually with a large club-like robust<br />
seta all serve to further distinguish the species.<br />
A number <strong>of</strong> names have been placed in synonymy<br />
with this species — Aega bicavata Nordenstam, 930,<br />
A. punctulata Miers, 88 and Aega urotoma Barnard,<br />
9 4. Material from <strong>New</strong> <strong>Zealand</strong> provisionally identified<br />
as Aega semicarinata proved to belong to two similar<br />
but distinct species, here identified as Aega semicarinata<br />
and Aega urotoma, the latter proving to be the same as<br />
specimens <strong>of</strong> that species from South Africa.<br />
<strong>The</strong> synonymy <strong>of</strong> Aega bicavata with A. semicarinata<br />
was first proposed by Menzies (1962), and the figures<br />
and description provided by Nordenstam agree well<br />
with Miers’ ( 879) description as well with the specimens<br />
examined here, and that species is retained as a<br />
junior synonym.<br />
Aega punctulata should never have been placed in<br />
synonymy as Miers’ (1881) description and figures<br />
more than adequately describe the critical points <strong>of</strong><br />
difference between the two species, including the<br />
smaller eyes and evenly rounded pleotelson posterior<br />
margin.<br />
<strong>The</strong> similar Aega urotoma, first placed into synonymy<br />
by Stebbing ( 920), has the antennule peduncle articles<br />
and 2 far more widely expanded, antenna peduncle<br />
article 5 flattened and expanded, smaller eyes, subtruncate<br />
or shallowly indented pleotelson which also lacks<br />
the prominent sub-lateral and posterior depressions<br />
seen in A. semicarinata (Table ).<br />
Another similar and very poorly characterised species<br />
is Aega webbii (Guérin-Méneville, 836) which is<br />
similar to A. semicarinata in eye size and in the emarginate<br />
shape <strong>of</strong> the posterior margin <strong>of</strong> the pleotelson. It
table 1. Comparison <strong>of</strong> Aega semicarinata with similar species <strong>of</strong> Aega.<br />
species references Eyes A2, articles 4 pereopod 2 pleotelson shape<br />
and 5 propodus<br />
A. semicarinata Present study Large, not Regular Without club-seta Emarginate, without<br />
medially united median point<br />
A. urotoma Present study Medium, Flat, expanded With club-seta Subtruncate, without<br />
widely separate median point<br />
A. chelipous Barnard 960 Large, not ? Without club-seta Subtruncate, with<br />
Barnard, 960 medially united median point<br />
A. concinna Hale 940 Small, Regular With club-seta Rounded<br />
Hale, 940 widely separate<br />
A. crenulata Kussakin 979 Eyes in contact Regular With club-seta Subtruncate<br />
Lütken, 859<br />
A. stroemii Kussakin 979 Eyes narrowly Regular ? Emarginate<br />
Lütken, 859 separated<br />
A. webbii Guérin- Schioedte & Medium, Flat? With club-seta Rounded, with<br />
Méneville, 836 Meinert 979b widely separate* median indentation<br />
is difficult to make detailed comparisons, but A. webbii<br />
differs in having antennule peduncle article 2 more<br />
strongly produced and a large robust seta opposing<br />
the dactylus <strong>of</strong> pereopod 2, character states lacking in<br />
both male and females <strong>of</strong> A. semicarinata.<br />
prey: <strong>The</strong> only record is that <strong>of</strong> Polyprion prognatus<br />
(Nordenstam 930).<br />
distribution: Straits <strong>of</strong> Magellan eastwards to <strong>New</strong><br />
<strong>Zealand</strong>, all records are south <strong>of</strong> about 35° latitude.<br />
Localities: Straits <strong>of</strong> Magellan, Kerguelen; Falkland<br />
Islands (Stebbing 1920 — record not confirmed); Chile<br />
(Menzies 962); Juan Fernandez Islands (Nordenstam<br />
930); South Africa (Kensley 978); Kerguelen, and<br />
<strong>of</strong>f the Crozet Islands in the southern Indian Ocean<br />
(Stephenson 1947 — record not confirmed), Marion<br />
Island (Kensley 980; Kussakin & Vasina 982); Macquarie<br />
Island (Hale 937).<br />
At depths between metres (‘amongst kelp’—Hale<br />
937) and 400 metres, material from <strong>New</strong> <strong>Zealand</strong> 27.5<br />
to 076 metres.<br />
Aega stevelowei sp. nov. (Figs 23–26)<br />
materiaL examined: Holotype: ♀ (non-ovig. 48 mm), <strong>of</strong>f<br />
Great Barrier Island, North Island [~ 36.3°S, 75.5°E],<br />
October 2004, from Hyperoglyphe antarctica, longline at<br />
~500 m, coll. Steve Lowe (NIWA 7973).<br />
Paratypes: ♀ (non-ovig. 40 mm), same data as holotype<br />
(NIWA 7974). ♂ (31 mm), 45 km southwest <strong>of</strong><br />
Beachport, Victoria, Australia, 37°45.00’S, 39°4 .00’E,<br />
24 October 98 , 390 m (NMV J277 4).<br />
desCription: Body 2.8 times as long as greatest width,<br />
dorsal surfaces punctate, widest at pereonite 5, lateral<br />
margins subparallel. Rostral point projecting anteri-<br />
50<br />
orly, not ventrally folded. Eyes large, medially united,<br />
anterior clear field 15% length <strong>of</strong> head, posterior clear<br />
field 43% length <strong>of</strong> head; each eye made up <strong>of</strong> ~24<br />
transverse rows <strong>of</strong> ommatidia, each row with ~8– 0<br />
ommatidia; eye colour black. Pereonite 1 and coxae 2–3<br />
each with posteroventral angle rounded, or right-angled<br />
(pereonite rounded, coxae 2 and 3 quadrate).<br />
Coxae 5–7 with entire oblique carina; posterior margins<br />
straight, posterolateral angle acute (less than 45°). Pleon<br />
with pleonite visible in dorsal view; pleonite 4 with<br />
posterolateral margins extending to but not beyond<br />
posterior margin <strong>of</strong> pleonite 5; pleonite 5 with posterolateral<br />
angles overlapped by lateral margins <strong>of</strong> pleonite<br />
4. Pleotelson 0.7 times as long as anterior width, dorsal<br />
surface without longitudinal carina; lateral margins<br />
weakly convex, crenulate (weakly), posterior margin<br />
at angle to lateral margins and converging to caudomedial<br />
point, with 3 RS.<br />
Antennule peduncle article 2 anterodistal lobe not<br />
extending beyond mid-point <strong>of</strong> article 3; articles 3 and<br />
4 0.6 times as long as combined lengths <strong>of</strong> articles and<br />
2, article 3 3.9 times as long as wide; flagellum with 15<br />
articles, extending to anterior <strong>of</strong> pereonite . Antenna<br />
peduncle article 2 inferior surface with distinct longitudinal<br />
suture; article 4 .8 times as long as wide, 0.8<br />
times as long as combined lengths <strong>of</strong> articles –3, with<br />
deep longitudinal groove, inferior margin 0 plumose<br />
setae, and short simple setae (minute, distal); article<br />
5 not markedly wider or flatter than article 4, 1.3 times<br />
as long as article 4, 3.0 as long as wide, inferior margin<br />
with 0 palmate setae, anterodistal angle with cluster<br />
<strong>of</strong> 3 short simple setae (plus 1 palmate seta); flagellum<br />
with 27 articles, extending to pereonite 4.<br />
Frontal lamina flat, longer than greatest width, rectangular,<br />
anterior margin rounded, forming median<br />
angle, posterior margin abutting clypeus.
Figure 23. Aega stevelowei sp. nov. Holotype, except H and I, paratype NIWA 7974. A, dorsal view; B, lateral view; C, head;<br />
D, frons; E, pleotelson; F, pleotelson, posterior margin; G, pleonites, alteral view; G, antenna; H, antennule; I, antenna peduncle,<br />
dorsal view.<br />
5
Figure 24. Aega stevelowei sp. nov. Paratype NIWA 7974. A, mandible; B, mandible palp article 3; C, maxillule apex;<br />
D, maxilla; E, maxilla apex; F, maxilliped; G, maxilliped articles 2–5; H, I, maxilliped articles 4 and 5.<br />
Mandible molar process absent; palp article 2 with<br />
3 distolateral setae (proximal 4 longest), palp article 3<br />
with 25 setae. Maxillule with 8 terminal and subterminal<br />
RS (proximal 2 hammer-head). Maxilla mesial lobe<br />
with 5 RS (2 lateral hooked, 3 mesial weakly curved);<br />
lateral lobe with 4 RS (proximal RS minute). Maxilliped<br />
endite with 2 apical setae (long CP); palp article 2 with<br />
0 RS (with 2 long straight simple setae); article 3 with<br />
3 recurved RS (small; and large curved RS); article 4<br />
with 7 hooked RS (5 large, 2 small; lateral margin with<br />
5 long straight RS); article 5 wholly (imperceptibly)<br />
fused to article 4, distally convex, with 6 RS.<br />
52<br />
Pereopod 1 basis .9 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 3 RS; merus inferior<br />
margin with RS (proximal), superior distal angle with<br />
2 RS; carpus 0.6 as long as merus, inferior margin with<br />
0 RS; propodus .4 times as long as proximal width,<br />
inferior margin with 0 RS, propodal palm with small<br />
distal lobe, dactylus smoothly curved, .0 as long as<br />
propodus. Pereopod 2 ischium inferior margin with<br />
RS, superior distal margin with 2 RS; merus inferior<br />
margin with 6 RS, set as two rows (<strong>of</strong> 4 + 2), superior<br />
distal margin with 2 acute RS (and 2 simple setae);
Figure 25. Aega stevelowei sp. nov. Holotype. A–E, pereopods , 2 and 7 respectively; D, distomesial margin, pereopod 7<br />
carpus.<br />
carpus similar in size to that <strong>of</strong> pereopod , inferodistal<br />
angle with RS, propodus without large club-shaped<br />
distal RS. Pereopod 3 similar to pereopod 2; propodus<br />
without large club-shaped distal RS. Pereopods<br />
5–7 inferior margins <strong>of</strong> ischium–carpus with short RS.<br />
Pereopod 6 similar to pereopod 7. Pereopod 7 basis 3.3<br />
times as long as greatest width, inferior margins with<br />
3 palmate setae; ischium 0.5 as long as basis, inferior<br />
margin with 4 RS (set as , and 2, plus submarginal),<br />
superior distal angle with 6 RS, inferior distal angle<br />
with 8 RS; merus .3 as long as ischium, 3.5 times as<br />
long as wide, inferior margin with 4 RS (set as and 3),<br />
superior distal angle with 4 RS, inferior distal angle<br />
with 5 RS; carpus .0 as long as ischium, 3.7 times as<br />
long as wide, inferior margin with 4 RS (set as and<br />
3), superior distal angle with 9 RS, inferior distal angle<br />
with RS; propodus 0.6 as long as ischium, 3.7 times<br />
as long as wide, inferior margin with 4 RS (set as ,<br />
53<br />
and 2), superior distal angle with 4 slender setae (<br />
distally plumose), inferior distal angle with 3 RS.<br />
Pleopod 1 exopod .6 times as long as wide, distally<br />
broadly rounded, lateral margin weakly concave, mesial<br />
margin strongly convex, with PMS on distal half;<br />
endopod 2. times as long as wide, distally subtruncate,<br />
lateral margin straight, with PMS on distal margin only,<br />
mesial margin with PMS on distal half; peduncle .5<br />
times as wide as long, mesial margin with 0 coupling<br />
hooks. Exopods <strong>of</strong> pleopods –3 each with distolateral<br />
margin not digitate; endopods <strong>of</strong> pleopods 3–5 each<br />
with distolateral point; pleopods 2–4 peduncle distolateral<br />
margin without prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 0 RS,<br />
posterior lobe about three-quarters as long as endopod.<br />
Uropod rami extending to pleotelson apex, marginal<br />
setae in two or three tiers, apices acute. Endopod apically<br />
deeply bifid, lateral process prominent (apex with
Figure 26. Aega stevelowei sp. nov. Female paratype NIWA 7974, except B and J, male paratype, NMV J277 4. A–D, pleopods<br />
–3, 5 respectively; E, distomesial margin, pleopod 3 exopod; F, uropod endopod apex; G, uropod exopod, apex; H, uropod<br />
endopod, ventral view; I, uropod; J, sternite 7 showing penial openings.<br />
54
4 teeth), lateral margin straight, without prominent<br />
excision, proximal lateral margin with 2 RS, distal<br />
lateral margin with 2 RS, mesial margin sinuate, with<br />
7 RS. Exopod not extending to end <strong>of</strong> endopod, 3.0 as<br />
long as greatest width, apically sub-bifid, mesial process<br />
prominent; lateral margin weakly convex, with 8<br />
RS; mesial margin sinuate, proximally concave, with<br />
4 RS.<br />
maLe: Similar to female. Maxilliped palp setation similar,<br />
but line <strong>of</strong> fusion on palp article 5 is visible, and<br />
the distal margin <strong>of</strong> palp article 5 has 6 RS as does the<br />
female. Penes low tubercles; penial openings separated<br />
by 8% <strong>of</strong> sternal width. Pleopod 2 appendix masculina<br />
with straight margins, 0.9 times as long as endopod,<br />
distally narrowly rounded.<br />
size: Holotype 48 mm; paratypes female 40 mm, mature<br />
male 3 mm.<br />
Variation: Pleotelson with 3– 7 RS as 6+7 or 8+7; the<br />
specimen with 7 RS had one margin with 0 RS, an<br />
asymmetric distribution <strong>of</strong> RS, which is probably aberrant.<br />
Uropod exopod mesial margin with 4 RS, lateral<br />
margin with 6–9 RS; uropod endopod mesial margin 7<br />
(6 once) lateral margin with 2+2 (2+ once). Pereopod<br />
merus inferior margin all with single proximal RS,<br />
distally with 2 or 3 simple setae; pereopods 2 and 3<br />
merus inferior margin with 4+2 RS. <strong>The</strong> uropodal and<br />
pleotelson RS are generally small, difficult to observe,<br />
and if missing a socket is hard to detect.<br />
<strong>The</strong> maxilliped palp has article 5 wholly fused in the<br />
female, but a faint trace <strong>of</strong> the line <strong>of</strong> fusion (or former<br />
articulation) is visible in the male.<br />
<strong>The</strong> dorsal setae are easily rubbed away, and are<br />
far less evident in the older net-caught male specimen<br />
from southern Australia.<br />
prey: Hyperoglyphe antarctica [bluenose, matiri (<strong>New</strong><br />
<strong>Zealand</strong>) or Antarctic butterfish, Centrolophidae].<br />
remarks: This distinctive species can be recognised by<br />
the finely setose dorsal body surfaces in conjunction<br />
with large, medially united eyes, ventrally flat and<br />
elongate frontal lamina, apically bifid uropod apices<br />
(when entire), uropod endopod with a distinctly sinuate<br />
mesial margin, and the weakly crenulated pleotelson<br />
posterior margins being angled towards a median<br />
point <strong>of</strong> inflexion (but without a produced point).<br />
Pereopod is unusual in that the merus inferior<br />
margin is largely devoid <strong>of</strong> robust setae, with only<br />
a single proximal robust seta in comparison to pereopods<br />
2 and 3 which have 6. Similarly reduced setation<br />
occurs in Aega falklandica but in that species the<br />
pereopod merus has a single distal robust seta on<br />
the inferior margin. <strong>The</strong> maxilliped is also unusual<br />
55<br />
within the genus in having numerous setae along the<br />
mesial margin <strong>of</strong> palp article 4, and two prominent<br />
circumplumose setae on the endite, in both the male<br />
and the female. <strong>The</strong> robust setae <strong>of</strong> the uropodal and<br />
pleotelson margins are relatively small, and those <strong>of</strong><br />
the pleotelson set in a marginal groove (as for species <strong>of</strong><br />
Aegapheles), and are <strong>of</strong>ten obscured by setae rendering<br />
them difficult to observe.<br />
Only one other species <strong>of</strong> Aega, A. punctulata (see<br />
Appendix 2) has setose body surfaces, but that species<br />
is otherwise abundantly distinct, with small well-<br />
separated eyes, short anteriorly rounded frontal<br />
lamina, pereopod merus with large robust setae,<br />
and rounded pleotelson posterior margin. <strong>The</strong> form <strong>of</strong><br />
the dorsal setae <strong>of</strong> A. punctulata differs from those <strong>of</strong><br />
A. stevelowei, the former with prominent stiff setae, the<br />
latter with fine flexible setae.<br />
distribution: Off Great Barrier Island, northeastern<br />
<strong>New</strong> <strong>Zealand</strong>, and Victoria, southern Australia; recorded<br />
depths <strong>of</strong> 390 and 500 metres.<br />
etymoLogy: Named for Mr Steve Lowe <strong>of</strong> Leigh, Auckland,<br />
who collected and donated significant material<br />
to this study.<br />
Aega urotoma Barnard, 9 4 (Figs 27–30)<br />
Aega urotoma Barnard, 9 4: 367, pl. 32A.– Kensley, 200 :<br />
227.<br />
Aega semicarinata.– Barnard, 9 6: 06 (not A. semicarinata<br />
Miers, 875).<br />
Aega webbi.– Trilles & Justine, 2004: 228, figs 9, 0<br />
(misidentification, not A. webbi Guérin-Menéville,<br />
836).<br />
materiaL examined: ♀ (non-ovig. 34 mm), <strong>of</strong>f southwestern<br />
South Island, 46°29.8’S, 66°02.3’E, 20 November<br />
986, stn AB /097/86 55 m, on ‘wing’ <strong>of</strong> Raja<br />
nasuta, trawl catch (NMNZ Cr. 20 7).<br />
Also examined: South Africa. ♀ (non-ovig. 44 mm),<br />
34°07’S, 25°54’E, 9 May 993, 0 m, coll. RV Africana<br />
(SafM A43 6). 2♀ (non-ovig. 36, 38 mm), south <strong>of</strong><br />
Still Bay, 35° 7’S, 2 °32’E, 27 May 993, 6 m, coll. RV<br />
Africana (SafM exA43 3). ♂ (2 mm), <strong>of</strong>f Table Bay,<br />
coll. S. African Mus. (BMNH 2003.23).<br />
Additional material: <strong>New</strong> caledonia. ♀ (non-ovig.<br />
30 mm), 22°55.7’S, 67° 7.0’E, 28 September 985,<br />
MUSORTOM IV, stn 2 5, 485–520 m, coll. B. Richer de<br />
Forges (MNHN Is.59 3). ♀ (non-ovig 20 mm), HALI-<br />
CAL 2, récolté sure un requin, Squalus melanurus, pêche<br />
a la palaugre, coll. Menon; (MNHN Is.59 4).<br />
desCription (<strong>of</strong> new zeaLand speCimen): Body 2.8 times<br />
as long as greatest width, dorsal surfaces punctate, widest<br />
at pereonite 5, lateral margins subparallel. Rostral<br />
point projecting anteriorly, not ventrally folded. Eyes<br />
small, combined widths less than 50% width <strong>of</strong> head,
Figure 27. Aega urotoma Barnard, 9 4. Female 32 mm (NMNZ Cr.9268). A, dorsal view, holotype; B, lateral view; C, head;<br />
D, frons; E, pleotelson posterior margin; F, antennule; G, antenna; H, antenna peduncle article –4, ventral view, ventral view;<br />
uropod.<br />
separated by about 29% width <strong>of</strong> head; each eye made<br />
up <strong>of</strong> ~ 8 transverse rows <strong>of</strong> ommatidia, each row with<br />
~ 0 ommatidia; eye colour dark brown. Pereonite 1 and<br />
coxae 2–3 each with posteroventral angle rounded, or<br />
with small distinct produced point (rounded with a<br />
small ventral point). Coxae 5–7 with entire oblique<br />
carina; posterior margins convex, posterolateral angle<br />
blunt (more than 45°). Pleon with pleonite visible in<br />
56<br />
dorsal view; pleonite 4 with posterolateral margins extending<br />
to but not beyond posterior margin <strong>of</strong> pleonite<br />
5; pleonite 5 with posterolateral angles overlapped by<br />
lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.8 times as long<br />
as anterior width, dorsal surface without longitudinal<br />
carina; lateral margins straight, crenulate, posterior<br />
margin subtruncate, with 0 RS.
Figure 28. Aega urotoma Barnard, 9 4. Female 32 mm (NMNZ Cr.9268). A, mandible; B, mandible palp, article 3; C, maxillule;<br />
D, maxillule apex; E, maxilla; F, maxilla apex; G, maxilliped; H, maxilliped palp; I, maxilliped palp article 5 (Leica).<br />
Antennule peduncle article 2 anterodistal lobe extending<br />
to end <strong>of</strong> article 3; articles 3 and 4 0.36 times as<br />
long as combined lengths <strong>of</strong> articles and 2, article 3 2.8<br />
times as long as wide (posterior margin with blade-like<br />
edge); flagellum with 8 articles, extending to mid-point<br />
57<br />
<strong>of</strong> eye. Antenna peduncle article 2 inferior surface with<br />
distinct longitudinal suture; article 4 .0 as long as<br />
wide, 0.8 times as long as combined lengths <strong>of</strong> articles<br />
–3, with deep longitudinal groove, inferior margin 0<br />
plumose setae, and 0 short simple setae; article 5 flat-
Figure 29. Aega urotoma Barnard, 9 4. Female 32 mm (NMNZ Cr.9268). A–D, pereopods , 2, 6 and 7 respectively; E, uropod<br />
endopod, apex; F, uropod exopod, apex.<br />
tened and expanded, .6 times as long as article 4, .5<br />
times as long as wide, inferior margin with 0 palmate<br />
setae, anterodistal angle with cluster <strong>of</strong> short simple<br />
seta; flagellum with 13 articles, extending to posterior<br />
<strong>of</strong> pereonite .<br />
Frontal lamina flat (lateral margins bent ventrally),<br />
as wide as long, lateral margins converging posteriorly,<br />
anterior margin rounded, with small median point,<br />
posterior margin not abutting clypeus.<br />
Mandible molar process absent; palp article 2 with<br />
58<br />
5 distolateral setae (4 large, small), palp article 3 with<br />
24 setae. Maxillule with 8 terminal and subterminal RS<br />
(proximal 4 hammer-head). Maxilla mesial lobe with<br />
4 RS (2 recurved, 2 straight); lateral lobe with 4 RS<br />
(large hooked). Maxilliped endite with apical seta;<br />
palp article 2 with 2 RS; article 3 with 6 recurved RS (<br />
being minute; with single simple seta); article 4 with<br />
6 hooked RS; article 5 partly fused to article 4, distally<br />
convex, with 6 RS (mesial 2 being hooked, remainder<br />
straight).
Figure 30. Aega urotoma Barnard, 9 4. Female 32 mm (NMNZ Cr.9268). A–E, pleopods –5 respectively; F, uropod; G,<br />
uropod exopod, ventral view.<br />
Pereopod 1 basis .6 times as long as greatest width<br />
(basis with prominent lateral carina); ischium 0.4 times<br />
as long as basis, inferior margin with 0 RS, superior<br />
distal margin with RS (small acute); merus inferior<br />
margin with 3 RS (with inferior lobe), set as two groups<br />
(<strong>of</strong> and 2), superior distal angle with RS (minute);<br />
carpus .0 as long as merus, inferior margin with 0<br />
RS; propodus . times as long as proximal width,<br />
inferior margin with 0 RS, propodal palm with small<br />
59<br />
distal lobe, dactylus abruptly hooked, .3 as long as<br />
propodus. Pereopod 2 ischium inferior margin with<br />
RS (round), superior distal margin with RS; merus<br />
inferior margin with 5 RS (set as 3 + 2), set as two<br />
groups, superior distal margin with 2 acute RS; carpus<br />
similar in size to that <strong>of</strong> pereopod , inferodistal<br />
angle with RS, propodus with large club-shaped<br />
distal RS. Pereopod 3 similar to pereopod 2; ischium<br />
inferior margin with 2 RS, propodus with large club-
shaped distal RS. Pereopods 5–7 inferior margins <strong>of</strong><br />
ischium–carpus with short RS. Pereopod 6 similar to<br />
pereopod 7 (more robust, with fewer RS on inferior<br />
margins). Pereopod 7 basis 2.4 times as long as greatest<br />
width (inferolateral margin strongly carinate), inferior<br />
margins with 5 palmate setae; ischium 0.5 as long as<br />
basis, inferior margin with 5 RS (set as , 3 and ), superior<br />
distal angle with 5 RS, inferior distal angle with<br />
4 RS; merus 0.8 as long as ischium, 2.2 times as long as<br />
wide, inferior margin with 4 RS (set as , 3), superior<br />
distal angle with 6 RS, inferior distal angle with 4 RS;<br />
carpus 0.7 as long as ischium, 2.2 times as long as wide,<br />
inferior margin with 3 RS (set singly), superior distal<br />
angle with 4 RS (short), inferior distal angle with 7 RS<br />
(short); propodus 0.6 as long as ischium, 2.9 times as<br />
long as wide, inferior margin with 4 RS (set as , and<br />
2), superior distal angle with 3 slender setae, inferior<br />
distal angle with 3 RS.<br />
Pleopod 1 exopod . times as long as wide, distally<br />
broadly rounded, lateral margin straight, mesial margin<br />
strongly convex, with PMS on distal half; endopod<br />
.8 times as long as wide, distally subtruncate, lateral<br />
margin strongly concave, with PMS on distal one-third,<br />
mesial margin with PMS on distal half; peduncle .8<br />
times as wide as long, mesial margin with 9 coupling<br />
hooks. Exopods <strong>of</strong> pleopods –3 each with distolateral<br />
margin not digitate; endopods <strong>of</strong> pleopods 3–5 each<br />
with distolateral point; pleopods 2–4 peduncle distolateral<br />
margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about three-quarters as long as endopod.<br />
Uropod rami not extending beyond pleotelson,<br />
marginal setae dense, in several tiers, apices broadly<br />
rounded. Endopod apically not bifid, lateral margin<br />
straight, without prominent excision, proximal lateral<br />
margin with RS, distal lateral margin with 2<br />
RS, mesial margin weakly convex, with 5 RS. Exopod<br />
not extending to end <strong>of</strong> endopod, 2.5 times as long as<br />
greatest width, apically not bifid (both rami with apical<br />
concavity); lateral margin convex, with 7 RS; mesial<br />
margin weakly convex, with 4 RS.<br />
Variation: <strong>The</strong> small number <strong>of</strong> specimens, only one<br />
from <strong>New</strong> <strong>Zealand</strong> waters, precludes precisely detailing<br />
the variation. <strong>The</strong> smaller specimens have the<br />
robust setae on the inferodistal angle <strong>of</strong> the ischium<br />
more acute than on large specimens.<br />
maLe.: Similar to the female. <strong>The</strong> single male specimen<br />
(South Africa, BMNH) was too brittle to dissect,<br />
but the penial openings are close-set but separate,<br />
and the appendix masculina is similar to that <strong>of</strong> Aega<br />
semicarinata.<br />
size: Present material 34 to 38 mm; Barnard’s specimen<br />
53 mm.<br />
60<br />
remarks: Aega urotoma bears a strong but superficial<br />
resemblance to Aega semicarinata, and has indeed<br />
been placed in synonymy with that species following<br />
the suggestion by Barnard ( 9 6) in a ‘Corrigenda’<br />
that the two species were the same [followed by Stebbing<br />
( 920) and later authors]. <strong>The</strong> two species, with<br />
overlapping distributions, are similar in general body<br />
shape and appearance, in the pattern <strong>of</strong> robust setae<br />
on the anterior pereopods and have similarly shaped<br />
uropods. Close examination shows numerous points<br />
<strong>of</strong> difference between the two species, and there is no<br />
doubt that Aega urotoma should be regarded as valid.<br />
Notably, in A. urotoma, the pleotelson posterior margin<br />
is not emarginate and the dorsal surface lacks the<br />
submedian depressions; the antennule and antenna <strong>of</strong><br />
A. urotoma are markedly dorso-ventrally compressed<br />
and expanded, particularly antenna peduncle article<br />
5 and this last character can be used to easily separate<br />
the two species. Further points <strong>of</strong> distinction are A.<br />
urotoma having smaller eyes, a prominent blade-like<br />
carina on the basis <strong>of</strong> all pereopods, pereopods 2 and<br />
3 are similar to each other, both bearing a club-like<br />
robust seta on the propodus, and the robust setae <strong>of</strong><br />
pereopods 6 and 7 are noticeably shorter and stouter<br />
than in Aega semicarinata.<br />
Trilles and Justine (2004) recorded, and described<br />
in part, specimens that they misidentified as Aega<br />
webbii (Guérin-Méneville, 836). During a visit to the<br />
Muséum national d’Histoire naturelle in Paris, the<br />
specimens examined by Trilles and Justine (2004) could<br />
not be located, but I examined further material from<br />
<strong>New</strong> Caledonia which agrees entirely with material<br />
here being identified as A. urotoma. Aega webbii has<br />
never been described in detail, but the figures given<br />
by Guérin-Méneville ( 836) show clearly that the eyes<br />
are far larger that <strong>of</strong> the present material, and that<br />
the posterior margin <strong>of</strong> the pleotelson is emarginate,<br />
character states that A. webbii shares with A. semicarinata.<br />
Digital images <strong>of</strong> the holotype (ANSP CA2779;<br />
kindly provided by Paul Callomon) also support these<br />
differences.<br />
prey: <strong>The</strong> only records to date are from Squalus melanurus<br />
the black-eared spurdog (Squalidae) in <strong>New</strong><br />
Caledonia and a trawl-caught Raja nasuta (Rajidae),<br />
the <strong>New</strong> <strong>Zealand</strong> rough skate (Rajidae).<br />
distribution: Cape Point (Barnard 9 4) and Cape<br />
Province (present material), South Africa; distribution<br />
is here extended to the southwestern Pacific, <strong>of</strong>f South<br />
Island, <strong>New</strong> <strong>Zealand</strong> and <strong>of</strong>f <strong>New</strong> Caledonia. <strong>The</strong><br />
species has long been placed in synonymy with Aega<br />
semicarinata and it is possible that some records <strong>of</strong> that<br />
species may be <strong>of</strong> A. urotoma. It is likely that A. urotoma,<br />
recorded here for the first time beyond South African<br />
waters, will have a Southern Ocean distribution. At<br />
depths <strong>of</strong> 0 to 329 metres.
Aega whanui sp. nov. (Figs 3 –33)<br />
materiaL examined: Holotype. ♀ (ovig. 59 mm), Lord<br />
Howe Rise, 27°50.03’S, 62°48.06’E, 5 May 989, 250<br />
m, coll. FRV Franklin (AM P43982).<br />
Additional material. ♀ (non-ovig. 49 mm,), Iles<br />
Tanimbar, Indonesia, Timor Sea, 08°39’S, 3 °08’E, 5<br />
November 99 , 084– 058 m, KARUBAR stn CP89,<br />
coll. RV Baruna Jaya (MNHN Is.5862).<br />
desCription: Body .6 times as long as greatest width,<br />
dorsal surfaces coarsely punctate, widest at pereonite<br />
5, lateral margins weakly ovate. Rostral point projecting<br />
anteriorly, not ventrally folded. Eyes moderate,<br />
combined widths 50–65% width <strong>of</strong> head, separated<br />
by about 38% width <strong>of</strong> head; eye colour red (ommatidia<br />
not distinct). Pereonite 1 and coxae 2–3 each<br />
with posteroventral angle right-angled. Coxae 5–7<br />
with entire oblique carina; posterior margins straight,<br />
posterolateral angle blunt (more than 45°). Pleon with<br />
pleonite largely concealed by pereonite 7; pleonite 4<br />
with posterolateral margins extending clearly beyond<br />
posterior margin <strong>of</strong> pleonite 5; pleonite 5 with posterolateral<br />
angles overlapped by lateral margins <strong>of</strong> pleonite<br />
4. Pleotelson 0.6 times as long as anterior width, dorsal<br />
surface with 2 sub-medial depressions; lateral margins<br />
convex, serrate, posterior margin with distinct short<br />
median point, with 2 RS.<br />
Antennule peduncle article 2 anterodistal lobe not<br />
extending beyond mid-point <strong>of</strong> article 3; articles 3 and<br />
4 0.7 times as long as combined lengths <strong>of</strong> articles and<br />
2, article 3 2.7 times as long as wide; flagellum with 23<br />
articles, extending to anterior <strong>of</strong> pereonite . Antenna<br />
peduncle article 2 inferior surface without distinct longitudinal<br />
suture; article 4 .6 times as long as wide, 0.9<br />
times as long as combined lengths <strong>of</strong> articles –3, with<br />
deep longitudinal groove, inferior margin 0 plumose<br />
setae, and 0 short simple setae; article 5 not markedly<br />
wider or flatter than article 4, 1.0 as long as article 4, 2.3<br />
times as long as wide, inferior margin with 4 palmate<br />
setae, anterodistal angle with cluster <strong>of</strong> 3 short simple<br />
setae; flagellum with 24 articles, extending to posterior<br />
<strong>of</strong> pereonite 2.<br />
Frontal lamina flat, longer than greatest width, lateral<br />
margins parallel, anterior margin rounded, without<br />
small median point, posterior margin not abutting<br />
clypeus.<br />
Mandible molar process absent; palp article 2 with<br />
4 distolateral setae (plus row <strong>of</strong> 4 submarginal and<br />
scattered small simple setae; all finely biserrate), palp<br />
article 3 with 35 setae (all finely biserrate; distal 2 markedly<br />
longer than remainder). Maxillule with 8 terminal<br />
and subterminal RS (proximal 3 falcate). Maxilla lateral<br />
lobe with 3 RS.<br />
Pereopod 1 basis .9 times as long as greatest width;<br />
ischium 0.3 times as long as basis, inferior margin with<br />
6<br />
RS (minute), superior distal margin with 0 RS ( slender<br />
seta); merus inferior margin with 0 RS, superior<br />
distal angle with 0 RS (4 slender setae); carpus 0.6 as<br />
long as merus, inferior margin with 0 RS; propodus<br />
.5 times as long as proximal width, inferior margin<br />
with 0 RS, propodal palm simple, without blade or<br />
process (concave), dactylus smoothly curved, . as<br />
long as propodus. Pereopod 2 ischium inferior margin<br />
with RS, superior distal margin with RS; merus<br />
inferior margin with 8 RS, set as single row, superior<br />
distal margin with acute RS; carpus similar in size<br />
to that <strong>of</strong> pereopod , inferodistal angle with 0 RS,<br />
propodus without large club-shaped distal RS. Pereopod<br />
3 similar to pereopod 2; propodus without large<br />
club-shaped distal RS. Pereopods 5–7 inferior margins<br />
<strong>of</strong> ischium–carpus with short RS. Pereopod 6 similar to<br />
pereopod 7. Pereopod 7 basis 3.2 times as long as greatest<br />
width, inferior margins with 4–8 palmate setae; ischium<br />
0.5 as long as basis, inferior margin with 9 RS (set as ,<br />
2, , and 4), superior distal angle with 5 RS, inferior<br />
distal angle with 7 RS; merus 0.8 as long as ischium,<br />
.9 times as long as wide, inferior margin with 8 RS (set<br />
as , 3 and 4), superior distal angle with 3 RS, inferior<br />
distal angle with 9 RS; carpus 0.8 as long as ischium, 2.3<br />
times as long as wide, inferior margin with 6 RS (set as<br />
3 and 3), superior distal angle with 9 RS, inferior distal<br />
angle with 0 RS; propodus 0.6 as long as ischium, 3.6<br />
times as long as wide, inferior margin with 5 RS (set as<br />
, 2 and 2), superior distal angle with 3 slender setae,<br />
inferior distal angle with 3 RS.<br />
Pleopod 1 exopod .8 times as long as wide, distally<br />
narrowly rounded, mesial margin weakly oblique,<br />
lateral margin straight, mesial margin strongly convex,<br />
with PMS on distal two-thirds; endopod 2.0 times<br />
as long as wide, distally subtruncate, lateral margin<br />
weakly concave, with PMS on distal margin only, mesial<br />
margin with PMS on entire margin; peduncle .8<br />
times as wide as long, mesial margin with 3 coupling<br />
hooks. Exopods <strong>of</strong> pleopods –3 each with distolateral<br />
margin not digitate; endopods <strong>of</strong> pleopods 3–5 each<br />
with distolateral point; pleopods 2–4 peduncle distolateral<br />
margin without prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about as long as endopod. Uropod rami<br />
extending to pleotelson apex, marginal setae in single<br />
tier, apices narrowly rounded. Endopod apically not<br />
bifid, lateral margin proximally convex and distally<br />
concave, without prominent excision, proximal lateral<br />
margin with 3 RS, distal lateral margin with 3 RS,<br />
mesial margin weakly convex, with 6–7 RS. Exopod<br />
extending to end <strong>of</strong> endopod, 3.2 times as long as<br />
greatest width, apically not bifid; lateral margin weakly<br />
convex, with 2 RS; mesial margin sinuate, proximally<br />
concave, with 5 RS (or 6).<br />
maLe: Not known.
Figure 31. Aega whanui sp. nov. Holotype. A, dorsal view; B, lateral view; C, head; D, frons; E, pleotelson and uropods; F,<br />
pleotelson, distal margin; G, antenna peduncle; H antennule.<br />
62
Figure 32. Aega whanui sp. nov. Holotype. A, mandible; B, mandible palp article 3; C, maxillule apex; D, maxillule; E, maxilla;<br />
F, maxilla apex; G, maxilliped articles 2–5.<br />
size: Females at 49 to 59 mm.<br />
Variation: <strong>The</strong> specimen from the Timor sea is less<br />
wide ( .9 times as long as wide) than the holotype, has<br />
black eyes, and the frontal lamina has a small median<br />
point; each eye with ~ 8 transverse rows <strong>of</strong> ommatidia,<br />
each row with ~ 2– 4 ommatidia.<br />
remarks: Aega whanui sp. nov. is a large and notably<br />
wide-bodied species, easily identified by the small<br />
eyes, ovate body shape, long antennule flagellum,<br />
short dactylus on pereopods –3 (about as long as<br />
63<br />
propodus), proportionally long basis on pereopods<br />
–3 and setation <strong>of</strong> the pereopods and uropods. <strong>The</strong>re<br />
are no closely similar species. Aega whanui has an<br />
unusual mandible morphology, with a near truncate<br />
distal portion which has the incisor reduced to a small<br />
triangular point. Whether this is also the case for the<br />
male is unknown, but the mandible incisor has not<br />
been shown to be sexually variable for any other aegid<br />
species. <strong>The</strong> weakly developed eyes <strong>of</strong> the holotype is<br />
presumably a preservation artefact.<br />
prey: Not known.
Figure 33. Aega whanui sp. nov. Holotype. A–C, pereopods , 2 and 7 respectively; D, pereopod propodus, later view; D,<br />
pereopod propodus, mesiodistal angle; F, pleopod ; G, uropod exopod, ventral view; H, uropod.<br />
64
distribution: Lord Howe Rise to the northwest <strong>of</strong> <strong>New</strong><br />
<strong>Zealand</strong>, and Timor Sea, <strong>of</strong>f the Tanimbar Islands,<br />
Indonesia; at depths from 084 to 250 m.<br />
etymoLogy: Whanui (pronunciation: ‘phanui’) is a<br />
Mäori word that means wide or broad (alluding to<br />
body shape).<br />
Aega sp.<br />
materiaL examined: 2 mancas (4.5, 5.0 mm), Wanganella<br />
Bank, Norfolk Ridge, 32°34.4’S, 67°3 .0’E, 29 January<br />
98 , 3 m, NIWA stn 0.63 (NMNZ Cr.487 ).<br />
remarks: <strong>The</strong> eyes are widely separate, the antenna is<br />
notably short, being only a little longer than the antennule,<br />
the body shape is elongate (similar to that <strong>of</strong> Aega<br />
alazon), the uropods are slender, with the distal margin<br />
<strong>of</strong> the exopod serrate. <strong>The</strong>se two small mancas cannot<br />
be identified as any named species. <strong>The</strong> specimens are<br />
<strong>of</strong> uniform appearance, but given that many <strong>Aegidae</strong><br />
will grow from four times to 0 times larger, it is not<br />
possible to be confident that apparent species-specific<br />
characters will not change with maturity.<br />
Aegapheles gen. nov.<br />
type speCies: Aega kixalles Bruce, 2004; here designated.<br />
diagnosis: Body moderately to strongly dorsally<br />
vaulted. Rostral point acute, projecting anteriorly. Eyes<br />
present, large, usually medially united. Pleonite not<br />
abruptly narrower than pereonite 7; pleonite 4 with<br />
lateral margins extending beyond posterior margin <strong>of</strong><br />
pleonite 5. Pleotelson produced to an acute, <strong>of</strong>ten elongate<br />
point. Antennule peduncle articles and 2 weakly<br />
flattened, not expanded; anterodistal article 2 weakly or<br />
not produced. Maxillule with –3 large broad-based RS,<br />
several small RS. Maxilliped palp article 5 wider than<br />
longer; endite present. Pereopods –3 merus inferior<br />
margin with large RS, usually set in one or more rows,<br />
pereopods 4–7 with long acute RS. Uropodal endopod<br />
lateral margin with weak to prominent excision; plane<br />
<strong>of</strong> endopod oblique, at angle <strong>of</strong> about 35° to that <strong>of</strong><br />
pleotelson and exopod.<br />
desCription: Body moderately to strongly vaulted,<br />
about 2 to 4 times as long as wide. Head with eyes, <strong>of</strong>ten<br />
large, may meet at midpoint; anterior margin with<br />
median rostral point. Coxae <strong>of</strong> pereonites 4–7 longer<br />
than respective segment, posteriorly produced. Pleon<br />
not abruptly narrower than pereon; pleonite 5 laterally<br />
65<br />
overlapped by pleonite 4; pleonites 3–5 posteriorly<br />
produced to an acute point.<br />
Frontal lamina with posterior margin not abutting<br />
clypeus. Mandible with uni- or bicuspid incisor;<br />
molar process present, reduced or absent. Maxilliped<br />
palp 5-articled, article shorter than wide, articles 3<br />
and 4 each with 2–6 stout recurved RS, article 5 sub-<br />
rectangular, with long flexible terminal setae; endite<br />
present, usually with –2 terminal setae. Uropodal<br />
rami with marginal setae in single tier.<br />
etymoLogy: From the Greek apheles (smooth, even—all<br />
species are smooth-bodied) in conjunction with Aega—<br />
indicating family affinity; alludes to the blood-feeding<br />
micropredator genus <strong>of</strong> mosquito. Gender feminine.<br />
remarks: <strong>The</strong> group <strong>of</strong> species, referred to as the ‘Aega<br />
deshaysiana-group’ by Bruce (2004a) forms a well-<br />
supported clade and is here established as the new<br />
genus Aegapheles. <strong>The</strong> unique apomorphy is the pleotelson<br />
posterior margin forming an extended point and<br />
the uropodal rami not reaching the posterior margin<br />
<strong>of</strong> the pleotelson. <strong>The</strong> genus is further characterised<br />
by all species having very large eyes either meeting at<br />
the midline or separated by the width <strong>of</strong> only one or<br />
two ommatidia, the posterior pereopods with elongate<br />
robust setae, the uropodal exopod lateral margin with<br />
a usually very distinct excision (very weakly present in<br />
some species <strong>of</strong> Aega), the robust setae on the inferior<br />
margins <strong>of</strong> the merus <strong>of</strong> pereopods and 2 forming one<br />
or more continuous rows and the plane <strong>of</strong> the uropodal<br />
exopod is at an oblique angle to the endopod (this state<br />
also occurring in Rocinela). <strong>The</strong> frontal lamina <strong>of</strong> species<br />
<strong>of</strong> Aegapheles is usually flat, <strong>of</strong>ten not distinctly defined<br />
posteriorly and does not abut against the clypeus; in a<br />
few species the posterior border is clearly defined.<br />
Seventeen named species are included in the genus,<br />
those below and under ‘Species included ...’ (p.2 3).<br />
Key to the new <strong>Zealand</strong> species <strong>of</strong> AegApheles<br />
Although not directly used to key the species <strong>of</strong> Aegapheles,<br />
the number <strong>of</strong> marginal robust setae on the<br />
uropodal and pleotelson margins are useful to confirm<br />
identity. <strong>The</strong> number <strong>of</strong> pleotelson robust setae<br />
is given in parentheses for each species at the end <strong>of</strong><br />
the couplet.<br />
Uropodal exopod extending posterior to endopod<br />
(i.e. longer than endopod) ....................................3<br />
– Uropodal exopod not extending posterior to endopod<br />
(i.e. as long as or shorter than endopod) ..<br />
..................................................................................2
2 Pereopod with propodal blade about as<br />
long as palm; frontal lamina anteriorly rounded,<br />
posteriorly narrow; pleotelson apex with<br />
distinct apical point (pleotelson with 8 RS) ..........<br />
......................................... Aegapheles copidis (p. 70)<br />
– Pereopod without distal propodal blade<br />
or lobe; frontal lamina quadrate; pleotelson<br />
apex produced, without distinct apical point;<br />
(pleotelson with 8– 0 RS) ........................................<br />
........................................ Aegapheles umpara (p. 8 )<br />
3. Eyes entirely united medially ..............................4<br />
– Eyes narrowly separate (pleotelson with 6–8<br />
RS) .....................................Aegapheles birubi (p. 68)<br />
4. Pereopods 5–7 superior margins <strong>of</strong> ischium to<br />
carpus without long setae .....................................5<br />
– Pereopods 5–7 superior margins <strong>of</strong> ischium to<br />
carpus with long setae (pleotelson with – 6<br />
RS) ..................................Aegapheles hamiota (p. 73)<br />
5. Frontal lamina ovate; pereopod propodal palm<br />
with prominent distal lobe; inferior margin <strong>of</strong><br />
pereopods 2 and 3 merus with near continuous<br />
row <strong>of</strong> 2– 5 RS (pleotelson with 0 RS) ................<br />
...................................Aegapheles rickbruscai (p. 79)<br />
– Frontal lamina anteriorly rounded; pereopod<br />
propodal palm with or without small distal<br />
lobe; inferior margin <strong>of</strong> pereopods 2 and 3 merus<br />
with 4–6 widely spaced RS ..................................6<br />
6. Frontal lamina lateral margins posteriorly narrowed;<br />
uropodal endopod with 2–4 RS proximal<br />
to lateral notch (pleotelson with 8– 0 RS) ............<br />
.......................................... Aegapheles alazon (p. 66)<br />
– Frontal lamina lateral margins sub-parallel; uropodal<br />
endopod with 6 or 7 RS proximal to lateral<br />
notch (pleotelson with 4– 8 RS) ...........................<br />
........................................Aegapheles mahana (p. 75)<br />
Aegapheles alazon (Bruce, 2004) comb. nov. (Fig. 34)<br />
Aega alazon Bruce, 2004: 156, figs 12–15, 62.– Poore, 2005: 6.<br />
materiaL examined: <strong>New</strong> <strong>Zealand</strong>: ♀ (non-ovig 3 mm),<br />
vicinity <strong>of</strong> the Snares, 47°20.0’S, 67°02,00’E, 0 October<br />
962, 74 m, stn B057 (NIWA 7930). ♀ (non-ovig 28<br />
mm), vicinity <strong>of</strong> the Snares, 48°46.00’S, 67°04.99’E, 3<br />
October 962, 43 m, stn B059 (NIWA 793 ). ♂ (25<br />
mm), vicinity <strong>of</strong> the Snares, 48°43.00’S, 67°3 .99’E,<br />
3 October 962, 6 m, stn B0593 (NIWA 7932).<br />
♀ (40 mm), west <strong>of</strong> Snares, 48°03.39’S, 66°45. 2’E,<br />
27 Feb 1993, 141–144 m, on fin <strong>of</strong> gemfish, coll. Tangaroa<br />
(NMNZ Cr. 2002). ♀ (non-ovig 2 mm), stn<br />
ABI/003/86, 46°00.0’S 70°42. ’E, 77 m, 5 November<br />
986, on gills <strong>of</strong> school shark, coll. B. Jones (NMNZ<br />
Cr. 20 8). ♀ (non-ovig. 58 mm), NW <strong>of</strong> McCauley Island,<br />
Kermadec Islands, 30°0 .5’S, 78°42.8’W, 30 Sept<br />
66<br />
993, 0 m, dropline (prey not recorded), coll. R Win<br />
on FV Te Maru 18 (AK 842 8). ♀ (non-ovig 28 mm),<br />
Z6 5, ex ling (dried at some point) (NIWA 7936). 2<br />
♀ (non-ovig 23, 33 mm), 7°25.00’S, 78° 0.00’E, 46 m<br />
(as 25 fms), 4/63, on horse mackerel, stn Z2 LH (NIWA<br />
7963). ♀ (non-ovig 35 mm), 7°25.00’S, 78° 0.00’E,<br />
79 m (as 43 fms), <strong>of</strong>f ‘groper’, Z3 /64 (NIWA 7964).<br />
♂ (32 mm), 7°25.00’S, 78° 0.00’E, 42 m (as 23 fms)<br />
Z2 2/63, coll. J. Graham (NIWA 7965). ♀ (non-ovig<br />
38 mm), Z2/63, J. Graham (NIWA 7966). ♂ (3 mm),<br />
7°25.00’S, 78° 0.00’E [vicinity <strong>of</strong> Fiji], on gurnard, Z2,<br />
5/62, 436, coll. Graham (NIWA 7967). Note: <strong>The</strong>re is<br />
some considerable doubt over the data for the specimens<br />
apparently taken at 7°S, the vicinity <strong>of</strong> Fiji (see<br />
comment Rocinela garricki, p. 69), as the host names<br />
are <strong>of</strong> <strong>New</strong> <strong>Zealand</strong> fishes.<br />
Additional material: south Atlantic: ♂ ( 4.2 mm),<br />
Discovery Expedition, Stn 87, from 2.2–0.8 miles S<br />
65ºE <strong>of</strong> South Hill, Inaccessible Is, Tristan Group, 8<br />
November 933, 35– 34 m (BMNH unreg). southwestern<br />
Pacific: Tonga: , (26 mm), 2° ’S, 75°27’W,<br />
6 June 2000, BORDAU 2, stn. CH 609, 385–405 m,<br />
coll. Bouchet et al. (MNHN Is.5879). <strong>New</strong> Caledonia: ,<br />
8°55.48’S, 63°22. ’E, 7 August 992, BATHUS 4, stn.<br />
CP927, 452–444 m, coll. B. Richer de Forges (MNHN<br />
Is.5865). (27 mm) <strong>New</strong> <strong>Zealand</strong>, <strong>of</strong>f Great Barrier<br />
Island, North Island, January 2006, old longline gear<br />
at ~500 m, coll. Steve Lowe (NIWA 23778).<br />
type LoCaLity: Off Port Elizabeth, South Africa (Bruce<br />
2004a).<br />
diagnosis (from Bruce 2004a): Eyes large, medially<br />
united, anterior clear field 21% length <strong>of</strong> head, posterior<br />
clear field 46% length <strong>of</strong> head; eye colour dark brown.<br />
Pleonite 4 with posterolateral margins extending clearly<br />
beyond posterior margin <strong>of</strong> pleonite 5; pleonite 5 with<br />
posterolateral angles overlapped by lateral margins <strong>of</strong><br />
pleonite 4. Pleotelson .0– .2 times as long as anterior<br />
width, dorsal surface without longitudinal carina;<br />
lateral margins convex, smooth, posterior margin with<br />
6– 0 RS.<br />
Antennule peduncle article 2 anterodistal lobe not<br />
extending beyond mid-point <strong>of</strong> article 3; flagellum extending<br />
to posterior margin <strong>of</strong> eye. Antenna peduncle<br />
article 2 inferior surface with indistinct groove; flagellum<br />
extending to posterior <strong>of</strong> pereonite .<br />
Frontal lamina flat, wider than long, lateral margins<br />
converging posteriorly, anterior margin rounded, with<br />
small median point, posterior margin not abutting<br />
clypeus.<br />
Pereopod 1 merus inferior margin with 3 RS, set as<br />
two groups (<strong>of</strong> and 2), superior distal angle with 2 RS<br />
(slender); carpus inferior margin with 0 RS; propodus<br />
.8 times as long as proximal width, inferior margin<br />
with 0 RS, propodal palm with small distal lobe, dac-
Figure 34. Aegapheles alazon (Bruce, 2004). A, dorsal view; B, head, dorsal view; C, frons; D, pereopod ; E, pereopod 2 (distal<br />
articles); F, pereopod 7; G, antennule; H, pleopods ; I, uropod.<br />
tylus smoothly curved, .2 as long as propodus. Pereopod<br />
2 merus inferior margin with 5 RS (set as 3 and 2),<br />
set as two groups. Pereopods 5–7 inferior margins <strong>of</strong><br />
ischium–carpus with long acute RS. Pereopod 3 similar<br />
to pereopod 2. Pereopod 7 basis 2.9 times as long as<br />
greatest width, inferior margins with 6 palmate setae<br />
(or more); ischium 0.6 as long as basis, inferior margin<br />
67<br />
with 5 RS (set as , 2 and 2), superior distal angle with<br />
7 RS, inferior distal angle with 5 RS; merus 0.9 as long<br />
as ischium, 2.6 times as long as wide, inferior margin<br />
with 4 RS (set as , and 2), superior distal angle with<br />
5 RS, inferior distal angle with 7 RS; carpus 0.8 as long<br />
as ischium, 2.9 times as long as wide, inferior margin<br />
with 2 RS (set as single cluster), superior distal angle
with 3 RS, inferior distal angle with 6 RS; propodus 0.7<br />
as long as ischium, 5.8 times as long as wide, inferior<br />
margin with 2 RS (set as single cluster), superior distal<br />
angle with 2 slender setae ( simple, palmate), inferior<br />
distal angle with 2 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by 5% <strong>of</strong> sternal width.<br />
Uropod peduncle posterior lobe about one-half<br />
as long as endopod. Uropod rami with apices narrowly<br />
rounded. Endopod apically not bifid, lateral<br />
margin proximally convex, with prominent excision,<br />
positioned about four-fifths along ramus, proximal<br />
lateral margin with 2 RS, distal lateral margin with 4<br />
RS, mesial margin strongly convex, with 3 RS. Exopod<br />
extending beyond end <strong>of</strong> endopod, 2.8 times as long as<br />
greatest width, apically not bifid; lateral margin weakly<br />
convex, with 0 RS; mesial margin sinuate, proximally<br />
concave, with 3 RS.<br />
size: Previously recorded to 58 mm, the size <strong>of</strong> the<br />
largest specimen examined here.<br />
Variation: Most specimens examined were in relatively<br />
poor condition, and the indicative range for robust<br />
setae on the pleotelson appears to be from 5+5 to 7+7.<br />
Uropod exopod (n = 9) mesial margin with 3–5, with<br />
3 (74%) or 4 (2 %) most frequent, lateral margin with<br />
9– 2 RS with 0 (84%) most frequent; uropod endopod<br />
mesial margin (n = 2 ) with 4 (7 %) or 5 ( 9%) most<br />
frequent (7 occurred on one specimen only), the lateral<br />
margin (n = 20) with the proximal RS at 2–4, distal RS 3<br />
or 4, with 3+4 (52%), 2+3 ( 4%) and 3+3 most frequent<br />
(4+3, 4+4 and 2+4 all occurred once).<br />
Pereopod setation <strong>of</strong> the merus is highly consistent<br />
across its range with +2 RS (95%), 2+2 occurring<br />
only once; pereopod 2 merus with 4+2 RS (73%) or 3+2<br />
(27%) most frequent and pereopod 3 with 4+2 (95%)<br />
most frequent (not included are NMNZ Cr.9265 and<br />
AK 84218, identified after counts were made).<br />
This is less variation, particularly for the uropodal<br />
endopod, than was recorded by Bruce (2004a) for the<br />
species across its entire range, suggesting that if good<br />
data can be obtained, consistent regional variation or<br />
cryptic species may be found to exist.<br />
remarks: Aegapheles alazon is most similar to A. birubi,<br />
the differentiating characters being that A. alazon has<br />
medially united eyes and lacks a propodal lobe on the<br />
palm <strong>of</strong> pereopods –3. Most <strong>of</strong> the NIWA specimens<br />
are in poor condition, several having dried out at some<br />
time in the past. In many <strong>of</strong> the specimens the eyes<br />
seem to have shrunk and drawn away from the cuticle,<br />
making it impossible to see if the eyes are medially<br />
united. All specimens identified here lack a significant<br />
propodal lobe on the palm <strong>of</strong> pereopods –3.<br />
68<br />
prey: In <strong>New</strong> <strong>Zealand</strong> — Carangidae, probably Trachurus<br />
novaezelandiae Richardson, 843 [as horse mackerel];<br />
Serranidae [as groper]; Rexea solandri (Cuvier, 832)<br />
(Gempylidae) [as gemfish]; Triglidae [as gurnard];<br />
and Ophidiidae, probably Genypterus blacodes (Forster,<br />
80 ) [as ling].<br />
distribution: Throughout <strong>New</strong> <strong>Zealand</strong> waters, extending<br />
north to <strong>New</strong> Caledonia, and northeast to Tonga.<br />
Previously recorded (Bruce 2004a) from South Africa<br />
(type locality), Tristan da Cunha, Seychelles, St Paul<br />
Is., southeastern Australia. Maximum recorded depth<br />
550 metres.<br />
Aegapheles birubi (Bruce, 2004) comb. nov. (Fig. 35)<br />
Aega birubi Bruce, 2004: 166, figs 18–21, 63.– Poore, 2005: 7.<br />
materiaL examined: ♀ (non-ovig. 34 mm), Cook Strait,<br />
4 February 2000, 65 m, from cheek <strong>of</strong> barracouta,<br />
coll. Pierce Black (NMNZ Cr.9949). Manca ( .5 mm),<br />
outside Wellington Harbour, 20 May 979, on pectoral<br />
fin <strong>of</strong> Polyprion oxygeneios, coll. C. Roberts (AK 4978).<br />
diagnosis (from Bruce 2004a): Eyes large, not medially<br />
united, separated by less than % width <strong>of</strong> head;<br />
eye colour pale brown. Pleonite 4 with posterolateral<br />
margins extending clearly beyond posterior margin<br />
<strong>of</strong> pleonite 5; pleonite 5 with posterolateral angles<br />
overlapped by lateral margins <strong>of</strong> pleonite 4. Pleotelson<br />
.0 as long as anterior width, dorsal surface without<br />
longitudinal carina; lateral margins convex, smooth,<br />
posterior margin with 6–8 RS.<br />
Antennule peduncle article 2 without anterodistal<br />
lobe; flagellum extending to mid-point <strong>of</strong> eye. Antenna<br />
peduncle article 2 inferior surface without distinct<br />
longitudinal suture; flagellum extending to middle <strong>of</strong><br />
pereonite .<br />
Frontal lamina flat, as wide as long, lateral margins<br />
converging posteriorly, anterior margin rounded, with<br />
small median point, posterior margin not abutting<br />
clypeus.<br />
Pereopod 1 merus inferior margin with 3 RS, set as<br />
two groups (<strong>of</strong> and 2), superior distal angle with 2<br />
RS (acute); carpus 0.5 as long as merus, inferior margin<br />
with 0 RS (with small lobe); propodus .4 times as long<br />
as proximal width, inferior margin with 0 RS, propodal<br />
palm with large distal lobe, dactylus smoothly curved,<br />
.7 as long as propodus. Pereopod 2 merus inferior margin<br />
with 6 RS (distal 2 on low lobe), set as single row,<br />
superior distal margin with 2 acute RS; carpus similar<br />
in size to that <strong>of</strong> pereopod , inferodistal angle with<br />
0 RS. Pereopod 3 similar to pereopod 2. Pereopods 5–7<br />
inferior margins <strong>of</strong> ischium–carpus with long acute<br />
RS. Pereopod 7 basis 2.8 times as long as greatest width,<br />
inferior margins with 7 palmate setae; ischium 0.6 as
Figure 35. Aegapheles birubi (Bruce, 2004). A, dorsal view; B, head, dorsal view; C, frons; D, antenna peduncle; E, antennule;<br />
F, pereopod (distal articles); G, pereopod 2 (distal articles); H, pereopod 7; I uropod.<br />
69
long as basis, inferior margin with 6 RS (set as , , 3<br />
and ), superior distal angle with 3 RS, inferior distal<br />
angle with 3 RS; merus 0.8 as long as ischium, .9 times<br />
as long as wide, inferior margin with 4 RS (set as 2 and<br />
2), superior distal angle with 5 RS, inferior distal angle<br />
with 5 RS; carpus 0.8 as long as ischium, 2.6 times as<br />
long as wide, inferior margin with 2 RS (single cluster),<br />
superior distal angle with 4 RS, inferior distal angle<br />
with 6 RS; propodus 0.7 as long as ischium, 4.5 times as<br />
long as wide, inferior margin with 2 RS (single cluster),<br />
superior distal angle with 3 slender setae (2 simple,<br />
palmate), inferior distal angle with 4 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by % <strong>of</strong> sternal width.<br />
Uropod peduncle ventrolateral margin with RS,<br />
posterior lobe about two-thirds as long as endopod.<br />
Uropod rami with apices narrowly rounded. Endopod<br />
apically not bifid, lateral margin proximally convex<br />
and distally straight, with prominent excision, positioned<br />
about three-quarters along ramus, proximal<br />
lateral margin with 3 RS, distal lateral margin with 4<br />
RS, mesial margin weakly convex, with 5 RS. Exopod<br />
extending beyond end <strong>of</strong> endopod, 3.0 as long as greatest<br />
width, apically not bifid; lateral margin weakly<br />
convex, with 0 RS; mesial margin sinuate, proximally<br />
concave, with 4 RS.<br />
remarks: <strong>The</strong> characteristic setation <strong>of</strong> the anterior pereopods,<br />
which has the distal robust seta on the merus<br />
notably longer than the preceding robust seta, allows<br />
ready identification; the sub-rectangular frontal lamina,<br />
lack <strong>of</strong> a lobe on the propodal palm <strong>of</strong> pereopods –3,<br />
uropodal exopod not extending beyond the endopod<br />
and number <strong>of</strong> robust setae on the uropods are further<br />
characters by which the species can be identified.<br />
prey: Recorded from hapuku, Polyprion oxygeneios<br />
Schneider & Forster, 80 (Polyprionidae); previously<br />
from barracouta (Thyristes atun) (Bruce 2004a).<br />
distribution: From the eastern Australia coast at Broken<br />
Bay to Tasmania, eastwards to the Cook Strait; at<br />
depths between 20–73 metres.<br />
Aegapheles copidis sp. nov. (Figs 36, 37)<br />
materiaL examined: Holotype, ♂ (22 mm), West Norfolk<br />
Ridge, 34°37.20’S, 68°57.03’E, 3 June 2003, 52 –539 m,<br />
coll. NORFANZ, RV Tangaroa (NIWA 23768).<br />
desCription: Body 2.3 times as long as greatest width,<br />
dorsal surfaces polished in appearance or sparsely<br />
punctate, widest at pereonite 5, lateral margins weakly<br />
ovate. Rostral point projecting anteriorly, not ventrally<br />
folded. Eyes large, medially united, anterior clear field<br />
70<br />
20% length <strong>of</strong> head, posterior clear field 27% length<br />
<strong>of</strong> head; each eye made up <strong>of</strong> ~20 transverse rows <strong>of</strong><br />
ommatidia, each row with ~ 3 ommatidia; eye colour<br />
dark brown. Pereonite 1 and coxae 2–3 each with posteroventral<br />
angle right-angled. Coxae 5–7 with entire oblique<br />
carina; posterior margins concave, posterolateral<br />
angle acute (less than 45°). Pleon with pleonite largely<br />
concealed by pereonite 7; pleonite 4 with posterolateral<br />
margins extending to but not beyond posterior margin<br />
<strong>of</strong> pleonite 5; pleonite 5 with posterolateral angles free,<br />
not overlapped by lateral margins <strong>of</strong> pleonite 4. Pleotelson<br />
.0 times as long as anterior width, dorsal surface<br />
without longitudinal carina; lateral margins sinuate,<br />
smooth, posterior margin with 8 RS.<br />
Antennule peduncle article 2 without anterodistal<br />
lobe; flagellum extending to mid-point <strong>of</strong> eye. Antenna,<br />
flagellum extending to posterior <strong>of</strong> pereonite 1.<br />
Frontal lamina flat, wider than long, lateral margins<br />
converging posteriorly, anterior margin rounded,<br />
without small median point, posterior margin not<br />
abutting clypeus.<br />
Maxilliped endite with apical setae; palp article 2<br />
with 3 RS; article 3 with 3 recurved RS; article 4 with 5<br />
hooked RS (4 large and small); article 5 articulating<br />
with article 4, longer than wide, sub-rectangular, with<br />
4 RS (all straight).<br />
Pereopod 1 basis 2.0 times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 2 RS; merus inferior<br />
margin with 3 RS, set as distal group, superior distal<br />
angle with 2 RS; carpus 0.4 as long as merus, inferior<br />
margin with 0 RS; propodus .2 times as long as proximal<br />
width, inferior margin with 0 RS, propodal palm<br />
with wide blade, dactylus smoothly curved, 2.2 as long<br />
as propodus. Pereopod 2 ischium inferior margin with<br />
RS, superior distal margin with 2 RS; merus inferior<br />
margin with 8 RS, set as two rows (distal paired rows<br />
<strong>of</strong> 3+3), superior distal margin with 2 acute RS; carpus<br />
similar in size to that <strong>of</strong> pereopod , inferodistal angle<br />
with RS. Pereopod 3 similar to pereopod 2. Pereopods<br />
5–7 inferior margins <strong>of</strong> ischium–carpus with long acute<br />
RS. Pereopod 6 similar to pereopod 7. Pereopod 7 basis<br />
2.9 times as long as greatest width, inferior margins<br />
with 2 palmate setae; ischium 0.5 as long as basis,<br />
inferior margin with 7 RS (set , 2, and 3), superior<br />
distal angle with 4 RS, inferior distal angle with 4<br />
RS; merus 0.7 as long as ischium, 2.6 times as long<br />
as wide, inferior margin with 5 RS (set as , and 3),<br />
superior distal angle with 7 RS, inferior distal angle<br />
with 6 RS; carpus .0 as long as ischium, 3.4 times as<br />
long as wide, inferior margin with 4 RS (set as and<br />
3), superior distal angle with 7 RS, inferior distal angle<br />
with 8 RS; propodus 0.8 as long as ischium, 4 times as<br />
long as wide, inferior margin with 3 RS (set as and<br />
2), superior distal angle with slender seta, inferior<br />
distal angle with 3 RS.
Figure 36. Aegapheles copidis sp. nov. Holotype. A, dorsal view; B, lateral view; C, head, dorsal view; D, frons; E, pleotelson<br />
posterior margin, dorsal view; F, maxilliped palp, article 5; G, maxilliped; H, pereopod ; I, pereopod 2.<br />
Penes low tubercles; penial openings separated by<br />
% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod 2.7 times as long as wide, distally<br />
narrowly rounded with strongly oblique mesial margin,<br />
lateral margin weakly concave, mesial margin strongly<br />
convex, with PMS on distal one-third; endopod 2.6<br />
times as long as wide, distally subtruncate, lateral<br />
7<br />
margin weakly concave, with PMS on distal margin<br />
only, mesial margin with PMS on distal two-thirds;<br />
peduncle .2 times as wide as long, mesial margin with<br />
8 coupling hooks. Pleopod 2 appendix masculina with<br />
straight margins, 0.9 times as long as endopod, distally<br />
narrowly rounded. Exopods <strong>of</strong> pleopods –3 each with<br />
distolateral margin not digitate; endopods <strong>of</strong> pleopods
Figure 37. Aegapheles copidis sp. nov. Holotype. A, pereopod 7; B, pereopod 7, propodus and distal margin <strong>of</strong> carpus, mesial<br />
side; C, pleopod ; D, pleopod 2; E, uropod, in situ; H, uropod exopod, ventral view, in situ.<br />
3–5 each with distolateral point; pleopods 2–4 peduncle<br />
distolateral margin without prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about one-half as long as endopod.<br />
Uropod rami with apices narrowly rounded. Endopod<br />
apically not bifid, lateral margin proximally convex,<br />
with prominent excision, positioned about three-<br />
quarters <strong>of</strong> the way along ramus, proximal lateral margin<br />
with 2 RS, distal lateral margin with 4 RS, mesial<br />
margin weakly convex, with 6 RS. Exopod extending<br />
72<br />
to end <strong>of</strong> endopod, 3.9 times as long as greatest width,<br />
apically not bifid; lateral margin weakly convex, with<br />
RS; mesial margin sinuate, proximally concave,<br />
with 4 RS.<br />
femaLe: Not known.<br />
size: Holotype 22 mm, a mature male; females probably<br />
larger.
Variation: <strong>The</strong> left and right uropodal exopod lateral<br />
margins had 0 and robust setae. <strong>The</strong> robust setae on<br />
the posterior margin <strong>of</strong> the pleotelson are apparently<br />
uneven though some may be missing; the probable<br />
number is 0 or 2.<br />
remarks: Aegapheles copidis sp. nov. can be identified<br />
by the prominent propodal blade on pereopods –3 together<br />
with the wide frontal lamina, and the relatively<br />
slender uropodal endopod, the lateral margin <strong>of</strong> which<br />
is weakly excavate; in addition the pleotelson apex extends<br />
only a little way beyond the posterior <strong>of</strong> the uropods.<br />
<strong>The</strong>re are three species <strong>of</strong> Aegapheles that have a<br />
propodal blade on pereopods –3. Aegapheles copidis has<br />
a longer propodal blade, shorter uropod exopod and<br />
weakly excised endopod lateral margin in comparison<br />
to A. kixalles Bruce, 2004; A. musorstom Bruce, 2004 has<br />
a symmetrically ovate frontal lamina, wide uropodal<br />
exopod which is longer than the endopod, pereopods<br />
2 and 3 with a single continuous row <strong>of</strong> robust setae<br />
on the inferior margin <strong>of</strong> the merus, and longer robust<br />
setae on pereopods 5–7. <strong>The</strong> tropical Aegapheles trulla<br />
Bruce, 2004 has a similar number <strong>of</strong> robust setae on the<br />
merus <strong>of</strong> pereopods –3 but these are not arranged in<br />
two rows; in addition the uropodal endopod lateral<br />
margin is weakly excised, uropodal exopod is very<br />
wide and the frontal lamina sub-circular.<br />
<strong>The</strong> single specimen was minimally dissected as,<br />
though very recently collected, high-grade absolute<br />
ethanol preservation had rendered it exceptionally<br />
brittle and fragile. <strong>The</strong> antenna, antennule, mouthparts<br />
and pleopods show few differences at species level, and<br />
direct observation suggests that these appendages are<br />
generally similar to those <strong>of</strong> other species <strong>of</strong> the genus<br />
(see Bruce 2004a).<br />
prey: No records.<br />
distribution: West Norfolk Ridge, northeastern <strong>New</strong><br />
<strong>Zealand</strong>; 52 –539 metres.<br />
etymoLogy: <strong>The</strong> epithet is the Latin copidis (cleaver,<br />
kitchen knife), alluding to the wide propodal blade on<br />
the anterior pereopods.<br />
Aegapheles hamiota (Bruce, 2004) comb. nov.<br />
(Fig. 38)<br />
Aega hamiota Bruce, 2004: 171, figs 22–25, 63.– Poore, 2005:<br />
7.<br />
materiaL examined: ♂ (24 mm), West Norfolk Ridge,<br />
32°36.49’S, 67°43.98’E, 29 May 2003, 699–707 m, coll.<br />
NORFANZ, RV Tangaroa (NIWA 23769).<br />
73<br />
diagnosis (from Bruce 2004a): Eyes large, medially<br />
united, anterior clear field 9% length <strong>of</strong> head, posterior<br />
clear field 34% length <strong>of</strong> head; eye colour dark brown.<br />
Pleonite 4 with posterolateral margins extending clearly<br />
beyond posterior margin <strong>of</strong> pleonite 5; pleonite 5 with<br />
posterolateral angles overlapped by lateral margins<br />
<strong>of</strong> pleonite 4. Pleotelson . times as long as anterior<br />
width, dorsal surface without longitudinal carina;<br />
lateral margins convex, smooth, posterior margin with<br />
– 6 RS.<br />
Antennule peduncle article 2 without anterodistal<br />
lobe; flagellum extending to mid-point <strong>of</strong> eye. Antenna<br />
peduncle article 2 inferior surface with distinct<br />
longitudinal suture; flagellum extending to posterior<br />
<strong>of</strong> pereonite .<br />
Frontal lamina flat, longer than greatest width, oval,<br />
anterior margin rounded, without small median point,<br />
posterior margin not abutting clypeus.<br />
Pereopod 1 merus inferior margin with 5 RS, set as<br />
three groups (<strong>of</strong> , 2 and 2), superior distal angle with<br />
7 RS (long acute); carpus 0.6 as long as merus, inferior<br />
margin with 0 RS; propodus .9 times as long as proximal<br />
width, inferior margin with 0 RS, propodal palm<br />
with small distal lobe, dactylus smoothly curved, .3<br />
as long as propodus. Pereopod 2 merus inferior margin<br />
with 9 RS, set as single row (with separation <strong>of</strong> distal<br />
2), superior distal margin with 5 acute RS (long acute);<br />
carpus longer than that <strong>of</strong> pereopod , with inferodistal<br />
lobe, inferodistal angle with RS. Pereopod 3 similar<br />
to pereopod 2. Pereopods 5–7 inferior margins <strong>of</strong> ischium–carpus<br />
with long acute RS, superior margins <strong>of</strong><br />
ischium–carpus with long, stiff, acute setae. Pereopod<br />
7 basis 3.3 times as long as greatest width, inferior<br />
margins with 6 palmate setae; ischium 0.5 as long as<br />
basis, inferior margin with 0 RS (7 long and 3 short),<br />
superior distal angle with 0 RS (5 stout lateral and 5<br />
long mesial), inferior distal angle with 0 RS (6 stout<br />
lateral and 4 long mesial); merus .0 as long as ischium,<br />
.9 times as long as wide, inferior margin with 5 RS (9<br />
long and 6 short), superior distal angle with 5 RS (3<br />
stout lateral and 2 long mesial), inferior distal angle<br />
with 8 RS (3 stout lateral and 5 long mesial); carpus .0<br />
as long as ischium, 3.0 as long as wide, inferior margin<br />
with 0 RS (7 long and 3 short), superior distal angle<br />
with 0 RS (3 stout lateral and 7 long mesial), inferior<br />
distal angle with RS (4 stout lateral and 7 long mesial);<br />
propodus 0.8 as long as ischium, 3.5 times as long<br />
as wide, inferior margin with 6 RS (4 long and 2 short),<br />
superior distal angle with 6 slender setae ( plumose),<br />
inferior distal angle with 4 RS.<br />
Penes low tubercles; penial openings separated by<br />
6% <strong>of</strong> sternal width.<br />
Uropod peduncle ventrolateral margin with 2 RS<br />
(short), posterior lobe about three-quarters as long as
Figure 38. Aegapheles hamiota (Bruce, 2004). A, dorsal view; B, head, dorsal view; C, frons; D, antennule; E, pereopod (distal<br />
articles); F, pereopod 2 (distal articles); G, pereopod 7; H, uropod; I, pleopod .<br />
74
endopod. Uropod rami with apices broadly rounded.<br />
Endopod apically not bifid, lateral margin proximally<br />
convex and distally convex, with prominent excision,<br />
positioned about two-thirds along ramus, proximal<br />
lateral margin with 6 RS, distal lateral margin with 9<br />
RS, mesial margin weakly convex, with 4 RS. Exopod<br />
extending beyond end <strong>of</strong> endopod, .9 times as long as<br />
greatest width, apically not bifid; lateral margin weakly<br />
convex, with RS; mesial margin convex, with 4 RS.<br />
remarks: Agrees well with the description given by<br />
Bruce (2004a). <strong>The</strong> species is immediately recognised<br />
by the united eyes, broad uropodal exopod and,<br />
uniquely within Aegapheles, by pereopods 6 and 7<br />
with long setae on the superior margin <strong>of</strong> the merus<br />
and carpus.<br />
distribution: <strong>New</strong>ly recorded from the vicinity <strong>of</strong> <strong>New</strong><br />
<strong>Zealand</strong>; previously <strong>New</strong> Caledonia and Queensland.<br />
At depths between approximately 500 to 700 metres.<br />
Aegapheles mahana sp. nov. (Figs 39–4 )<br />
materiaL examined: All material from <strong>New</strong> <strong>Zealand</strong>.<br />
Holotype: ♂ (37 mm), Rumble V Sea Mount,<br />
36°8.38’S, 78° .77’E, 23 May 200 , 603–365 m, coll.<br />
RV Tangaroa (NIWA 7939).<br />
Paratypes: Broken (pleon and pleotelson missing,<br />
estimated size 6 cm), Rumble V Sea Mount, 36°8.35’S,<br />
78° .74’E, 24 May 200 , 520–367 m, coll. RV Tangaroa<br />
(NIWA 7940). ♀ (non-ovig., broken c. 35 mm, dissected),<br />
Rumble V Sea Mount, 36°8.07’S, 78° 2.07’E,<br />
24 May 200 , 40–698 m, coll. RV Tangaroa (NIWA<br />
794 ).<br />
Additional material. ♂ (3 mm), vicinity <strong>of</strong> Chatham<br />
Island, 43°04.00’S, 78°38.99’W, 3 September 963, 549<br />
m, stn A09 0 (NIWA 7942). Manca (22 mm), South<br />
Norfolk Ridge, 33°22.6 ’S, 70° 2.70’E, June 2003,<br />
5 4–540 m, coll. NORFANZ, RV Tangaroa (NIWA<br />
7943). ♀ (non-ovig. 28 mm), South Norfolk Ridge,<br />
33°20.5 ’S, 70° 3.98’E, June 2003, 6 4–675 m, coll.<br />
NORFANZ, RV Tangaroa (NIWA 7944). ♀ (non-ovig.<br />
35 mm; all anterior pereopods broken), South Norfolk<br />
Ridge, 33°23.60’S, 70° 2.38’E, June 2003, 469–490 m,<br />
coll. NORFANZ, RV Tangaroa (NIWA 7945). 2 ♂ (32,<br />
42 mm), West Norfolk Ridge, 34°37.20’S, 68°57.03’E,<br />
3 June 2003, 52 –539 m, coll. NORFANZ, RV Tangaroa<br />
(NIWA 7946).<br />
desCription: Body 3.0 times as long as greatest width,<br />
dorsal surfaces polished in appearance and sparsely<br />
punctate, widest at pereonite 4, lateral margins subparallel.<br />
Rostral point projecting anteriorly, not ventrally<br />
folded. Eyes large, medially united, anterior clear field<br />
8% length <strong>of</strong> head, posterior clear field 35% length<br />
<strong>of</strong> head; each eye made up <strong>of</strong> ~22 transverse rows <strong>of</strong><br />
75<br />
ommatidia, each row with ~ 0 ommatidia; eye colour<br />
dark brown. Pereonite 1 and coxae 2–3 each with posteroventral<br />
angle rounded. Coxae 5–7 with entire oblique<br />
carina; posterior margins concave, posterolateral angle<br />
acute (less than 45°). Pleon with pleonite visible in<br />
dorsal view; pleonite 4 with posterolateral margins<br />
extending clearly beyond posterior margin <strong>of</strong> pleonite<br />
5; pleonite 5 with posterolateral angles overlapped by<br />
lateral margins <strong>of</strong> pleonite 4. Pleotelson .0 as long as<br />
anterior width, dorsal surface with longitudinal carina;<br />
lateral margins sinuate, smooth, posterior margin with<br />
6 RS.<br />
Antennule peduncle article 2 without anterodistal<br />
lobe; articles 3 and 4 0.7 times as long as combined<br />
lengths <strong>of</strong> articles and 2, article 3 3.2 times as long as<br />
wide; flagellum with 11 articles, extending to posterior<br />
margin <strong>of</strong> eye. Antenna peduncle article 2 inferior<br />
surface with distinct longitudinal suture; article 4 .9<br />
times as long as wide, .2 times as long as combined<br />
lengths <strong>of</strong> articles –3, without deep longitudinal<br />
groove, inferior margin 0 plumose setae, and short<br />
simple seta; article 5 0.9 times as long as article 4, 2.4<br />
times as long as wide, inferior margin with 2 palmate<br />
setae, anterodistal angle with cluster <strong>of</strong> 4 short simple<br />
setae; flagellum with 16 articles, extending to middle<br />
<strong>of</strong> pereonite .<br />
Frontal lamina flat, longer than greatest width, lateral<br />
margins converging posteriorly, anterior margin<br />
rounded, without small median point, posterior margin<br />
not abutting clypeus.<br />
Mandible molar process present, minute; palp article<br />
2 with 6 distolateral setae (plus scattered small simple<br />
setae), palp article 3 with 24 setae. Maxillule with 8 terminal<br />
RS (large, falcate). Maxilla mesial lobe with 2 RS<br />
( hooked, straight); lateral lobe with 3 RS (hooked).<br />
Maxilliped endite with apical seta; palp article 2 with<br />
2 RS ( small); article 3 with 5 recurved RS (plus<br />
distal serrate slender seta); article 4 with 5 hooked RS<br />
(4 large, small); article 5 articulating with article 4,<br />
distally convex, with 3 RS (2 curved, straight, plus 3<br />
stiff slender setae).<br />
Pereopod 1 basis 2 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin<br />
with 0 RS; merus inferior margin with 5 RS, set as<br />
two groups (<strong>of</strong> 3 and 2), superior distal angle with 2<br />
RS; carpus 0.4 as long as merus, inferior margin with<br />
0 RS; propodus .7 times as long as proximal width,<br />
inferior margin with 0 RS, propodal palm with small<br />
distal lobe, dactylus smoothly curved, .5 as long as<br />
propodus. Pereopod 2 ischium inferior margin with 2<br />
RS; merus inferior margin with 6 RS, set as single row<br />
(weak separation <strong>of</strong> 4 and 2), superior distal margin<br />
with 2 acute RS; carpus longer than that <strong>of</strong> pereopod<br />
, with inferodistal lobe, inferodistal angle with RS.<br />
Pereopod 3 similar to pereopod 2. Pereopods 5–7 inferior<br />
margins <strong>of</strong> ischium–carpus with long acute RS.
Figure 39. Aegapheles mahana sp. nov. Holotype, A–G, H–I, paratype NIWA 794 . A, dorsal view; B, lateral view; C, pleonites,<br />
lateral margin; D, head, dorsal view; E, frons; F, pleotelson, posterior margin, dorsal view; G, sternite 7 showing penial papillae;<br />
H, antennule; I, antenna; J, antenna peduncle articles –3, ventral view.<br />
Pereopod 6 similar to pereopod 7. Pereopod 7 basis 2.9<br />
times as long as greatest width, inferior margins with<br />
3 palmate setae; ischium 0.6 as long as basis, inferior<br />
margin with 7 RS (loosely set , 3 and 3), superior distal<br />
angle with 3 RS, inferior distal angle with 4 RS; merus<br />
76<br />
0.8 as long as ischium, 2.4 times as long as wide, inferior<br />
margin with 6 RS (as 2 close-set groups <strong>of</strong> 3 and<br />
3), superior distal angle with 8 RS, inferior distal angle<br />
with 7 RS; carpus .0 as long as ischium, 3. times as<br />
long as wide, inferior margin with 5 RS (set as and
Figure 40. Aegapheles mahana sp. nov. Paratype NIWA 794 . A, left mandible (broken); B, right mandible (broken) C, mandible<br />
palp article 3; D, maxillule apex; E, maxilla apex; F, maxilliped articles 2–5; G, maxilliped article 5 (Leica).<br />
2), superior distal angle with 6 RS, inferior distal angle<br />
with 6 RS; propodus 0.7 as long as ischium, 4.5 times as<br />
long as wide, inferior margin with 4 RS (set as 2 and 2),<br />
superior distal angle with small slender seta, inferior<br />
distal angle with 3 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by 6% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod 2.2 times as long as wide, distally<br />
narrowly rounded with strongly oblique mesial mar-<br />
77<br />
gin, lateral margin weakly concave, mesial margin<br />
strongly convex, with PMS on distal half; endopod<br />
2.3 times as long as wide, distally subtruncate, lateral<br />
margin weakly concave, with PMS on distal half, mesial<br />
margin with PMS on distal two-thirds; peduncle .4<br />
times as wide as long, mesial margin with coupling<br />
hooks. Pleopod 2 appendix masculina with straight margins,<br />
0.7 times as long as endopod, distally narrowly<br />
rounded. Exopods <strong>of</strong> pleopods –3 each with distola-
Figure 41. Aegapheles mahana sp. nov. Holotype, except G and H, paratype NIWA 794 . A–C, pereopods , 2 and 7 respectively;<br />
D, pleopod ; E, pleopod 2; F, uropod; G, uropod; H, uropod exopod, ventral view.<br />
78
teral margin not digitate; endopods <strong>of</strong> pleopods 3–5<br />
each with distolateral point; pleopods 2–4 peduncle<br />
distolateral margin without prominent acute RS.<br />
Uropod peduncle ventrolateral margin with RS,<br />
posterior lobe about one-half as long as endopod.<br />
Uropod rami apices narrowly rounded. Endopod<br />
apically not bifid, lateral margin proximally convex<br />
and distally straight, with prominent excision, positioned<br />
about three-quarters along ramus, proximal<br />
lateral margin with 6 RS, distal lateral margin with 4<br />
RS, mesial margin weakly convex, with 3 RS. Exopod<br />
extending beyond end <strong>of</strong> endopod, 3.5 times as long<br />
as greatest width, apically not bifid; lateral margin<br />
straight, with 9 RS; mesial margin sinuate, proximally<br />
concave, with 4 RS.<br />
femaLe: Similar to the male, but for the sexual characters;<br />
no ovigerous females present.<br />
size: Males 3 to 42 mm (mean = 35.5); female 28 to 25<br />
mm; single manca (with pereopod 7 present but undeveloped)<br />
22 mm; from a large damaged specimen the<br />
species may reach an estimated size <strong>of</strong> up to 6 cm.<br />
Variation: All four type specimens have received at<br />
least some damage so it is not possible to quantify the<br />
range in number <strong>of</strong> the robust setae. <strong>The</strong> shape and<br />
proportion <strong>of</strong> the uropod rami is consistent between<br />
specimens. Pleotelson RS potentially ranging from 4<br />
to 8 (counted as 8+9, 7+9). Uropod exopod mesial<br />
margin with 4 or 5 RS, lateral margin 8 or 9; uropod<br />
endopod mesial margin varied from 3 to 6 RS, lateral<br />
margin 6+3–6, once 7+5.<br />
<strong>The</strong> robust setae on the merus <strong>of</strong> pereopods –3<br />
present a constant pattern, with pereopod with 3+2<br />
(all) pereopod 2 4+2 (all) and pereopod 3 merus had<br />
4+2, 4+3 and 5+2; in the largest specimen (NIWA 7940)<br />
the RS on pereopod 3 form a continuous row.<br />
Fresh colour is a rich orange–brown, with dark<br />
brown eyes; chromatophores not apparent. A conspicuous<br />
feature <strong>of</strong> all the freshly caught specimens was the<br />
dark brown colour <strong>of</strong> the robust setae on the pereopods,<br />
uropodal rami and pleotelson.<br />
remarks: Aegapheles mahana sp. nov. can be identified<br />
by the following combination <strong>of</strong> characters: large eyes,<br />
entirely united and extending almost fully to the anterior<br />
margin <strong>of</strong> the head; anteriorly rounded frontal<br />
lamina; pereopods –3 propodus palm without blade,<br />
with small distal lobe; inferior margin <strong>of</strong> pereopod<br />
merus with 3+2 robust setae, that <strong>of</strong> pereopods 2 and<br />
3 with 4+2 robust setae; and the relatively slender and<br />
distally acute uropodal rami with the exopod extending<br />
distinctly beyond the posterior <strong>of</strong> the endopod; and<br />
the lateral margin <strong>of</strong> the uropodal endopod with 6 or 7<br />
setae proximal (or anterior) to the lateral notch.<br />
79<br />
Similar species in the region include Aegapheles<br />
alazon Bruce, 2004, A. kixalles Bruce, 2004 and A. warna<br />
Bruce, 2004. Aegapheles kixalles, known from the vicinity<br />
<strong>of</strong> <strong>New</strong> Caledonia, is most similar in eye size and<br />
shape, and in the shape and setation <strong>of</strong> the pereopods<br />
and uropods, but is immediately distinguished by<br />
having a narrow propodal blade on pereopods –3.<br />
Aegapheles alazon is a widely distributed and sympatric<br />
species which differs in having smaller eyes with<br />
a much larger anterior clear field, pereopod 1 usually<br />
with +2 robust setae on the merus (compared to 3+2)<br />
and the uropodal endopod has only 2–4 robust setae<br />
anterior to the lateral notch; in addition the lateral margins<br />
<strong>of</strong> the frontal lamina are more strongly convex in<br />
A. alazon. Aegapheles warna presents a similar pattern <strong>of</strong><br />
setation to the uropods, but the robust setae are larger,<br />
more numerous and the exopod is wider than that <strong>of</strong><br />
A. mahana, extending only slightly beyond the endopod<br />
while in A. mahana the exopod extends well beyond<br />
the endopod apex and is noticeably more slender (3.5<br />
times as long as wide) than that <strong>of</strong> A. warna (2.9 times<br />
as long as wide).<br />
prey: No records.<br />
distribution: Northeastern <strong>New</strong> <strong>Zealand</strong> and in the<br />
vicinity <strong>of</strong> the Chatham Islands; potentially at depths<br />
between 365 and 40 metres (some dredge samples<br />
covered substantial depth ranges up the sides <strong>of</strong> the<br />
seamounts).<br />
etymoLogy: <strong>The</strong> epithet, mahana, is a Mäori word<br />
meaning warm, in reference to the volcanic activity <strong>of</strong><br />
these sea mounts.<br />
Aegapheles rickbruscai (Bruce, 2004) comb. nov.<br />
(Fig. 42)<br />
Aega rickbruscai Bruce, 2004: 196, figs 40–44, 64.<br />
materiaL examined: ♂ (44 mm), Cavalli Sea Mount, <strong>of</strong>f<br />
northern North Island, 34°07.2 ’S, 74°05.64’E, 6 April<br />
2002, 554–549 m, NZOI stn Z 055, coll. S. O’Shea on<br />
RV Kaharoa (NIWA 3444).<br />
diagnosis (from Bruce 2004a): Eyes large, medially<br />
united, anterior clear field 11% length <strong>of</strong> head, posterior<br />
clear field 34% length <strong>of</strong> head; eye colour dark brown.<br />
Pleonite 4 with posterolateral margins extending clearly<br />
beyond posterior margin <strong>of</strong> pleonite 5; pleonite 5 with<br />
posterolateral angles overlapped by lateral margins <strong>of</strong><br />
pleonite 4. Pleotelson 0.9 times as long as anterior width,<br />
dorsal surface without longitudinal carina; lateral margins<br />
convex, smooth, posterior margin with 0 RS.<br />
Antennule peduncle article 2 without anterodistal<br />
lobe; flagellum extending to mid-point <strong>of</strong> eye. An-
Figure 42. Aegapheles rickbruscai (Bruce, 2004). A, dorsal view; B, head, dorsal view; C, frons; D, maxilliped palp article 5;<br />
E, maxilla apex; F, pereopod (distal articles); G, pereopod 2 (distal articles); H, pereopod 7; I, antennule; J, uropod.<br />
80
tenna peduncle article 2 inferior surface with distinct<br />
longitudinal suture; flagellum extending to posterior<br />
<strong>of</strong> pereonite .<br />
Frontal lamina flat, longer than greatest width, oval<br />
(rounded-ovate in shape), anterior margin rounded,<br />
with small median point, posterior margin not abutting<br />
clypeus.<br />
Pereopod 1 merus inferior margin with 8 RS, set as<br />
two groups (<strong>of</strong> 6 and 2), superior distal angle with 6<br />
RS (long acute); carpus 0.5 as long as merus, inferior<br />
margin with 0 RS (with distal flange); propodus 1.3<br />
times as long as proximal width, inferior margin with<br />
0 RS, propodal palm with large distal lobe, dactylus<br />
smoothly curved, .6 as long as propodus. Pereopod 2<br />
merus inferior margin with 2 RS, set as single row,<br />
superior distal margin with acute RS; carpus similar<br />
in size to that <strong>of</strong> pereopod , inferodistal angle with<br />
0 RS. Pereopod 3 similar to pereopod 2. Pereopods 5–7<br />
inferior margins <strong>of</strong> ischium–carpus with long acute<br />
RS. Pereopod 7 basis 3.2 times as long as greatest width,<br />
inferior margins with 4 palmate setae (mesial and lateral<br />
margins with 5 and 9 respectively); ischium 0.6 as<br />
long as basis, inferior margin with RS (set as row <strong>of</strong><br />
7 long and 5 short marginal RS), superior distal angle<br />
with 7 RS, inferior distal angle with 9 RS (2 long and 7<br />
short); merus 0.8 as long as ischium, .7 times as long<br />
as wide, inferior margin with 8 RS (3 long acute and<br />
short RS and 4 short submarginal RS), superior distal<br />
angle with 8 RS, inferior distal angle with 7 RS (3 short,<br />
4 slender and long); carpus 0.9 as long as ischium, 2.8<br />
times as long as wide, inferior margin with 5 RS (set<br />
as 3 long and 2 short), superior distal angle with 9 RS,<br />
inferior distal angle with 0 RS; propodus 0.8 as long<br />
as ischium, 4.3 times as long as wide, inferior margin<br />
with 5 RS (3 long and 2 short), superior distal angle<br />
with 7 slender setae (simple and palmate, including<br />
RS), inferior distal angle with 4 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by 7% <strong>of</strong> sternal width.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about three-quarters as long as endopod.<br />
Uropod rami apices with endopod narrowly rounded<br />
and exopod broadly rounded. Endopod apically not<br />
bifid, lateral margin proximally convex, with prominent<br />
excision, positioned about three-quarters along<br />
ramus, proximal lateral margin with 2 RS, distal lateral<br />
margin with 5 RS, mesial margin weakly convex (appearing<br />
straight), with 5 RS. Exopod extending beyond<br />
end <strong>of</strong> endopod, 2.5 times as long as greatest width,<br />
apically not bifid; lateral margin weakly convex, with<br />
RS; mesial margin convex, with 6 RS.<br />
remarks: Aegapheles rickbruscai can be identified by the<br />
huge eyes, with very little anterior free space on the<br />
head, the ovate and anteriorly acute frontal lamina and<br />
the continuous row <strong>of</strong> 6+2 robust setae on the inferior<br />
8<br />
margin <strong>of</strong> pereopod , 2– 5 robust setae on the inferior<br />
margins <strong>of</strong> the merus <strong>of</strong> pereopods 2 and 3; pereopods<br />
5–7 have easily observed long robust setae, the<br />
uropodal exopod is relatively broad and the posterior<br />
margins <strong>of</strong> the pleotelson lack robust setae.<br />
<strong>The</strong> most similar species in <strong>New</strong> <strong>Zealand</strong> waters is<br />
A. hamiota, that species being readily distinguished by<br />
having long, acute robust setae on the superior margins<br />
<strong>of</strong> the ischium, merus and carpus <strong>of</strong> pereopods 5–7.<br />
A. musorstom, presently known only from the vicinity<br />
<strong>of</strong> <strong>New</strong> Caledonia but may occur more widely, is also<br />
similar but readily separated by the presence <strong>of</strong> a conspicuous<br />
blade on the propodus <strong>of</strong> pereopods –3.<br />
distribution: Previously known from the vicinity <strong>of</strong><br />
<strong>New</strong> Caledonia, the range is here extended southwards<br />
to <strong>New</strong> <strong>Zealand</strong>.<br />
Aegapheles umpara (Bruce, 2004) comb. nov.<br />
(Fig. 43)<br />
Aega umpara Bruce, 2004: 208, figs 49–52, 64.– Poore, 2005:<br />
7.<br />
materiaL examined: Specimen, vicinity <strong>of</strong> Kermadec<br />
Islands, 30.2530°S, 78.4033°W, 90 m, Challenger Centenary,<br />
May 994, stn K0837 (NIWA 3479). ♂ (29 mm),<br />
Lord Howe Rise, 3 °52.28’S, 59° 6.6 ’E, 23 May 2003,<br />
on Carcharhinus galapagensis, 68–9 m, coll. NORFANZ,<br />
RV Tangaroa (NIWA 23770).<br />
diagnosis (from Bruce 2004a): Eyes large, medially<br />
united, anterior clear field 17% length <strong>of</strong> head, posterior<br />
clear field 48% length <strong>of</strong> head; eye colour dark brown.<br />
Pleonite 4 with posterolateral margins extending clearly<br />
beyond posterior margin <strong>of</strong> pleonite 5; pleonite 5 with<br />
posterolateral angles free, not overlapped by lateral<br />
margins <strong>of</strong> pleonite 4. Pleotelson . times as long as<br />
anterior width, dorsal surface without longitudinal<br />
carina; lateral margins weakly convex (very), smooth,<br />
posterior margin with 8 RS.<br />
Antennule peduncle article 2 without anterodistal<br />
lobe; flagellum extending to posterior margin <strong>of</strong> eye.<br />
Antenna peduncle article 2 inferior surface without<br />
distinct longitudinal suture; flagellum extending to<br />
middle <strong>of</strong> pereonite 2.<br />
Frontal lamina flat, longer than greatest width,<br />
rectangular, anterior margin anteriorly truncate, with<br />
small median point, posterior margin not abutting<br />
clypeus.<br />
Pereopod 1 merus inferior margin with 3 RS, set as<br />
two groups (set as and 2), superior distal angle with<br />
RS (and 2 simple setae); propodus .3 times as long<br />
as proximal width, inferior margin with 0 RS, propodal<br />
palm simple, without blade or process, dactylus<br />
smoothly curved (but strongly recurved), .5 as long
Figure 43. Aegapheles umpara (Bruce, 2004). A, dorsal view; B, head, dorsal view; C, frons; D, pereopod ; antennule;<br />
E, pereopod 2 (distal articles); F, pereopod 7; G, antennule; H, uropod.<br />
as propodus. Pereopod 2 merus inferior margin with 5<br />
RS (set as 3 and 2), set as two groups, superior distal<br />
margin with 2 acute RS; carpus inferodistal angle<br />
with RS (minute). Pereopod 3 similar to pereopod<br />
2. Pereopods 5–7 inferior margins <strong>of</strong> ischium–carpus<br />
82<br />
with long acute RS. Pereopod 7 basis 3.5 times as long<br />
as greatest width, inferior margins with 6–8 palmate<br />
setae; ischium 0.5 as long as basis, inferior margin with<br />
6 RS (set as , 3, and ), superior distal angle with 5<br />
RS, inferior distal angle with 6 RS; merus 0.7 as long
as ischium, .8 times as long as wide, inferior margin<br />
with 5 RS (set as 2 and 3), superior distal angle with 6<br />
RS, inferior distal angle with 6 RS; carpus 0.9 as long<br />
as ischium, 3.3 times as long as wide, inferior margin<br />
with 2 RS (single cluster), superior distal angle with 4<br />
RS, inferior distal angle with 7 RS; propodus 0.7 as long<br />
as ischium, 4.2 times as long as wide, inferior margin<br />
with 2 RS, superior distal angle with 2 slender setae<br />
(palmate), inferior distal angle with 2 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by 3.3% <strong>of</strong> sternal width.<br />
Uropod peduncle ventrolateral margin with RS<br />
(and 3 short setae), posterior lobe about two-thirds<br />
as long as endopod. Uropod rami apices narrowly<br />
rounded. Endopod apically not bifid, lateral margin<br />
proximally convex (weakly), with prominent excision,<br />
positioned about two-thirds along ramus, proximal<br />
lateral margin with 4 RS, distal lateral margin with 5<br />
RS, mesial margin weakly convex, with 5 RS. Exopod<br />
not extending to end <strong>of</strong> endopod, 2.8 times as long as<br />
greatest width; lateral margin weakly convex, with 8<br />
RS; mesial margin sinuate, proximally concave, with<br />
4 RS.<br />
remarks: <strong>The</strong> characteristic setation <strong>of</strong> the anterior<br />
pereopods, which has the distal robust setae on the<br />
merus notably longer than the preceding robust seta,<br />
allows ready identification; the sub-rectangular frontal<br />
lamina, lack <strong>of</strong> a lobe on the propodal palm <strong>of</strong> pereopods<br />
–3, the uropodal exopod not extending beyond<br />
the endopod and number <strong>of</strong> robust setae on the uropods<br />
are further characters by which the species can<br />
be identified.<br />
prey: Previously recorded from bony fish, sharks and<br />
rays (Bruce 2004a); here recorded from the Galapagos<br />
shark, Carcharhinus galapagensis (Snodgrass & Heller,<br />
905) (Carcharhinidae).<br />
distribution: From eastern Australia at Moreton Bay<br />
and the Solitary Islands to Elizabeth Reef, Norfolk<br />
Island and the Kermadec Islands; here recorded from<br />
Lord Howe Rise; depth range is relatively shallow,<br />
from a few metres to 75 metres, considerably shallower<br />
than most related species.<br />
Genus Aegiochus Bovallius, 885<br />
Aegiochus Bovallius, 885: 4.<br />
Aega (Ramphion) Brusca, 983: .– Kensley & Schotte, 989:<br />
6 (key) [type species Aega plebeia Hansen, 897; by<br />
original designation].<br />
type speCies: Type species is Aega nordenskjoldii Bovallius,<br />
885, by monotypy; junior synonym <strong>of</strong> A. ventrosa<br />
M. Sars, 859. Four specimens <strong>of</strong> A. ventrosa (ZMUC)<br />
were examined.<br />
83<br />
diagnosis: Body moderately to strongly dorsally<br />
vaulted. Rostral point separating antennule peduncles,<br />
appearing truncate in dorsal view, ventrally or<br />
ventrally and posteriorly directed. Eyes present, <strong>of</strong>ten<br />
large, sometimes medially united. Pleonite not<br />
abruptly narrower than pereonite 7; pleonite 4 with<br />
lateral margins extending to or beyond posterior margin<br />
<strong>of</strong> pleonite 5. Antennule peduncle articles and 2<br />
cylindrical, not flattened or expanded. Maxillule with<br />
–3 large broad-based RS, several small RS. Maxilliped<br />
palp article 5 longer that wide, subtruncate, with long<br />
setae; endite present. Pereopods –3 merus inferior<br />
margin with small RS, usually set as two groups.<br />
desCription: Body moderately to strongly vaulted,<br />
about 2 to 4 times as long as wide. Head with eyes,<br />
<strong>of</strong>ten large, may meet at midpoint; anterior margin<br />
with median (rostral) point. Coxae 4–7 longer than<br />
respective segment, posteriorly produced. Pleon not<br />
abruptly narrower than pereon; pleonites all visible,<br />
not posteriorly widest, pleonite 5 laterally overlapped<br />
by pleonite 4.<br />
Frontal lamina present, with free posterior margin<br />
or with posterior stem. Mandible with uni- or bicuspid<br />
incisor; molar process present, reduced or absent. Maxillule<br />
with single large, flat, broad-based RS, several<br />
small RS. Maxilliped palp 5-articled, article shorter<br />
than wide, articles 3 and 4 each with 2–6 stout recurved<br />
RS; endite present, usually with –2 terminal setae.<br />
Uropod rami with endopod and exopod co-planar,<br />
rami extending to or slightly beyond pleotelson apex,<br />
marginal setae in single tier, apices acute.<br />
remarks: <strong>The</strong> genus Aegiochus was established by<br />
Bovallius ( 885) on the basis <strong>of</strong> a biological misinterpretation.<br />
<strong>The</strong> holotype <strong>of</strong> Aegiochus nordenskjoldii was<br />
an intermoult specimen, this fact being quickly recognised<br />
by subsequent workers (e.g. Hansen 890; Sars<br />
899) who placed the genus into synonymy with Aega.<br />
<strong>The</strong> species was also quickly recognised to be a junior<br />
synonym <strong>of</strong> Aegiochus ventrosa (as Aega ventrosa).<br />
Brusca ( 983) recognised that there were two large<br />
‘groups’ within Aega, and established the subgenera<br />
Rhamphion and the nominate subgenus. Brusca’s<br />
subgeneric assignments were based on direct examination<br />
<strong>of</strong> 23 species and literature for a further 30<br />
species (<strong>of</strong> a then total <strong>of</strong> about 60 species), with the<br />
primary basis for distinguishing the two subgenera<br />
being the presence (or absence) <strong>of</strong> expanded peduncular<br />
articles on the antennule, presence or absence<br />
<strong>of</strong> a distinct rostrum, and presence or absence <strong>of</strong><br />
recurved ‘spines’ (robust setae) on the ‘apex <strong>of</strong> the<br />
maxilliped palp’. Some workers initially followed<br />
the use <strong>of</strong> the subgenera (e.g. Bruce 983; Wetzer<br />
990; Kensley & Schotte 989) but later found that<br />
critical characters were inconsistently present within
the subgenera and could not apply the subgeneric<br />
concepts (e.g. Bruce 988, 996, 2004a; Kensley & Chan<br />
200 ). Notwithstanding, Brusca had recognised an<br />
evident division within the genus, one that the present<br />
analysis supports and validates, albeit using different<br />
and rather more character states.<br />
Brusca ( 983) was apparently unaware <strong>of</strong> some<br />
<strong>of</strong> the several older and available names that were in<br />
synonymy with Aega. Aega plebeia, the type species <strong>of</strong><br />
Rhamphion falls within the genus concept and within<br />
the major clade that contains Aegiochus, so that name<br />
takes priority.<br />
Thirty-seven named species are included in the<br />
genus, those below, and those listed under ‘Species<br />
included ...’ (p.2 3).<br />
Key to the new <strong>Zealand</strong> species <strong>of</strong> Aegiochus<br />
Eyes large, medially united ..................................4<br />
– Eyes separate ..........................................................2<br />
2. Pleotelson entire .....................................................4<br />
– Pleotelson with quadrate median excision ........3<br />
3. Pereopod dactylus .4 times as long as propodus;<br />
coxae 5–7 dorsally weakly concave, posteriorly<br />
produced and acute ...................................<br />
................................................Aegiochus beri (p. 85)<br />
– Pereopod dactylus .0 times as long as propodus;<br />
coxae 5–7 dorsally convex, posteriorly<br />
weakly rounded ..............Aegiochus riwha (p. 4 )<br />
4. Rostrum folded ventrally and posteriorly, appearing<br />
truncate in dorsal view; in contact with or<br />
overlapping anterior margin <strong>of</strong> frontal lamina ...<br />
..................................................................................7<br />
– Rostrum ventral or ventrally directed, not folded<br />
posteriorly, appearing subtruncate or absent in<br />
dorsal view, not overlapping anterior margin <strong>of</strong><br />
frontal lamina .........................................................5<br />
5. Frontal lamina ventrally flat, pentagonal (‘cirolanid-like’);<br />
adult males with rostrum extended<br />
to form prominent process and pereonite with<br />
paired sub-lateral processes giving tri-horned appearance;<br />
eyes huge, largely occupying anterior<br />
margin <strong>of</strong> head ............Aegiochus vigilans (p. 50)<br />
– Frontal lamina short, with posterior stem, not<br />
ventrally flat; males and females without processes<br />
on head or pereonite ; eyes narrowly joined<br />
medially ...................................................................6<br />
6. Eye join medially narrowly (2 or 3 ommatidia);<br />
pereopods 2 and 3 carpus inferior margin distinctly<br />
lobed with prominent RS; average size<br />
approx. 4 mm .............Aegiochus bertrandi (p. 88)<br />
– Eye join moderately wide (3–5 ommatidia); pereopods<br />
2 and 3 carpus inferior margin not lobed,<br />
with only small RS; average size approx. mm<br />
............................................. Aegiochus coroo (p. 93)<br />
84<br />
7. Eyes separate ..........................................................9<br />
– Eyes medially united .............................................8<br />
8. Eyes moderate in size, occupying less than 40%<br />
head length; frontal lamina posterior margin<br />
concave; posteroventral angles <strong>of</strong> pereonite<br />
and coxae 2 and 3 acute, each with posteroventral<br />
point; appendix masculina straight. ......................<br />
............................................Aegiochus kakai (p. 09)<br />
– Eyes large in size, occupying more than 60% head<br />
length; frontal lamina posterior margin straight;<br />
posteroventral angles <strong>of</strong> pereonite and coxae 2<br />
and 3 truncate, each without posteroventral point;<br />
appendix masculina sinuate ...................................<br />
......................................... Aegiochus kanohi (p. 4)<br />
9. Frontal lamina with distinct posterior blade;<br />
pereopod dactylus as long as or longer than<br />
propodus ............................................................... 0<br />
– Frontal lamina without distinct posterior blade;<br />
pereopod dactylus slightly shorter (0.9) than<br />
propodus .................... Aegiochus insomnis (p. 03)<br />
0. Pereopods –3 without distinct, triangular distal<br />
blade .......................................................................<br />
– Pereopods –3 with distinct, triangular distal<br />
blade ...............................Aegiochus piihuka (p. 28)<br />
. Pleotelson without RS ......................................... 2<br />
– Pleotelson with 2 or more RS ............................. 3<br />
2. Adult size ‘small’ (average 7.7 mm males, about<br />
2.4 mm females); eyes separated by 7% head<br />
width; uropod endopod mesial margin usually<br />
with 3 RS; dactylus .4 times as long as propodus<br />
.............................. Aegiochus gordoni (p. 97)<br />
– Adult size ‘tiny’ (average 5.4 mm males, about 8.0<br />
mm females); eyes separated by 6% head width;<br />
uropod endopod mesial margin with or 2 RS;<br />
dactylus .2 times as long as propodus ................<br />
......................................Aegiochus nohinohi (p. 23)<br />
3. Eyes large, separated by less than 5% head width;<br />
pleotelson with 0 or more RS .......................... 4<br />
– Eyes small, separated by 32% head width; pleotelson<br />
with 2 RS ...............Aegiochus laevis (p. 20)<br />
4. Eyes separated by 2% head width; anterior<br />
pereopods stout, propodus .7 times as long as<br />
wide; coxae not conspicuous in dorsal view, not<br />
posteriorly produced ...............................................<br />
......................................Aegiochus pushkini (p. 34)<br />
– Eyes separated by 4% head width; anterior<br />
pereopods slender, propodus 2.9 times as long as<br />
wide; coxae conspicuous in dorsal view; coxae 6<br />
and 7 posteriorly produced, acute .........................<br />
............................................. Aegiochus tara (p. 46)
Aegiochus beri (Bruce, 983), comb. nov. (Figs 44, 45)<br />
Aega (Rhamphion) beri Bruce, 1983: 773, figs 11, 12.<br />
Aega beri.– Bruce, Lew Ton & Poore, 2002: 60.– Springthorpe<br />
and Lowry, 994: 39.<br />
materiaL examined: 3 ♂ (32, 23, 8 mm), imm. ♀ ( 9<br />
mm), east <strong>of</strong> Lord Howe Island, Tasman Sea, 30°3 .0–<br />
9.4’S, 6 °54.2–40.6’E, 29 December 975, 2 0 m, coll.<br />
J.E. Watson on RV Dmitry Mendeleev (NMV J8885).<br />
♂ (27 mm), 32°26.70’S, 6 °46.95’E, 25 May 2003,<br />
30– 47 m, coll. NORFANZ, R.V. Tangaroa (NIWA<br />
23766). ♀ (non-ovig. 2 mm), 34° 2. 8’S, 62°4 . 8’E, 26<br />
May 2003, 748–772 m, coll. NORFANZ, R.V. Tangaroa<br />
(NIWA 23767). ♀ ( 9 mm, non-ovig.), <strong>of</strong>f Queensland,<br />
7° 9’S, 47° ’E, 2 May 986, 406 m, stn 30-2, coll.<br />
CIDARIS (QM W 3393).<br />
type LoCaLity: East <strong>of</strong> Port Jackson, NSW, 33°38.36’S,<br />
52° 5.09’E, at a depth <strong>of</strong> 945–972 m.<br />
desCription: Body 2.9 times as long as greatest width,<br />
dorsal surfaces polished in appearance, sparsely<br />
punctate, widest at pereonite 5, lateral margins weakly<br />
ovate. Rostral point folded ventrally and posteriorly.<br />
Eyes large, not medially united, separated by about<br />
0% width <strong>of</strong> head; each eye made up <strong>of</strong> ~ 6 transverse<br />
rows <strong>of</strong> ommatidia, each row with ~8 ommatidia; eye<br />
colour dark brown. Pereonite 1 and coxae 2–3 each with<br />
posteroventral angle with small distinct produced<br />
point. Coxae 5–7 with entire oblique carina; posterior<br />
margins convex (weakly), posterolateral angle acute<br />
(less than 45°). Pleon with pleonite largely concealed<br />
by pereonite 7; pleonite 4 with posterolateral margins<br />
extending to but not beyond posterior margin <strong>of</strong> pleonite<br />
5; pleonite 5 with posterolateral angles free, not<br />
overlapped by lateral margins <strong>of</strong> pleonite 4. Pleotelson<br />
0.8 times as long as anterior width, dorsal surface<br />
without longitudinal carina; lateral margins convex,<br />
serrate, posterior margin with truncate median excision,<br />
with 3 RS.<br />
Antennule peduncle extending to posterior <strong>of</strong> pereonite<br />
. Antenna flagellum extending to middle <strong>of</strong><br />
pereonite 3.<br />
Frontal lamina flat, as wide as long, diamond shaped,<br />
anterior margin acute, forming median angle, posterior<br />
margin forming narrow stem.<br />
Pereopod 1 basis 2.6 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
0 RS (3 small submarginal simple setae), superior distal<br />
margin with RS; merus inferior margin with 3 RS, set<br />
as two groups (minute, set as 2 and ), superior distal<br />
angle with 0 RS (and simple and plumose setae);<br />
carpus 0.9 as long as merus, inferior margin with<br />
RS (minute); propodus 2.3 times as long as proximal<br />
width, inferior margin with 0 RS, propodal palm<br />
85<br />
simple, without blade or process (concave), dactylus<br />
smoothly curved, .4 as long as propodus. Pereopod<br />
2 ischium inferior margin with RS, superior distal<br />
margin with RS; merus inferior margin with 4 RS (set<br />
as 2+2), set as two groups, superior distal margin with<br />
0 acute RS; carpus longer than that <strong>of</strong> pereopod , with<br />
inferodistal lobe (lobe weak), inferodistal angle with<br />
2 RS. Pereopod 3 similar to pereopod 2. Pereopods 5–7<br />
inferior margins <strong>of</strong> ischium–carpus with long acute RS.<br />
Pereopod 6 similar to pereopod 7. Pereopod 7 basis 3.0 as<br />
long as greatest width, inferior margins with 2 palmate<br />
setae; ischium 0.5 as long as basis, inferior margin with<br />
4 RS (set loosely as 2, and ), superior distal angle with<br />
7 RS, inferior distal angle with 5 RS; merus .2 as long<br />
as ischium, 2.2 times as long as wide, inferior margin<br />
with 4 RS (set singly), superior distal angle with 5 RS,<br />
inferior distal angle with 8 RS; carpus . as long as<br />
ischium, 2.5 times as long as wide, inferior margin with<br />
3 RS (set as and 2), superior distal angle with 4 RS,<br />
inferior distal angle with 8 RS; propodus 0.9 as long as<br />
ischium, 3.8 times as long as wide, inferior margin with<br />
3 RS (set singly), superior distal angle with 3 slender<br />
setae, inferior distal angle with 4 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7, or<br />
short rectangular lobes; penial openings separated by<br />
4% <strong>of</strong> sternal width, penial process 0.9 times as long<br />
as basal width.<br />
Pleopod 1 exopod .6 times as long as wide, distally<br />
broadly rounded, lateral margin weakly convex,<br />
mesial margin strongly convex, with PMS on distal<br />
one-third; endopod 2 times as long as wide, distally<br />
rounded, lateral margin straight, with PMS on distal<br />
one-third, mesial margin with PMS on distal one-third;<br />
peduncle .9 times as wide as long, mesial margin<br />
with coupling hooks. Pleopod 2 appendix masculina<br />
with straight margins, 0.8 times as long as endopod.<br />
Exopods <strong>of</strong> pleopods –3 each with distolateral margin<br />
not digitate; pleopods 2–4 peduncle distolateral margin<br />
without prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about one-half as long as endopod. Endopod<br />
apically not bifid, lateral margin straight, without<br />
prominent excision, proximal lateral margin with 0 RS,<br />
distal lateral margin with 2 RS, mesial margin weakly<br />
convex, with 0 RS. Exopod 2.8 times as long as greatest<br />
width, apically not bifid; lateral margin weakly<br />
convex, with 3 RS; mesial margin sinuate, proximally<br />
concave, with 5 RS.<br />
size: Males 8–32 mm, female 9 mm; male holotype<br />
45.3 mm.<br />
Variation: <strong>The</strong> number <strong>of</strong> robust setae on the pleotelson<br />
and mesial margin <strong>of</strong> the uropodal endopod<br />
are relatively inconsistent, including between the two
Figure 44. Aegiochus beri (Bruce, 983). A, male 32 mm (NMV J8885). A, dorsal view; B, lateral view; C, head; D, frons;<br />
E, pleonites, lateral view; F, pleotelson posterior margin; G, sternite 7 showing penial position; H, penes.<br />
86
Figure 45. Aegiochus beri (Bruce, 1983). All figs NMNZ Cr.9260. A–C, pereopods 1, 2 and 7 respectively; D, pleopod 1;<br />
E, pleopod 2; F, uropod; G, uropod exopod, ventral view.<br />
87
sides <strong>of</strong> the one specimen. <strong>The</strong> pleotelson has – 6<br />
RS (with the largest male having the least setae). Uropod<br />
endopod mesial margin 7– 2 RS (mean 9), lateral<br />
margin always 2 RS; uropod exopod mesial margin 5<br />
or 6 RS, lateral margin 2 or 5 RS.<br />
<strong>The</strong> robust setae on the inferior margin <strong>of</strong> the merus<br />
are minute, and it was not possible to obtain accurate<br />
counts under the stereomicroscope.<br />
<strong>The</strong> holotype, the largest known specimen, was<br />
described as having the penial openings flush with<br />
sternite 7. <strong>The</strong>re are three males in the present material,<br />
the largest <strong>of</strong> these with the penes forming short<br />
rectangular lobes, as illustrated. <strong>The</strong> smaller males<br />
have the penial openings flush. Whether this is related<br />
to age or sexual maturity is unclear.<br />
remarks: Aegiochus beri is one <strong>of</strong> two species in the region<br />
that have the apex <strong>of</strong> the pleotelson with a distinct<br />
rectangular notch. Aegiochus beri is further characterised<br />
by the following combination <strong>of</strong> characters: large<br />
but separate eyes, elongate coxae on pereonites 6 and<br />
7, the diamond-shaped frontal lamina with a distinct<br />
posterior stem, the relatively long dactylus on pereopods<br />
–3 (pereopod dactylus is .4 times as long as<br />
the propodus) and the straight appendix masculina<br />
which has a narrowly rounded blunt apex.<br />
<strong>The</strong> most similar species is Aegiochus riwha sp.<br />
nov. (p. 4 ), which is easily distinguished by having<br />
a much shorter dactylus on pereopods –3 (pereopod<br />
dactylus is .0 times as long as the propodus v. .4<br />
times as long for A. beri), coxae without acute points,<br />
the shorter coxae on pereonites 5–7 and males with a<br />
weakly spatulate appendix masculina.<br />
prey: Not known.<br />
distribution: Previously recorded <strong>of</strong>f southeastern<br />
Australia. Present material from the Tasman Sea, east <strong>of</strong><br />
Lord Howe Island and from the Coral Sea <strong>of</strong>f Queensland.<br />
At depths between 748 and 406 metres.<br />
Aegiochus bertrandi sp. nov. (Figs 46–48)<br />
materiaL examined: All material from southwest <strong>of</strong> <strong>New</strong><br />
Caledonia, vicinity <strong>of</strong> Norfolk Ridge.<br />
Holotype: ♂ ( 6.0 mm), Norfolk Ridge, 9°04.0’S,<br />
63°27.50’E, 8 September 985, 260 m, MUSORSTOM<br />
IV, stn 0 84 (MNHN Is. 5903).<br />
Paratypes: 2 ♂ ( 2.3, 4.8 [dissected] mm), Norfolk<br />
Ridge, 24°42.26’S, 68°09.52’E, 27 October 986, 230 m,<br />
CHALCAL II, DW7 , coll. B. Richer de Forges (MNHN<br />
Is.5907). ♂ ( 5. mm), 20°4 .80’S, 67°00.20’E, 4 February<br />
989, 282 m, MUSORSTOM VI, DW399, coll. B.<br />
Richer de Forges (MNHN Is.5906). ♂ ( 3.4 mm), Banc<br />
Kaimon–Maru, Norfolk Ridge, 24°43’S, 68°09’E, 22<br />
June 200 , 227–232 m, NORFOLK , DW 676, coll.<br />
88<br />
Lazouet, Bouchet, Richer de Forges, N.O. Alis (MNHN<br />
Is.5902). ♀ (non-ovig. 2.8 mm), Banc Kaimon–Maru,<br />
Norfolk Ridge, 24°44’S, 68° 0’E, 22 June 200 , 228–240<br />
m, NORFOLK , CP 68 , coll. Lazouet, Boisselier,<br />
Richer de Forges, N.O. Alis (MNHN Is.5900). ♂ ( 8.0<br />
mm), Norfolk Ridge, 22°56.0’S, 67°20.0’E, 30 August<br />
985, 300 m, MUSORSTOM IV, 0227 (MNHN Is.<br />
5905). ♀ (ovig. 6.0 mm), Norfolk Ridge, 22°48.03’S,<br />
67°29.03’E, 2 May 993, 299–302 m, BATHUS 2,<br />
DW727, coll. B. Richer de Forges, N.O. Alis (MNHN<br />
Is.5899). 2 ♂ ( 4.4, 5.8 [telson damaged] mm), Norfolk<br />
Ridge, 23°4 .2’S, 68°00.5’E, 7 September 989, 280<br />
m, SMIB 5, DW76, coll. B. Richer de Forges (NIWA<br />
24007). ♀ (non-ovig. 5.2 mm), Norfolk Ridge, 23°38’S,<br />
67°42’E, 22 May 989, 448 m, SMIB 3, DW 3, coll. B.<br />
Richer de Forges (MNHN Is.5904). ♀ (ovig. 5.4 mm),<br />
Norfolk Ridge, 23°4 .5’S, 67°59.4’E, 23 May 989, 338<br />
m, SMIB 3, DW 8, coll. B. Richer de Forges, N. Vauban<br />
(MNHN Is.590 ).<br />
Other material (equivocal): ♀ (ovig. 5.6 mm),<br />
Norfolk Ridge, 22°20.0’S, 68°42.3’E, 3 September<br />
989, 255 m, SMIB 5, DW93, coll. B. Richer de Forges<br />
(MNHN Is.5908). 2, Lagon du nord, DW 074, 9°50.8’S,<br />
64°00.6’E, 28 m (MNHN Is.5909).<br />
desCription: Body dorsal surfaces smooth or polished<br />
in appearance, widest at pereonite 5, lateral margins<br />
weakly ovate. Rostral point ventrally directed. Eyes<br />
large, medially united, anterior clear field 21% length<br />
<strong>of</strong> head, posterior clear field 79% length <strong>of</strong> head; each<br />
eye made up <strong>of</strong> ~ 2 transverse rows <strong>of</strong> ommatidia,<br />
each row with ~7 ommatidia; eye colour pale brown.<br />
Pereonite 1 and coxae 2–3 each with posteroventral angle<br />
right-angled. Coxae 5–7 with entire oblique carina;<br />
posterior margins concave, posterolateral angle acute<br />
(less than 45°). Pleon with pleonite visible in dorsal<br />
view; pleonite 4 with posterolateral margins extending<br />
to but not beyond posterior margin <strong>of</strong> pleonite 5;<br />
pleonite 5 with posterolateral angles free, not overlapped<br />
by lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.9<br />
times as long as anterior width, dorsal surface without<br />
longitudinal carina; lateral margins weakly convex,<br />
serrate, posterior margin converging to caudomedial<br />
point, with 0 RS.<br />
Antennule peduncle articles 3 and 4 .03 times as<br />
long as combined lengths <strong>of</strong> articles and 2, article 3<br />
2.7 times as long as wide; flagellum with 13 articles,<br />
extending to posterior <strong>of</strong> pereonite . Antenna peduncle<br />
article 4 .5 times as long as wide, 0.7 times as long<br />
as combined lengths <strong>of</strong> articles –3, inferior margin 0<br />
plumose setae, and 0 short simple setae (anterodistal<br />
angle 5 simple + plumose); article 5 .5 times as long<br />
as article 4, 2.3 times as long as wide, inferior margin<br />
with palmate seta (at distal angle), anterodistal angle<br />
with cluster <strong>of</strong> 3 short simple setae; flagellum with 19<br />
articles, extending to pereonite 4.
Figure 46. Aegiochus bertrandi sp. nov. A–H, holotype, remainder male paratype ( 4.8 mm, MNHN Is. 5907). A, dorsal view;<br />
B, lateral view; C, head; D, frons, anterior view; E, frons, ventral view; F, penial processes; G, antennule; H, antenna peduncle;<br />
I, pleonites, lateral view; J, sternite 7; K, pleotelson posterior margin.<br />
89
Figure 47. Aegiochus bertrandi sp. nov. Male paratype ( 4.8 mm, MNHN Is. 5907). A, maxilliped; B, maxilliped articles 3–5;<br />
C, maxilla apex; D, maxillule apex; E, pereopod ; F, pereopod 2; G, pereopod 7.<br />
90
Figure 48. Aegiochus bertrandi sp. nov. Male paratype ( 4.8 mm, MNHN Is. 5907). A–D, pleopods –3, 5 respectively; E, uropod<br />
exopod, ventral view; F, uropod, dorsal view.<br />
Frontal lamina flat, as wide as long, diamond shaped<br />
or posteriorly rounded, anterior margin acute, without<br />
small median point, posterior margin not abutting<br />
clypeus or forming narrow stem.<br />
Mandible molar process present, small distinct flat<br />
lobe. Maxillule with 4 terminal RS ( large, 3 slender;<br />
small triangular spines proximal to RS). Maxilla mesial<br />
9<br />
lobe with 3 RS ( hooked; 2 straight, serrate); lateral lobe<br />
with 3 RS. Maxilliped endite with 0 apical setae; palp<br />
article 2 with 2 RS (small); article 3 with 2 recurved RS<br />
(and 4 straight RS); article 4 with 4 hooked RS; article<br />
5 with 3 RS (all straight; serrate, 2 simple).<br />
Pereopod 1 basis 2.5 times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with
0 RS, superior distal margin with 2 RS; merus inferior<br />
margin with 2 RS, set as two groups, superior distal<br />
angle with 0 RS (2 simple setae); carpus . as long as<br />
merus, inferior margin with RS; propodus .8 times<br />
as long as proximal width, inferior margin with RS<br />
(distal), propodal palm simple, without blade or process,<br />
dactylus smoothly curved, . as long as propodus.<br />
Pereopod 2 ischium inferior margin with 0 RS, superior<br />
distal margin with 2 RS; merus inferior margin with<br />
4 RS, set as two groups, superior distal margin with 0<br />
acute RS (3 simple setae); inferodistal angle with RS.<br />
Pereopod 3 similar to pereopod 2. Pereopods 5–7 inferior<br />
margins <strong>of</strong> ischium–carpus with short RS. Pereopod 6<br />
similar to pereopod 7. Pereopod 7 basis 2.9 times as<br />
long as greatest width, inferior margins with 9 palmate<br />
setae; ischium 0.5 as long as basis, inferior margin<br />
with 3 RS (set as and 2), superior distal angle with 4<br />
RS, inferior distal angle with 2 RS; merus .2 as long<br />
as ischium, 2.7 times as long as wide, inferior margin<br />
with 4 RS (set as 2 and 2), superior distal angle with 9<br />
RS, inferior distal angle with 5 RS; carpus .0 as long<br />
as ischium, 3. times as long as wide, inferior margin<br />
with 3 RS (set as and 2), superior distal angle with<br />
2 RS, inferior distal angle with 8 RS; propodus .0<br />
as long as ischium, 5.7 times as long as wide, inferior<br />
margin with 6 RS (set as , , 2 and 2), superior distal<br />
angle with 2 slender setae (and RS and palmate<br />
seta), inferior distal angle with 3 RS.<br />
Penes low tubercles; penial openings separated by<br />
2% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod .9 times as long as wide, distally<br />
broadly rounded, lateral margin straight, mesial margin<br />
weakly convex (proximally angled), with PMS on<br />
distal two-thirds; endopod .8 times as long as wide,<br />
distally rounded, lateral margin straight, with PMS on<br />
distal margin only, mesial margin with PMS on distal<br />
half; peduncle .8 times as wide as long, mesial margin<br />
with 6 coupling hooks. Pleopod 2 appendix masculina<br />
basally swollen, 0.9 times as long as endopod, distally<br />
acute. Exopods <strong>of</strong> pleopods –3 each with distolateral<br />
margin digitate (prominent, acute); endopods <strong>of</strong> pleopods<br />
3–5 each with distolateral point; pleopods 2–4<br />
peduncle distolateral margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about two-thirds as long as endopod.<br />
Endopod apically not bifid, lateral margin straight, without<br />
prominent excision, proximal lateral margin with<br />
RS, distal lateral margin with 2 RS, mesial margin<br />
weakly convex, with 5 RS. Exopod not extending to end<br />
<strong>of</strong> endopod, 3.0 as long as greatest width, apically not<br />
bifid; lateral margin weakly convex, with 10 RS; mesial<br />
margin straight, distally convex, with 4 RS.<br />
femaLe: As for the male, but lacking sexual characters.<br />
92<br />
size: Males 2.3– 8.0 mm (mean = 5.0, n = 8), ovi-<br />
gerous females 5.4– 6.0 mm, non-ovigerous 2.8 and<br />
5.2 mm.<br />
Variation: Pleotelson (n = 2) always without RS. Uropod<br />
(n = 24, all margins) exopod mesial margin usually<br />
with 3 (8%) or 4 (92%) RS, lateral margin with 9 (each<br />
42%) or 0 (each 50%) RS ( occurring on one specimen);<br />
uropod endopod mesial margin with 4 ( 7%) or<br />
5 (83%) RS (one specimen with 3), lateral margin with<br />
+ ( 00%) RS.<br />
remarks: Aegiochus bertrandi sp. nov. is best identified<br />
by the narrowly united eyes, weak rostral point in dorsal<br />
view, anteriorly acute frontal lamina, small robust<br />
setae on pereopod , the inferior margin <strong>of</strong> the carpus <strong>of</strong><br />
pereopods 2 and 3 distinctly lobed with one prominent<br />
robust seta, pleopodal exopods with digitate margins,<br />
relatively wide and serrated pleotelson that lacks robust<br />
setae, pleotelson posterior margins converging to<br />
a distinct sub-acute apical point, and the characteristic<br />
shape and setation <strong>of</strong> the uropodal rami.<br />
This species is very similar to the sympatric Aegiochus<br />
coroo Bruce, 983, but differs consistently in a<br />
number <strong>of</strong> characters, including larger robust setae on<br />
pereopods –3, a distinct lobe on the inferior margin <strong>of</strong><br />
the carpus <strong>of</strong> pereopods 2 and 3, narrower eye join (as<br />
few as two ommatidia), narrower pleotelson posterior<br />
margin, narrower (or more acute) uropod endopod<br />
which extends beyond the pleotelson, the uropodal<br />
endopod mesial margin has five robust setae (three<br />
or four in A. coroo) and the most distal notch on the<br />
uropodal exopod lateral margin always has a robust<br />
seta (slender setae only in A. coroo); A. bertrandi is also<br />
larger (male average length 5.0 mm) than A. coroo<br />
(male average length .4 mm). <strong>The</strong> counts for uropodal<br />
robust setae do not differ much between the two<br />
species but the differences are consistent.<br />
distribution: Not recorded from <strong>New</strong> <strong>Zealand</strong> nearshore<br />
waters to date, but is present within the northern<br />
<strong>New</strong> <strong>Zealand</strong> chart area (CANZ 997). <strong>The</strong> most southerly<br />
record is at 24°44’S on the Norfolk Ridge, most<br />
northerly at 9°S in the vicinity <strong>of</strong> <strong>New</strong> Caledonia; it<br />
is quite probable that the species is more widespread<br />
in the region. At depths <strong>of</strong> 230 to 448 metres.<br />
etymoLogy: Named for Dr Bertrand Richer de Forges,<br />
recognising his superb efforts in developing collections<br />
<strong>of</strong> marine invertebrates from the southwestern<br />
Pacific and his contribution to taxonomy <strong>of</strong> the decapod<br />
crustaceans.
Aegiochus coroo (Bruce, 983), comb. nov.<br />
(Figs 49–5 )<br />
Aega (Rhamphion) coroo Bruce, 1983: 770, figs 9, 10.<br />
Aega coroo. Springthorpe & Lowry, 994: 43. – Bruce, Lew<br />
Ton & Poore, 2003: 6 .<br />
materiaL examined: <strong>New</strong> <strong>Zealand</strong>. ♂ ( 2.0 mm), ♀<br />
(ovig. 2.5 mm), northwest <strong>of</strong> Taranaki, 38°37.99’S,<br />
72°40.99’E, 28 March 968, E908, 256 m (NIWA 24006).<br />
<strong>New</strong> caledonia and Norfolk ridge. 4 ♂ (8.5, 0.3, .7<br />
[drawn dissected], 3.0 mm), 4 ♀ (ovig. 2.5, non-ovig.<br />
4.2, 4. , 0.4 mm), Banc Eponge, Norfolk Ridge,<br />
24°56’S, 68°22’E, 23 June 200 , 533–545 m, NORFOLK<br />
, DW 688, coll. Lazouet, Boisselier, Richer de Forges,<br />
N.O. Alis (MNHN Is.5925). ♀ (non-ovig. 4.5 mm),<br />
Coral Sea, 20°06.084–0.6. 85’S, 60°23.544–22.835’E, 20<br />
October 2005, 490–550 m, DW26 9, (MNHN Is.59 9).<br />
♂ (9.0 mm), Norfolk Ridge, 24°06.960–0.6. 2’S,<br />
59°4 .270–4 .500’E, 8 October 2005, 350–400 m,<br />
DW2520, coll. (MNHN Is.59 8). ♂ ( 2.8 mm), Norfolk<br />
Ridge, 22°53’S, 67° 2’E, 28 June 200 , 403–429 m,<br />
NORFOLK , DW 734, coll. Lazouet, Bouchet, Richer<br />
de Forges, N.O. Alis (MNHN Is.5927). ♂ ( .5 mm), ♀<br />
(ovig. 3.5 mm), Banc Eponge, Norfolk Ridge, 24°55’S,<br />
68°22’E, 23 June 200 , 508–54 m, NORFOLK ,<br />
DW 684, coll. Lazouet, Bouchet, Richer de Forges, N.O.<br />
Alis (MNHN Is.5926 [small tube with female]). ♂ ( .0<br />
mm), Banc Branchiopode, Norfolk Ridge, 234°28’S,<br />
67°5 ’E, 2 October 2003, 37 m, NORFOLK 2, stn 2024<br />
(MNHN Is. 59 5). ♂ ( .5 mm), 24°56’S, 68°2 ’E, 2<br />
May 987, 505 m, SMIB 3, DW6, coll. Richer de Forges,<br />
N.O. Vauban (MNHN Is. 5944). solomon Islands. ♂<br />
(~ 3 mm), 09°46.4’S, 60°52.3’E, 7 October 200 , 254–<br />
28 m, SALOMON , DW 856, coll. N.O. Alis (MNHN<br />
Is.5920). Indonesia. ♂ ( 2.5 mm), (non-ovig. 2.2 mm),<br />
Tanimbar Islands, 07°59’S, 33°02’E, 20 October 99 ,<br />
84– 86 m, KARUBAR, DW50, coll. N.O. Baruna Jaya<br />
(MNHN Is.5922). ♂ ( .5 mm), Kai Islands, 057° 8’S,<br />
32°38’E, 24 October 99 , 246 m, KARUBAR, DW 4,<br />
coll. N.O. Baruna Jaya (MNHN Is. 592 ).<br />
Additional material. Southwest <strong>of</strong> <strong>New</strong> Caledonia,<br />
vicinity <strong>of</strong> Norfolk ridge. 5, 24°55’S, 68°22’E, 505–<br />
5 5 m, BIOCAL, DW66 (MNHN Is.5937). , 22° 5’S,<br />
67° 5’E, 440 m, BIOCAL, DW77 (MNHN Is.5936).<br />
3, 20°35’S, 66°54’E, 460 m, BIOCAL, DW83 (MNHN<br />
Is.5938). , 22°49.32’S, 66°44.68’E, 300–370 m, BATHUS<br />
2, DW73 (MNHN Is.5952). , 24°54’S, 68°2 ’E, 540–<br />
570 m, BERyx , CP08 (MNHN Is.595 ). , 24°52’S,<br />
68°22’E, 635–680 m, BERyx , DW09 (MNHN<br />
Is.5949). 4, 24°53’S, 68°2 ’E, 565–600 m, BERyx ,<br />
DW 0 (MNHN Is.5950). , 24°44.6’S, 68°09.3’E, 230<br />
m, CHALCAL II, CP20 (MNHN Is.59 7). , 24°54.00’S,<br />
68°2 .0 ’E, 500 m, CHALCAL II, CP2 (MNHN<br />
Is.59 6). 20, 24°54.5’S, 68°22.3’E, 527 m. CHALCAL<br />
II, DW72 (NIWA 24005). , 9°04.0’S, 63°27.5’E,<br />
93<br />
260 m, MUSORSTOM IV, 0 84 (MNHN Is.5932). ,<br />
22°5 .3’S, 67° 2.0’E, 405–430 m, MUSORSTOM IV,<br />
02 3 (MNHN Is.5933). , 22°52.5’S, 67° .8’E, 390–420<br />
m, MUSORSTOM IV, 0230 (MNHN Is.5934). 5, 24°56’S,<br />
68°22’E, 520 m, SMIB 3, DW (MNHN Is.5943). ,<br />
24°53’S, 68°22’E, 530–537 m, SMIB 3, DW2 (MNHN<br />
Is.5945). 2, 24°55’S, 68°22’E, 5 3 m, SMIB 3, DW3<br />
(MNHN Is.5946). , 24°55’S, 68°22’E, 502–5 2 m, SMIB<br />
3, DW5 (MNHN Is.594 ). , 24°55.2’S, 68°2 .7’E, 5 –<br />
522 m, SMIB 8, DW 46 (MNHN Is.5947). 2, 24°55. ’S,<br />
68°2 .6’E, 5 0 m, SMIB 8, DW 48 (MNHN Is.5948).<br />
further lots from the region, not listed individually,<br />
MNHN, not registered). Eastern Australia. , northern<br />
Queensland, 7°53’S, 46°53’E, 96 m (QM W 8823). ,<br />
southern Queensland, 28° 7.47’S, 58°37.89’E, 4 9 m<br />
(AM P74739). 5, East <strong>of</strong> Nobby’s Head, NSW, 32°53’S,<br />
52°35’E, 75 m, (AM P37503). , east <strong>of</strong> Long Reef,<br />
NSW, K85- 2-08, 9 December 985, 74 m, (AM<br />
P43977). , east <strong>of</strong> Long Reef Point, NSW, 33°46’S,<br />
5 °43’E, 75 m, (AM P375 0). 7, East <strong>of</strong> Long Reef<br />
Point, NSW, 33°43–44’S, 5 °46’E, 74 m, (AM P37506).<br />
, southeast <strong>of</strong> Moruya Point, NSW, 35°58’–36°03’S,<br />
50°30–27’E, 384 m, (AM P37522). 3, <strong>of</strong>f Merimbula,<br />
NSW, 36°52.5’S, 50° 8. ’E, 52 m (AM P45309). ,<br />
east <strong>of</strong> Bermagui coast, NSW, 37°26.5’S, 50° 7.0’E,<br />
coll. CSIRO (AM P375 3). 4, ‘7-5 50 (AM P375 9). ,<br />
<strong>of</strong>f Bermagui, SS 05/94/ 56, 5 September 994, 46<br />
m (AM P43969). 4, east <strong>of</strong> Bermagui, NSW, 37°25.2’S,<br />
50° 8.5’E, 220 m (AM P453 0). , transect east <strong>of</strong> Bermagui,<br />
NSW, 36°22.6’S, 50° 4.9’E, 8 September 994,<br />
277 m (AM P44 50).<br />
type LoCaLity: Off Sydney, <strong>New</strong> South Wales, 33°59’S,<br />
5 °35’E.<br />
desCription: Body 2.8 times as long as greatest width,<br />
dorsal surfaces polished in appearance, widest at<br />
pereonite 5, lateral margins subparallel. Rostral point<br />
ventrally directed. Eyes large, medially united, anterior<br />
clear field 23% length <strong>of</strong> head, posterior clear field 45%<br />
length <strong>of</strong> head; each eye made up <strong>of</strong> ~ 2 transverse<br />
rows <strong>of</strong> ommatidia, each row with ~8 ommatidia; eye<br />
colour dark brown, or pale brown. Pereonite 1 and coxae<br />
2–3 each with posteroventral angle right-angled. Coxae<br />
5–7 with entire oblique carina; posterior margins concave,<br />
posterolateral angle acute (less than 45°). Pleon<br />
with pleonite visible in dorsal view; pleonite 4 with<br />
posterolateral margins extending to but not beyond<br />
posterior margin <strong>of</strong> pleonite 5; pleonite 5 with posterolateral<br />
angles free, not overlapped by lateral margins <strong>of</strong><br />
pleonite 4. Pleotelson 0.8 times as long as anterior width,<br />
dorsal surface without longitudinal carina; lateral margins<br />
convex, serrate, posterior margin converging to<br />
caudomedial point, with 0 RS.<br />
Antennule peduncle extending to posterior <strong>of</strong> pereonite<br />
. Antenna flagellum extending to pereonite 4.
Figure 49. Aegiochus coroo (Bruce, 983). Male ( .7 mm, MNHN Is. 5925). A, dorsal view; B, lateral view; C, head; D, frontal<br />
lamina, anterior view; E, frons, ventral view; F, penial processes; G, sternite 7; H, pleonites, lateral view; I, pleotelson posterior<br />
margin.<br />
Frontal lamina flat, as wide as long, diamond shaped<br />
or posteriorly rounded (depending on perspective),<br />
anterior margin acute, without small median point,<br />
posterior margin forming narrow stem.<br />
Mandible molar process present, minute. Maxillule<br />
with 3 terminal RS ( large, 2 slender; small triangular<br />
spines proximal to RS). Maxilla mesial lobe with 3 RS (2<br />
serrate, simple); lateral lobe with 3 RS. Maxilliped endite<br />
with apical seta; palp article 2 with 2 RS (straight);<br />
article 3 with recurved RS (weakly recurved and 2<br />
94<br />
straight RS); article 4 with 4 hooked RS; article 5 with<br />
2 RS ( large, serrate; slender, simple).<br />
Pereopod 1 basis 3.2 times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 0 RS ( slender seta);<br />
merus inferior margin with 0 RS, superior distal angle<br />
with 0 RS ( slender seta); carpus .0 as long as merus,<br />
inferior margin with 0 RS (2 small nodules); propodus<br />
.8 times as long as proximal width, inferior margin<br />
with 0 RS, propodal palm simple, without blade or
Figure 50. Aegiochus coroo (Bruce, 983). Male ( .7 mm, MNHN Is. 5925). A, maxilliped; B, maxilla apex; C, maxilliped articles<br />
3–5; D, maxillule apex; E, pereopod ; F, pereopod 2; G, pereopod 2 merus; H, pereopod 3, distal articles; I, pereopod 7.<br />
process, dactylus smoothly curved, .3 as long as propodus.<br />
Pereopod 2 ischium inferior margin with 0 RS,<br />
superior distal margin with 0 RS ( stiff seta); merus<br />
inferior margin with 3 RS, set as two groups ( +2),<br />
95<br />
superior distal margin with 0 acute RS (2 stiff setae);<br />
carpus similar in size to that <strong>of</strong> pereopod , inferodistal<br />
angle with RS (and 2 distinct nodules). Pereopod 3<br />
similar to pereopod 2. Pereopods 5–7 inferior margins
Figure 51. Aegiochus coroo (Bruce, 983). Male ( .7 mm, MNHN Is.5925). A, pleopods ; B pleopod 2; C, uropod, dorsal view;<br />
D, uropod exopod, ventral view.<br />
<strong>of</strong> ischium–carpus with short RS. Pereopod 6 similar<br />
to pereopod 7. Pereopod 7 basis 2.5 times as long as<br />
greatest width, inferior margins with 8 palmate setae;<br />
ischium 0.4 as long as basis, inferior margin with 3 RS<br />
(set singly), superior distal angle with 3 RS, inferior<br />
distal angle with 2 RS; merus .6 as long as ischium, 2.9<br />
times as long as wide, inferior margin with 3 RS (set as<br />
and 2), superior distal angle with 8 RS, inferior distal<br />
angle with 5 RS; carpus .6 as long as ischium, 3.2 times<br />
as long as wide, inferior margin with 2 RS (set singly),<br />
superior distal angle with 8 RS, inferior distal angle<br />
with 5 RS; propodus .3 as long as ischium, 4.2 times<br />
as long as wide, inferior margin with 2 RS (set singly),<br />
superior distal angle with slender seta, inferior distal<br />
angle with 3 RS.<br />
Penes low tubercles; penial openings separated by<br />
2% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod .9 times as long as wide, distally<br />
broadly rounded, lateral margin straight, mesial<br />
margin strongly convex, with PMS from distal half;<br />
endopod .6 times as long as wide, distally rounded,<br />
96<br />
lateral margin straight, with PMS on distal margin only,<br />
mesial margin with PMS on distal half; peduncle .7<br />
times as wide as long, mesial margin with 5 coupling<br />
hooks. Pleopod 2 appendix masculina with straight<br />
margins, 0.8 times as long as endopod, distally narrowly<br />
rounded. Exopods <strong>of</strong> pleopods –3 each with<br />
distolateral margin digitate; endopods <strong>of</strong> pleopods 3–5<br />
each with distolateral point; pleopods 2–5 peduncle<br />
distolateral margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about two-thirds as long as endopod.<br />
Uropod rami extending beyond pleotelson (just a little).<br />
Endopod apically sub-bifid, mesial process prominent,<br />
lateral margin straight, without prominent excision,<br />
proximal lateral margin with RS, distal lateral margin<br />
with 2 RS, mesial margin weakly convex, with 4<br />
RS. Exopod not extending to end <strong>of</strong> endopod, 2.4 times<br />
as long as greatest width, apically sub-bifid, lateral<br />
process prominent; lateral margin weakly convex,<br />
with 9 RS; mesial margin straight, distally convex,<br />
with 3 RS.
size: Males 8.5– 3.0 mm (mean = .4, n = 3), ovigerous<br />
females 2.5– 3.50 mm, non-ovigerous 2.2– 4.5<br />
mm (mean = 3.8, n = 5).<br />
Variation: Pleotelson (n = 6) always without RS. Uropod<br />
(n = 32, all margins) exopod mesial margin usually<br />
with 2 ( 6%) or 3 (8 %) RS (4 once), lateral margin with<br />
8 or 9 (each 47%) RS (7 and 0 each occurring once);<br />
uropod endopod mesial margin with 4 (94%) RS (one<br />
specimen with 3), lateral margin with + (3 %) or<br />
+2 (69%) RS.<br />
remarks: Aegiochus coroo is a wide-ranging species,<br />
apparently common, at first glance similar to several<br />
other small species <strong>of</strong> the genus. <strong>The</strong> digitate pleopod<br />
exopods immediately separate it from most other<br />
Aegiochus species. Within that group <strong>of</strong> small-sized<br />
species that have digitate pleopods A. coroo is readily<br />
identified by the medially united eyes, lack <strong>of</strong> a rostral<br />
point in dorsal view, anteriorly acute frontal lamina,<br />
pereopod devoid <strong>of</strong> robust setae, small robust setae<br />
on pereopods 2 and 3, relatively wide serrated pleotelson<br />
posterior margin which lacks robust setae, and<br />
the characteristic shape and setation <strong>of</strong> the uropodal<br />
rami. <strong>The</strong> pleotelson is evenly serrated and typically<br />
there are, on either side, two sub-lateral notches that<br />
are a little wider than the remainder, each bearing<br />
three setae within the notch. Care is need in assessing<br />
the shape <strong>of</strong> the frontal lamina, which can appear<br />
posteriorly rounded or diamond-shaped depending<br />
on perspective.<br />
<strong>The</strong>re are a few points <strong>of</strong> difference to the description<br />
given by Bruce ( 983). <strong>The</strong> present description corrects<br />
these and provides more detail for the pereopods,<br />
some mouthparts and the uropods. Bruce ( 983) stated<br />
that the vasa deferentia opened flush with the ventral<br />
surface <strong>of</strong> sternite 7; on the larger males examined here<br />
it is evident that a pair <strong>of</strong> low papillae is present. <strong>The</strong><br />
figure <strong>of</strong> maxilliped palp (Bruce 1983, fig. 9k) omits article<br />
5, and that is here fully illustrated. <strong>The</strong>re are some<br />
minor differences in details <strong>of</strong> setation, attributable<br />
to perspective and differences in microscope resolution.<br />
In all significant characters, the present material<br />
agrees well with the description and figures <strong>of</strong> Bruce<br />
( 983) and the species presents a consistent appearance<br />
throughout its geographical range.<br />
<strong>The</strong> sympatric species, Aegiochus bertrandi sp. nov.,<br />
is very similar, but has noticeably narrower join between<br />
the eyes, more slender uropodal endopod, more<br />
strongly angular pleotelson, larger robust setae on the<br />
inferior margins <strong>of</strong> pereopod –3, the inferior margin<br />
<strong>of</strong> the carpus <strong>of</strong> pereopods 2 and 3 is lobate, and the<br />
mesial margin <strong>of</strong> the uropodal endopod consistently<br />
has five robust setae (as opposed to consistently four<br />
97<br />
in A. coroo). <strong>The</strong> two species are otherwise nearly<br />
identical, and care is needed in identification. Other<br />
similar species in the <strong>New</strong> <strong>Zealand</strong> region are A. kakai<br />
sp. nov. and A. kanohi sp. nov., both <strong>of</strong> similar size to<br />
A. coroo, both with large, medially united eyes. Both<br />
these species have a blade-like frontal lamina, robust<br />
setae on the pleotelson and pleopods –3 exopods are<br />
not digitate. Those other <strong>New</strong> <strong>Zealand</strong> species which<br />
do have digitate pleopods all have separate eyes.<br />
prey: Not known.<br />
distribution: Northern <strong>New</strong> <strong>Zealand</strong>, <strong>New</strong> Caledonia,<br />
Solomon Islands, eastern Australia, also Indian Ocean,<br />
Arafura Sea at Kei and Tanimbar Islands; potentially<br />
widespread in the western Pacific; recorded depths <strong>of</strong><br />
230 to 600 metres in the Pacific; most shallow at 184<br />
metres in Indonesia.<br />
Aegiochus gordoni sp. nov. (Figs 52–55)<br />
materiaL: Holotype: ♂ (7.6 mm), Chatham Rise,<br />
42°45.89’S, 79°59. 6’W, 9 April 200 , 800–757 m, coll.<br />
RV Tangaroa (NIWA 23872).<br />
Paratypes: 4 ♂ (7.5, 8.0, 8. dissected, 2.5 mm), 35<br />
♀ ( 3 ovig .0– 3.3 mm, 22 non-ovig. 8.0– 5.0 mm),<br />
same data as holotype (NIWA 23873). ♂ (7.5 mm), 7<br />
♀ (ovig. 0.5, .5, 2.2, 2.5 broken, non-ovig .0,<br />
2.5 mm), manca (6.5 mm), Chatham Rise, 42°45.76’S,<br />
79°59.29’W, 6 April 200 , 064–750 m, coll. RV Tangaroa<br />
(NIWA 23874).<br />
Unmeasured. , Rumble V Sea Mount, 36°8.48–79’S,<br />
78° .70–53’E, 24 May 200 , 755–360 m, coll. RV<br />
Tangaroa (NIWA 23878); 2, Rumble V sea mount, 24<br />
May 200 , 36°8.07–40’S, 78° 2.07– .8 ’E, 40–698<br />
m, coll. RV Tangaroa (NIWA 23879). chatham rise: ,<br />
42°45.93’S, 79°59.34’W, 5 April 200 , 875–757 m, coll.<br />
R.V Tangaroa (NIWA 23875); , 42°45.68’S, 79°59.33’W,<br />
2 April 200 , 920–77 m, coll. RV Tangaroa (NIWA<br />
23876); 4, 42°42.84’S, 79°57.5 ’W, 8 April 200 , 980–<br />
893 m, coll. RV Tangaroa (NIWA 23877); , J 2/7/85,<br />
rock cavities (NMNZ Cr. 2020); , 42.7 58–7 52°S,<br />
80.0378–0352°E, 4 June 2006, 985– 050 m, (NIWA<br />
25667).<br />
desCription: Body 2.3 times as long as greatest width,<br />
dorsal surfaces polished in appearance, widest at pereonite<br />
5, lateral margins ovate. Rostral point folded<br />
ventrally and posteriorly. Eyes large, not medially<br />
united, separated by about 7% width <strong>of</strong> head; each<br />
eye made up <strong>of</strong> ~ 2 transverse rows <strong>of</strong> ommatidia,<br />
each row with ~8 ommatidia; eye colour dark brown.<br />
Pereonite 1 and coxae 2–3 each with posteroventral angle<br />
right-angled. Coxae 5–7 with entire oblique carina;
Figure 52. Aegiochus gordoni sp. nov. A–F, holotype, remainder male paratype (8. mm, NIWA 23873). A, dorsal view; B, lateral<br />
view; C, head; D, frons, ventral view; E, frontal lamina and rostrum, anterior view; pleotelson, posterior margin; F, sternite 7<br />
showing penial openings; G, pleotelson margins; H, antennule (distal flagellar articles missing); I, antenna peduncle.<br />
98
Figure 53. Aegiochus gordoni sp. nov. Male paratype (8. mm, NIWA 23873). A, mandible (broken); B, mandible palp article<br />
3; C, maxillule; D, maxillule apex; E, maxilla ; F, maxilla apex; G, maxilliped; H, maxilliped articles 2–5.<br />
posterior margins straight, posterolateral angle blunt<br />
(coxae 5 and 6, more than 45°) and acute (coxa 7, acute<br />
less than 45°). Pleon with pleonite largely concealed<br />
by pereonite 7; pleonite 4 with posterolateral margins<br />
not extending to posterior margin <strong>of</strong> pleonite 5; pleonite<br />
5 with posterolateral angles free, not overlapped<br />
by lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.7 times as<br />
long as anterior width, dorsal surface without longitudinal<br />
carina; lateral margins convex, serrate (finely),<br />
posterior margin converging to caudomedial point,<br />
with 0 RS.<br />
Antennule peduncle articles 3 and 4 .0 times as<br />
long as combined lengths <strong>of</strong> articles and 2, article 3<br />
99<br />
3.0 times as long as wide; flagellum with 10 articles,<br />
extending to pereonite 2. Antenna peduncle article 4 .9<br />
times as long as wide, 0.7 times as long as combined<br />
lengths <strong>of</strong> articles –3, inferior margin 2 plumose setae,<br />
and 4 short simple setae; article 5 .4 times as long as<br />
article 4, 2.8 times as long as wide, inferior margin<br />
with 3 palmate setae, anterodistal angle with cluster <strong>of</strong><br />
2 short simple setae (and 2 pappose setae); flagellum<br />
with articles, extending to posterior <strong>of</strong> pereonite 3.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, triangular, anterior<br />
margin acute, forming median angle, posterior<br />
margin not abutting clypeus.
Figure 54. Aegiochus gordoni sp. nov. Male paratype (8. mm, NIWA 23873). A–E, pereopods –3, 6 and 7 respectively.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 3 distolateral setae (large serrate<br />
and 2 simple), palp article 3 with 6 setae (all serrate).<br />
Maxillule with 5 terminal RS (4 slender, large).<br />
Maxilla mesial lobe with 2 RS ( strongly serrate,<br />
hooked); lateral lobe with 3 RS. Maxilliped endite with<br />
0 apical setae; palp article 2 with 2 RS; article 3 with 3<br />
recurved RS (slender); article 4 with 4 hooked RS (and<br />
small slender; unevenly spaced with proximal 3 distal);<br />
article 5 with 4 RS (straight, <strong>of</strong> which serrate).<br />
Pereopod 1 basis 2.5 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin<br />
with 0 RS, superior distal margin with RS; merus<br />
00<br />
inferior margin with RS (proximal, and long distal<br />
seta), superior distal angle with 0 RS (2 slender setae);<br />
carpus 0.9 as long as merus, inferior margin with RS;<br />
propodus 2. times as long as proximal width, inferior<br />
margin with 2 RS (small, close-set), propodal palm<br />
simple, without blade or process, dactylus smoothly<br />
curved, .4 as long as propodus. Pereopod 2 ischium<br />
inferior margin with 0 RS, superior distal margin with<br />
RS; merus inferior margin with 4 RS (distal RS large,<br />
remainder minute), set as two groups, superior distal<br />
margin with 2 acute RS; carpus longer than that <strong>of</strong> pereopod<br />
, with inferodistal lobe, inferodistal angle with<br />
RS. Pereopod 3 similar to pereopod 2 (RS on merus
Figure 55. Aegiochus gordoni sp. nov. Male paratype (8. mm, NIWA 23873). A–E, pleopods –5 respectively; F, uropod; G<br />
uropod endopod apex; H, uropod exopod apex; I, proximal robust seta, uropod endopod mesial margin.<br />
0
and carpus larger, 5 RS on inferior margin <strong>of</strong> carpus).<br />
Pereopods 5–7 inferior margins <strong>of</strong> ischium–carpus with<br />
short RS. Pereopod 6 similar to pereopod 7 (shorter,<br />
with more and longer RS; carpus inferior margin with<br />
+ 2 RS). Pereopod 7 basis 2.8 times as long as greatest<br />
width, inferior margins with 9 palmate setae; ischium<br />
0.4 as long as basis, inferior margin with 5 RS (set as ,<br />
2 and 2), superior distal angle with 2 RS, inferior distal<br />
angle with 2 RS (small); merus .3 as long as ischium,<br />
2.5 times as long as wide, inferior margin with 2 RS<br />
(set as and ), superior distal angle with 3 RS, inferior<br />
distal angle with 2 RS; carpus .5 as long as ischium,<br />
3.3 times as long as wide, inferior margin with 2 RS<br />
(single cluster), superior distal angle with 9 RS (many<br />
biserrate), inferior distal angle with 6 RS; propodus .3<br />
as long as ischium, 4.0 times as long as wide, inferior<br />
margin with 2 RS (single cluster), superior distal angle<br />
with 4 slender setae ( RS, plumose and 2 simple),<br />
inferior distal angle with 3 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by 8% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod .6 times as long as wide, distally<br />
narrowly rounded, mesial margin weakly oblique,<br />
lateral margin weakly convex, mesial margin strongly<br />
convex, with PMS on distal half, with ~36 PMS; endopod<br />
.9 times as long as wide, distally rounded, lateral<br />
margin convex, with PMS on distal half, mesial<br />
margin with PMS on distal one-third, endopod with<br />
~2 PMS; peduncle .9 times as wide as long, mesial<br />
margin with 7 coupling hooks. Pleopod 2 exopod with<br />
~43 PMS, endopod with ~27 PMS; appendix masculina<br />
with straight margins, .2 times as long as endopod<br />
(extending beyond distal margin <strong>of</strong> ramus), distally<br />
narrowly rounded. Pleopod 3 exopod with ~48 PMS,<br />
endopod with ~ 3 PMS. Pleopod 4 exopod with ~42<br />
PMS, endopod with ~8 PMS. Pleopod 5 exopod with<br />
~38 PMS. Exopods <strong>of</strong> pleopods –3 each with distolateral<br />
margin digitate; endopods <strong>of</strong> pleopods 3–5<br />
each without distolateral point; pleopods 2–4 peduncle<br />
distolateral margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about one-half as long as endopod. Endopod<br />
apically shallowly bifid, lateral margin proximally<br />
convex, without prominent excision, proximal lateral<br />
margin with 0 RS, distal lateral margin with RS, mesial<br />
margin straight, with 3–4 RS. Exopod extending to<br />
end <strong>of</strong> endopod, 2.8 times as long as greatest width,<br />
apically sub-bifid, mesial process prominent; lateral<br />
margin convex, with 8 RS; mesial margin convex, with<br />
3 RS.<br />
femaLe: Similar to males, but wider (2. times as long as<br />
greatest width at pereonite 6); the pleon is proportionally<br />
shorter ( 8% BL) than in the male (2 % BL).<br />
02<br />
size: Average lengths <strong>of</strong> type material: males 7.7 mm (n<br />
= 6), ovigerous females 2.0 mm (n = 6), non-ovigerous<br />
females 2.8 mm (n = 22); emergent mancas measured<br />
3.0–4.3 mm, and eggs measured .4– .6 mm.<br />
Variation: Robust setae: (n = 5) pleotelson always<br />
without RS. Uropod exopod mesial margin 3 (50%)<br />
or 4 (40%), 2 and 5 occurring, lateral margin 9 (83%),<br />
0 RS occurring once, 8 RS 3 times (possibly owing<br />
to damage); uropod endopod mesial margin with 2<br />
(20%) or 3 (73%) 4 and 5 occurring once each, lateral<br />
margin with only large RS distal to the pappose seta.<br />
<strong>The</strong>re is no discernable difference between males and<br />
females. <strong>The</strong> most distal robust setae are <strong>of</strong>ten very<br />
fine and occasionally absent, accounting for most <strong>of</strong><br />
the observed variation.<br />
<strong>The</strong> lateral margin <strong>of</strong> the uropodal endopod has<br />
a small robust seta set inside the bifid apex. This seta<br />
is largely obscured by the apical slender setae, and<br />
proved to be too small to observe by light microscopy<br />
and was not counted.<br />
<strong>The</strong> small size <strong>of</strong> the robust setae on pereopods<br />
and 2 precluded making accurate direct counts <strong>of</strong> pereopod<br />
robust setae using light microscopy.<br />
remarks: Aegiochus gordoni is characterised by having<br />
large but well-separated eyes, the antennule flagellum<br />
extending to pereonite 2 and a shield-shaped pleo-<br />
telson, which is finely serrate and totally lacking robust<br />
setae or notches; the uropods are provided with small<br />
and fine robust setae, the smallest <strong>of</strong> which are difficult<br />
to observe under light microscopy; the uropodal endopod<br />
lateral margin is markedly straight. <strong>The</strong> anterior<br />
pereopods are weakly ‘spined’ with the small robust<br />
setae close-set to the inferior margin, particularly on<br />
the propodus; the pleopods have the exopod disto-<br />
lateral margin with digitate serrations, and the appendix<br />
masculina extends well beyond the distal margin<br />
<strong>of</strong> the endopod.<br />
<strong>The</strong>re are three Southern Ocean species which appear<br />
similar. All <strong>of</strong> these species are described from<br />
single specimens only, and most descriptions lack<br />
sufficient detail for accurate comparisons to be made.<br />
<strong>The</strong>se species are Aegiochus crozetensis (Kussakin &<br />
Vasina, 982) (Crozet Islands, southern Indian Ocean,<br />
280 metres), Aegiochus pushkini Kussakin and Vasina,<br />
982 (Ob’ Bank, southern Indian Ocean, 4 0 m; here<br />
recorded from <strong>New</strong> <strong>Zealand</strong>) and Aegiochus uschakovi<br />
Kussakin, 967 (Drake Passage, Argentina, 95– 20<br />
m). All species share a similar appearance, but were<br />
described as having pleotelson ‘denticles’, taken here<br />
to mean robust setae.<br />
Aegiochus pushkini, redescribed herein, is nearly<br />
twice the size <strong>of</strong> A. gordoni, has narrower eyes, the pleopod<br />
rami lack marginal serrations and the pleotelson<br />
and uropods are provided with robust setae.
<strong>New</strong> figures <strong>of</strong> the holotypes <strong>of</strong> Aegiochus crozetensis<br />
and A. uschakovi are given (Appendix 2; Figs 4 , 45<br />
respectively). A. pushkini has a far shorter antennule<br />
flagellum, figured as extending only to the anterior <strong>of</strong><br />
pereonite . Aegiochus uschakovi, at 8 mm, is substantially<br />
larger than A. gordoni, and is figured as having<br />
the posterior coxae far more strongly produced, the<br />
antennule flagellum with 15 articles (10 in A. gordoni),<br />
antennal flagellum with 19 articles (11 in A. gordoni)<br />
and the propodal palm as lacking setae. Examination<br />
<strong>of</strong> the type material <strong>of</strong> these Southern Ocean species<br />
confirms that they are all distinct from A. gordoni.<br />
<strong>The</strong> deeply serrate pleopod exopod is not unique<br />
to A. gordoni, and has been figured for several other<br />
<strong>New</strong> <strong>Zealand</strong> species, the most similar being Aegiochus<br />
coroo and Aegiochus laevis. Aegiochus coroo has the eyes<br />
united, and a more rounded and notched posterior<br />
margin to the pleotelson. <strong>The</strong>re are further similarities<br />
between these three species in the unarmed pleotelson,<br />
a large molar process on the mandible, maxillule lateral<br />
lobe which has one prominently large and several small<br />
robust setae (in contrast to the more usual three large<br />
and several small terminal setae) and the anterior pereopods<br />
with very few and small robust setae. Aegiochus<br />
laevis is small, and readily separated from A. gordoni by<br />
having small eyes, somewhat similar in size to those<br />
<strong>of</strong> cirolanids.<br />
prey: Not known.<br />
distribution: Recorded from the Chatham Rise to the<br />
east <strong>of</strong> South Island and Brothers Sea Mounts to the<br />
northeast <strong>of</strong> North Island; at depths from 360 to 40<br />
metres (the minimum depth was recorded from a haul<br />
that ran from 360 to 755 metres, all other records are<br />
from 698 metres or greater).<br />
etymoLogy: Named for Dennis Gordon, scientist,<br />
natural historian, biological enthusiast and untiring<br />
advocate for taxonomic research in <strong>New</strong> <strong>Zealand</strong>—in<br />
recognition <strong>of</strong> his huge direct and indirect contribution<br />
to knowledge <strong>of</strong> the <strong>New</strong> <strong>Zealand</strong> marine fauna.<br />
Aegiochus insomnis sp. nov. (Figs 56–60)<br />
materiaL: Holotype: ♂ (6.9 mm) Poor Knights Islands,<br />
pass south <strong>of</strong> landing, 9 May 969, 33–37 m, from<br />
sponge and [bryozoa], SCUBA (AK 73308).<br />
Paratypes: 2 ♀ (ovig. 7.2, 7.8, 8.0 [dissected], 8.4, 9.0,<br />
9.5; non-ovig. 6.5, 6.8, 7.0, 7.5, 8.4, 9.0 [dissected] mm),<br />
same data as holotype (AK 73309; NIWA 28462 [2]).<br />
desCription: Body 2.2 times as long as greatest width,<br />
dorsal surfaces polished in appearance and sparsely<br />
punctate, widest at pereonite 5, lateral margins ovate.<br />
Rostral point folded ventrally and posteriorly. Eyes<br />
large, not medially united, separated by about 23%<br />
width <strong>of</strong> head; each eye made up <strong>of</strong> ~ 0 transverse<br />
rows <strong>of</strong> ommatidia, each row with ~8 ommatidia; eye<br />
colour red. Pereonite 1 and coxae 2–3 each with posteroventral<br />
angle right-angled (point s<strong>of</strong>t, not abrupt).<br />
Coxae 5–7 with entire oblique carina; posterior margins<br />
straight and sinuate (coxa 7 is sinuate), posterolateral<br />
angle acute (less than 45°) (coxa 5 not acute). Pleon with<br />
pleonite largely concealed by pereonite 7; pleonite 4<br />
with posterolateral margins not extending to posterior<br />
margin <strong>of</strong> pleonite 5; pleonite 5 with posterolateral<br />
angles free, not overlapped by lateral margins <strong>of</strong> pleonite<br />
4. Pleotelson 0.8 times as long as anterior width,<br />
dorsal surface without longitudinal carina; lateral margins<br />
convex, serrate, posterior margin evenly rounded,<br />
with 0 RS.<br />
Antennule peduncle articles 3 and 4 . times as<br />
long as combined lengths <strong>of</strong> articles and 2, article<br />
3 3.1 times as long as wide; flagellum with 8 articles,<br />
extending to posterior <strong>of</strong> pereonite . Antenna peduncle<br />
article 4 2.0 times as long as wide, 0.8 times as long<br />
as combined lengths <strong>of</strong> articles –3, inferior margin 0<br />
plumose setae, and 0 short simple setae (anterodistal<br />
angle with 5 long simple setae); article 5 .2 times as<br />
long as article 4, 2.6 times as long as wide, inferior margin<br />
with 2 palmate setae (at distal angle), anterodistal<br />
angle with cluster <strong>of</strong> 4 short simple setae (long simple<br />
setae, and 1 pappose seta); flagellum with 13 articles,<br />
extending to posterior <strong>of</strong> pereonite 2.<br />
Frontal lamina flat, longer than greatest width, diamond<br />
shaped, anterior margin acute, without small<br />
median point or forming median angle, posterior<br />
margin not abutting clypeus.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 5 distolateral setae (3 biserrate,<br />
2 simple), palp article 3 with 3 setae (serrate; distal 3<br />
markedly longer than remainder). Maxillule with 5 terminal<br />
RS ( large, 4 slender). Maxilla mesial lobe with 2<br />
RS ( biserrate); lateral lobe with 3 RS. Maxilliped endite<br />
with 0 apical setae; palp article 2 with 2 RS (slender);<br />
article 3 with 4 straight RS; article 4 with 4 hooked RS<br />
(and simple seta); article 5 with 3 RS ( large, and 2<br />
small slender).<br />
Pereopod 1 basis 2.6 times as long as greatest width;<br />
ischium 0.3 times as long as basis, inferior margin<br />
with 0 RS, superior distal margin with RS; merus<br />
inferior margin with RS (plus 2 short and long<br />
simple setae), set as distal group, superior distal angle<br />
with 0 RS ( simple seta); carpus 0.8 as long as merus,<br />
inferior margin with RS; propodus 2.6 times as long<br />
as proximal width, inferior margin with 2 RS (set as<br />
minute and distally), propodal palm simple, without<br />
blade or process, dactylus smoothly curved, 0.9 as long<br />
as propodus. Pereopod 2 ischium inferior margin with<br />
0 RS, superior distal margin with RS; merus inferior<br />
margin with 3 RS (set as and 2; 2 simple setae), set<br />
03
Figure 56. Aegiochus insomnis sp. nov. A–F, holotype, remainder female paratype (9.0 mm). A, dorsal view; B, lateral view;<br />
C, head; D, frons, ventral view; E, pleotelson, posterior margin; F, penial openings; G, antennule (terminal flagellar articles<br />
missing); H, antenna peduncle.<br />
04
Figure 57. Aegiochus insomnis sp. nov. Female paratype (9.0 mm). A, mandible; B, right mandible, incisor and molar process; C,<br />
mandible palp article 3; D, maxillule; E, maxillule apex; F, maxilla; G, maxilla apex; H, maxilliped; I, maxilliped articles 2–5.<br />
as two groups, superior distal margin with acute RS<br />
(and 2 simple setae); carpus similar in size to that <strong>of</strong><br />
pereopod , inferodistal angle with RS (and simple<br />
seta). Pereopod 3 similar to pereopod 2 (but merus<br />
inferior margin with 5 RS, and RS larger). Pereopods<br />
5–7 inferior margins <strong>of</strong> ischium–carpus with short RS.<br />
Pereopod 6 similar to pereopod 7 (but more robust, distal<br />
margins <strong>of</strong> merus and carpus with more numerous<br />
biserrate RS). Pereopod 7 basis 3.0 as long as greatest<br />
width, inferior margins with 7 palmate setae; ischium<br />
0.4 as long as basis, inferior margin with 3 RS (set<br />
singly), superior distal angle with 4 RS, inferior distal<br />
angle with 4 RS; merus . as long as ischium, 2.2 times<br />
as long as wide, inferior margin with 3 RS (set as and<br />
05
Figure 58. Aegiochus insomnis sp. nov. Female paratype (9.0 mm). A–E, pereopods –3, 6 and 7 respectively.<br />
2), superior distal angle with 7 RS, inferior distal angle<br />
with 6 RS; carpus .3 as long as ischium, 2.8 times as<br />
long as wide, inferior margin with 4 RS (set as ,<br />
and 2), superior distal angle with 8 RS (5 biserrate),<br />
inferior distal angle with 6 RS; propodus .0 as long<br />
as ischium, 3.6 times as long as wide, inferior margin<br />
with 4 RS (set as , and 2), superior distal angle with<br />
3 slender setae ( RS, plumose and 2 simple), inferior<br />
distal angle with 4 RS.<br />
Penes low tubercles; penial openings separated by<br />
% <strong>of</strong> sternal width.<br />
06<br />
Pleopod 1 exopod .9 times as long as wide, distally<br />
narrowly rounded, mesial margin weakly oblique,<br />
lateral margin weakly convex, mesial margin strongly<br />
convex, with PMS on distal half, with ~36 PMS; endopod<br />
.9 times as long as wide, distally rounded, lateral<br />
margin straight, with PMS on distal one-third, mesial<br />
margin with PMS on distal margin only, endopod with<br />
~ 6 PMS; peduncle .9 times as wide as long, mesial<br />
margin with 5 coupling hooks. Pleopod 2 exopod with<br />
~37 PMS, endopod with ~ 8 PMS; appendix masculina<br />
basally swollen, .3 times as long as endopod, distally
Figure 59. Aegiochus insomnis sp. nov. A, B, holotype, remainder female paratype (9.0 mm). A–D pleopods –3, 5 respectively;<br />
E, uropod; F, uropod endopod, apex; G, uropod exopod, apex.<br />
07
Figure 60. Aegiochus insomnis sp. nov. Ovigerous female paratype 8.0 mm. A, maxilliped; B, maxilliped palp articles 3–5; C,<br />
oostegite, sternite 5 (anterior to top <strong>of</strong> page); D, setae from mesial margin <strong>of</strong> oostegite; E, setae from anterolateral margin <strong>of</strong><br />
oostegite.<br />
acute. Pleopod 3 exopod with ~40 PMS, endopod with<br />
~6 PMS. Pleopod 4 exopod with ~37 PMS, endopod with<br />
~5 PMS. Pleopod 5 exopod with ~37 PMS. Exopods <strong>of</strong><br />
pleopods –3 each with distolateral margin digitate;<br />
endopods <strong>of</strong> pleopods 3–5 each without distolateral<br />
point; pleopods 2–4 peduncle distolateral margin with<br />
prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 3 RS,<br />
posterior lobe about one-half as long as endopod.<br />
Endopod apically deeply and equally bifid, lateral<br />
margin straight, without prominent excision, proximal<br />
lateral margin with 0 RS, distal lateral margin with<br />
RS, mesial margin weakly convex, with 3 RS. Exopod<br />
not extending to end <strong>of</strong> endopod, 2.4 times as long as<br />
greatest width, apically deeply bifid, mesial process<br />
prominent; lateral margin convex, with 7 RS; mesial<br />
margin convex, with 3 RS.<br />
femaLe: Similar to males, ovigerous females slightly<br />
wider and slightly larger. Brood pouch composed <strong>of</strong> 5<br />
pairs <strong>of</strong> oostegites arising from sternites –5, becoming<br />
progressively larger towards the posterior, oostegite<br />
5 extending to the posterior <strong>of</strong> sternite and bearing<br />
plumose seta on the mesial margin, stout simple setae<br />
along the posterior margin.<br />
size: <strong>The</strong> single male measured 6.9 mm; ovigerous<br />
females 7.2–9.5 mm (mean = 8.3 mm); non-ovigerous<br />
females 6.5–9.0 mm (mean = 7.5 mm).<br />
08<br />
Variation: Pleotelson (n = 2) always without RS.<br />
Uropod exopod mesial (n = 22) margin usually with 3<br />
(82%), or 2 ( 8%) RS, lateral margin (n = 24) with 7 (88%)<br />
or 8 ( 2%) RS; uropod endopod mesial margin (n = 24)<br />
with 0 (38%), (33%) or 2 (29%) RS, lateral margin with<br />
only (88%) or 0 ( 2%) RS distal to the pappose seta.<br />
<strong>The</strong>re is no discernable difference between males and<br />
females. <strong>The</strong> distal robust setae are frequently small<br />
and fine.<br />
<strong>The</strong> relatively small size <strong>of</strong> the species precluded<br />
making accurate direct counts <strong>of</strong> the robust setae on<br />
pereopods and 2 although <strong>of</strong> the specimens examined<br />
the pattern, number and size appears consistent.<br />
remarks: Aegiochus insomnis sp. nov. is readily identified<br />
by the relatively small eyes, diamond-shaped and<br />
flat (‘Metacirolana-like’) frontal lamina, short pereopod<br />
dactylus (about as long as the propodus), deeply serrated<br />
pleopod exopods and the long acute appendix<br />
masculina. <strong>The</strong>re are two similar species in <strong>New</strong> <strong>Zealand</strong>:<br />
Aegiochus nohinohi sp. nov. and Aegiochus gordoni<br />
sp. nov. Both <strong>of</strong> those species have larger eyes, bladelike<br />
posterior margin on the frontal lamina and have<br />
a much longer dactylus on pereopod ( .2– .4 times<br />
as long as the propodus).<br />
prey: Not known.
distribution: Known only from the Poor Knights Islands,<br />
northern <strong>New</strong> <strong>Zealand</strong>.<br />
etymoLogy: <strong>The</strong> epithet insomnis (from the L. meaning<br />
sleepless; noun in apposition) is a play on the name <strong>of</strong><br />
the type locality.<br />
Aegiochus kakai sp. nov. (Figs 6 –64)<br />
materiaL examined: Holotype. ♂ ( 4.2 mm), Chatham<br />
Rise, 42°43.20’S, 79°57.63’W, 2 April 200 , 0 2–890<br />
m, coll. RV Tangaroa (NIWA 23863).<br />
Paratypes. All Chatham Rise. 4 ♂ ( 2.5 [damaged],<br />
3.6, 4.5 dissected, 6.0 mm), 5 ♀ (ovig. 5.0, 5.5; nonovig.<br />
2.5, 5.5, 24.5), same data as holotype (NIWA<br />
23864). 6 ♂ ( .0. .2, 2.2, 2.8, 3.5, 4.5 mm), 4 ♀<br />
(ovig. 3.8, 5.0, 6.5; non-ovig. 6.5 mm), 5 April 200 ,<br />
42°42.84’S, 79°57.5 ’W, 980–893 m, coll. RV Tangaroa<br />
(NIWA 23865). ♀ (non-ovig. 8.5 mm), Chatham Rise,<br />
5 April 200 , 42°45.93’S, 79°59.34’W, 875–757 m, coll.<br />
RV Tangaroa (NIWA 23866); 2 ♀ (ovig. 4.5, 5.0 mm),<br />
6 April 200 , 42°47. 7’S, 79°59. 2’W, 993–900 m, coll.<br />
RV Tangaroa (NIWA 23867).<br />
Non-type material, some not measured. ♀ (non-ovig.),<br />
7 April 200 , 42°48.24’S, 79°59.27’E, 0 3–93 m<br />
(NIWA 23868). 2 ♀ (ovig.), 2 April 200 , 42°42.76’S,<br />
79°54.45’W, 080– 008 m (NIWA 23869). ♀ (ovig.), 8<br />
April 200 , 42°47.27’S, 79°59.8 ’W, 042–880 m (NIWA<br />
23870). ♀ (ovig. 5.7 mm) 6 July 994, 43°5 .47’S,<br />
74° 7.08’W, 754 m (NIWA 23870). 2, 42.7275–7307°S,<br />
80. 0 0–0973°E, 7 June 2006, 000– 07 m, (NIWA<br />
25670, 2567 ). , 42.7 58–7 32°S, 80. 432–0480°E,<br />
4 June 2006, 950– 070 m (NIWA 25668). , 42.7627–<br />
7575°S, 80.0748–0773°E, 28 May 2006, 0 9– 08 m<br />
(NIWA 25656).<br />
desCription: Body 2. times as long as greatest width,<br />
dorsal surfaces polished in appearance, widest at pereonite<br />
5, lateral margins weakly ovate. Rostral point<br />
folded ventrally and posteriorly. Eyes large, medially<br />
united, anterior clear field 27% length <strong>of</strong> head, posterior<br />
clear field 44% length <strong>of</strong> head; each eye made up<br />
<strong>of</strong> ~ 6 transverse rows <strong>of</strong> ommatidia, each row with<br />
~9 ommatidia; eye colour red, or dark brown. Pereonite<br />
1 and coxae 2–3 each with posteroventral angle with<br />
small distinct produced point. Coxae 5–7 with entire<br />
oblique carina; posterior margins concave, posterolateral<br />
angle acute (less than 45°). Pleon with pleonite<br />
visible in dorsal view; pleonite 4 with posterolateral<br />
margins extending clearly beyond posterior margin <strong>of</strong><br />
pleonite 5; pleonite 5 with posterolateral angles overlapped<br />
by lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.7<br />
times as long as anterior width, dorsal surface without<br />
longitudinal carina; lateral margins convex, serrate or<br />
notched, posterior margin converging to caudomedial<br />
point, with 8– 0 RS.<br />
Antennule peduncle articles 3 and 4 . times as<br />
long as combined lengths <strong>of</strong> articles and 2, article 3<br />
4.1 times as long as wide; flagellum with 10 articles,<br />
extending to posterior <strong>of</strong> pereonite . Antenna peduncle<br />
article 4 2.9 times as long as wide, 0.9 times as long as<br />
combined lengths <strong>of</strong> articles –3, inferior margin with<br />
plumose seta, and 4 short simple setae (anterodistal<br />
3 simple + plumose); article 5 .0 times as long as article<br />
4, 2.7 times as long as wide, inferior margin with 3<br />
palmate setae, anterodistal angle with cluster <strong>of</strong> 3 short<br />
simple setae; flagellum with 15 articles, extending to<br />
posterior <strong>of</strong> pereonite 3.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, posterior<br />
margin concave, anterior margin anteriorly truncate<br />
(narrowly), forming median angle, posterior margin<br />
not abutting clypeus.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with distolateral setae (4 large<br />
and 5 small biserrate, 2 simple distally), palp article 3<br />
with 20 setae (all biserrate). Maxillule with 5 terminal<br />
RS ( large, 4 slender). Maxilla mesial lobe with 3 RS (<br />
straight, 2 biserrate); lateral lobe with 2 RS (hooked).<br />
Maxilliped endite with 0 apical setae; palp article 2<br />
with 2 RS; article 3 with 4 recurved RS (2 slender, 2<br />
hooked); article 4 with 4 hooked RS; article 5 with 4<br />
RS (straight).<br />
Pereopod 1 basis 2.7 times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with RS; merus inferior<br />
margin with 2 RS (minute), set as two groups, superior<br />
distal angle with 0 RS (2 short slender setae); carpus 0.7<br />
as long as merus, inferior margin with RS; propodus<br />
2.3 times as long as proximal width, inferior margin<br />
with 0 RS, propodal palm simple, without blade or<br />
process, dactylus smoothly curved, .4 as long as propodus.<br />
Pereopod 2 ischium inferior margin with 0 RS,<br />
superior distal margin with RS; merus inferior margin<br />
with 4 RS (distalmost large, remainder minute), set as<br />
two groups, superior distal margin with acute RS<br />
(small plus 2 slender setae); carpus longer than that <strong>of</strong><br />
pereopod , with inferodistal lobe, inferodistal angle<br />
with RS. Pereopod 3 similar to pereopod 2. Pereopods<br />
5–7 inferior margins <strong>of</strong> ischium–carpus with short<br />
RS. Pereopod 6 similar to pereopod 7 (but longer, with<br />
slightly more RS). Pereopod 7 basis 3.6 times as long<br />
as greatest width, inferior margins with palmate<br />
setae; ischium 0.4 as long as basis, inferior margin<br />
with 3 RS (set as , and ), superior distal angle with<br />
2 RS, inferior distal angle with 4 RS; merus .2 as long<br />
as ischium, 2.5 times as long as wide, inferior margin<br />
with 3 RS (set as and 2), superior distal angle with 7<br />
09
Figure 61. Aegiochus kakai sp. nov. A–G, holotype, remainder male paratype NIWA 23864. A, dorsal view; B, lateral view; C,<br />
head; D, frons; E, frons, anterior view; F, pleotelson; G, pleotelson, posterior margin; H, sternite 7 showing penial openings;<br />
I, antennule; J, antenna peduncle.<br />
0
Figure 62. Aegiochus kakai sp. nov. Male paratype 4.5 mm, NIWA 23864. A, mandible; B, mandible palp, article 3; C, maxillule<br />
apex; D, maxilla; E, maxilla apex; F, maxilliped; G, maxilliped articles 2–5.<br />
RS, inferior distal angle with 6 RS; carpus .0 as long<br />
as ischium, 2.6 times as long as wide, inferior margin<br />
with 3 RS (set as and 2), superior distal angle with 9<br />
RS (6 <strong>of</strong> which are biserrate), inferior distal angle with<br />
5 RS; propodus .0 as long as ischium, 4 times as long<br />
as wide, inferior margin with 5 RS (seta as , 2 and 2),<br />
superior distal angle with 3 slender setae, inferior distal<br />
angle with 3 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7, mutually<br />
adjacent.<br />
Pleopod 1 exopod 2.0 as long as wide, distally narrowly<br />
rounded, mesial margin weakly oblique, lateral<br />
margin straight, mesial margin strongly convex, with<br />
PMS on distal half, with ~54 PMS; endopod 2.2 times as<br />
long as wide, distally rounded, lateral margin straight,<br />
with PMS on distal half, mesial margin with PMS on<br />
distal one-third, endopod with ~33 PMS; peduncle .8<br />
times as wide as long, mesial margin with 7 coupling<br />
hooks. Pleopod 2 exopod with ~66 PMS, endopod with<br />
~43 PMS; appendix masculina basally swollen, . times<br />
as long as endopod (middle part with prominent cuticular<br />
scales), distally acute. Pleopod 3 exopod with ~75<br />
PMS, endopod with ~ 5 PMS. Pleopod 4 exopod with<br />
~70 PMS, endopod with ~ 0 PMS. Pleopod 5 exopod
Figure 63. Aegiochus kakai sp. nov. Male paratype 4.5 mm, NIWA 23864. A–E, pereopods , 2, 6 and 7 respectively.<br />
with ~60 PMS. Exopods <strong>of</strong> pleopods –3 each with<br />
distolateral margin not digitate; endopods <strong>of</strong> pleopods<br />
3–5 each with distolateral point; pleopods 2–4 peduncle<br />
distolateral margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS<br />
(and lateral), posterior lobe about one-half as long<br />
as endopod. Endopod apically sub-bifid, mesial process<br />
prominent, lateral margin straight, without prominent<br />
excision, proximal lateral margin with RS, distal lateral<br />
margin with 2 RS, mesial margin weakly convex,<br />
with 7 RS. Exopod extending beyond end <strong>of</strong> endopod<br />
2<br />
(slightly), 3.5 times as long as greatest width, apically<br />
sub-bifid, mesial process prominent; lateral margin<br />
convex, with RS; mesial margin straight, distally<br />
convex, with 4 RS.<br />
femaLe: Ovigerous females have the BL .8 times as<br />
long as the greatest width, with ovate lateral margins;<br />
otherwise similar in appearance to males other than for<br />
the sexual characters. Brood pouch <strong>of</strong> oostegites arising<br />
from the coxae/sternite <strong>of</strong> pereopods –5.
Figure 64. Aegiochus kakai sp. nov. Male paratype 4.5 mm, NIWA 23864. A–D, pleopods –3, 5 respectively; E, uropod<br />
endopod apex; F, uropod.<br />
size: Males .0– 6.0 mm (mean = 3.3 mm, n = );<br />
ovigerous females 3.8– 6.5 mm (mean = 5.0 mm,<br />
n = 7); non-ovigerous females 2.5–24.5 mm (mean =<br />
7.5 mm, n = 5).<br />
Variation: Robust setae: pleotelson RS (n = 8) varies<br />
from 4+4 (22%) or 4+5 (39%) and 5+5 (28%) with 4+6<br />
and 6+6 each occurring once. Uropod exopod (n = 36)<br />
mesial margin most <strong>of</strong>ten with 4 (25%) or 5 (64%) or<br />
3
3 and 6 each occurring twice; lateral margin ( %)<br />
or 2 (6 %) or 3 (28%); uropod endopod (n = 36)<br />
mesial margin varied from 5 to 8 RS with 5 ( 4%), 6<br />
(25%) and 7 (58%) the most frequent, 8 occurring once,<br />
lateral margin with +2 (83%) with occasional damageinduced<br />
variations <strong>of</strong> + and 0+ ; one specimen had<br />
2+2. <strong>The</strong>re is no discernable difference between males<br />
and females.<br />
Pereopods present a constant appearance, but no<br />
detailed counts were made owing to the difficulty <strong>of</strong><br />
observing small setae under light microscopy without<br />
dissection.<br />
Recently (2006) collected and fresh material <strong>of</strong> this<br />
species from the Chatham Rise (NIWA 25656, 25668,<br />
25670, 2567 ) had bronze coloured eyes which are<br />
very narrowly (less than the width <strong>of</strong> an ommatidium)<br />
separated, this separation being less or not apparent in<br />
long-preserved specimens.<br />
remarks: <strong>The</strong> united eyes, acute and produced posterior<br />
margins <strong>of</strong> coxal plates 2–4, wide frontal lamina<br />
with the posterior margin forming a distinctly concave<br />
blade, distinctive long and straight appendix masculina<br />
in the males and the setation <strong>of</strong> robust setae on the<br />
uropods and pleotelson all allow ready identification<br />
<strong>of</strong> this species.<br />
Aegiochus kakai sp. nov. shares a great many characters<br />
with Aegiochus kanohi sp. nov., including the<br />
general appearance and setation <strong>of</strong> the antennule,<br />
antenna, pereopods, pleopods, uropods and posterior<br />
margin <strong>of</strong> the pleotelson. In particular the general<br />
morphology <strong>of</strong> the appendix masculina <strong>of</strong> the two species<br />
is similar, both being basally swollen and distally<br />
slender. Despite the overall similarity <strong>of</strong> appearance<br />
there are numerous clear-cut points <strong>of</strong> difference and<br />
the two species are easy to distinguish. Aegiochus kakai<br />
has the eyes meeting medially, but the eyes themselves<br />
are far smaller than in A. kanohi; the frontal lamina is<br />
far wider than in A. kanohi and the posterior margin is<br />
concave; the posteroventral angles <strong>of</strong> pereonite and<br />
coxae 2 and 3 are acute and produced (truncate in A.<br />
kanohi); and the appendix masculina is straight (sinuate<br />
in A. kanohi). <strong>The</strong>re are further differences in the<br />
setation <strong>of</strong> the uropods and posterior margin <strong>of</strong> the<br />
pleotelson, which is detailed in the ‘variation’ section<br />
for each species.<br />
Other superficially similar species from the southern<br />
Indian Ocean are Aegiochus crozetensis Kussakin<br />
and Vasina, 982 and A. uschakovi Kussakin, 967, but<br />
these both have widely separated eyes.<br />
prey: Not known. One sample (NIWA 23864) had<br />
sponge tissue tangled up with the specimens suggesting<br />
the possibility <strong>of</strong> at least temporary association<br />
with hexactinellid sponges.<br />
4<br />
distribution: Recorded only from the Chatham Rise region,<br />
<strong>of</strong>f eastern South Island, <strong>New</strong> <strong>Zealand</strong>; at depths<br />
from 757 to 080 metres.<br />
etymoLogy: Kakai is a Mäori word meaning to nibble<br />
or bite frequently; noun in apposition.<br />
Aegiochus kanohi sp. nov. (Figs 65–68)<br />
materiaL examined: Holotype: ♂ ( 4.8 mm), Chatham<br />
Rise, 42°45.89’S, 79°59. 6’W, 9 April 200 , 800–757<br />
m, coll. RV Tangaroa (NIWA 240 9).<br />
Paratypes: 7 ♂ ( 0.5, .0, .5, 2.0, 2.5 dissected,<br />
2.5, 3.0 mm), 2 ♀ (ovig. 4.0, 4.0, 5.0, 5.5, 5.5,<br />
5.6, 6.0; non-ovig. .6, 2.5, 5.0, 5.0, 6.0 mm),<br />
same data as holotype (NIWA 24020). ♀ (non-ovig<br />
0.5 mm), Chatham Rise, 6 April 200 , 42°45.76’S,<br />
79°59.29’W, 064–750, coll. RV Tangaroa (NIWA<br />
2402 ). ♂ ( 2.0 mm), Chatham Rise, 5 April 200 ,<br />
42°45.93’S, 79°59.34’W, 875–757 m, coll. RV Tangaroa<br />
(NIWA 24022).<br />
Additional material: ♂ (9.5 mm), ♀ ( 3.0 mm ovig),<br />
manca (7.5 mm), north <strong>of</strong> Chatham Rise, 3 September<br />
963, 43°04.00’S, 78°38.99’W, 549 m, stn. A9 0 (two<br />
tubes) (NIWA 24023). ♀ ( 5.8 mm, ovig, v. poor condition),<br />
vicinity <strong>of</strong> Hikurangi Trough, 5 September<br />
987, 39°5 .90’S, 77°25. 9’E, 4 3 m (NIWA 24024).<br />
<strong>New</strong> caledonia: 2 ♀ (ovig. 2.0, 3.2 mm), 24°39.3 ’S,<br />
68°39.67’E, 29 October 986, 600 m, CHALCAL II, stn<br />
DW75 (MNHN Is.59 ). 2 ♀ (non-ovig. 9.9, 2.9 mm),<br />
<strong>New</strong> Caledonia, 24°54.5’S, 68°22.3’E, 28 October 986,<br />
527 m, CHALCAL II, stn DW72 (MNHN Is.59 2).<br />
desCription: Body 2.5 times as long as greatest width,<br />
dorsal surfaces smooth, widest at pereonite 5, lateral<br />
margins weakly ovate. Rostral point folded ventrally<br />
and posteriorly. Eyes large, medially united (very narrow<br />
gap present), anterior clear field 5% length <strong>of</strong> head,<br />
posterior clear field 28% length <strong>of</strong> head; each eye made<br />
up <strong>of</strong> ~ 5 transverse rows <strong>of</strong> ommatidia, each row<br />
with ~9 ommatidia; eye colour black. Pereonite 1 and<br />
coxae 2–3 each with posteroventral angle right-angled.<br />
Coxae 5–7 with entire oblique carina (coxae 5–7 dorsal<br />
margin concave); posterior margins straight, posterolateral<br />
angle acute (less than 45°). Pleon with pleonite<br />
visible in dorsal view; pleonite 4 with posterolateral<br />
margins not extending to posterior margin <strong>of</strong> pleonite<br />
5; pleonite 5 with posterolateral angles free, not overlapped<br />
by lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.9<br />
times as long as anterior width, dorsal surface without<br />
longitudinal carina; lateral margins convex, serrate or<br />
notched, posterior margin converging to caudomedial<br />
point, with 8– 0 RS.<br />
Antennule peduncle articles 3 and 4 .0 as long<br />
as combined lengths <strong>of</strong> articles and 2, article 3 3.4<br />
times as long as wide (narrowing distally); flagellum
Figure 65. Aegiochus kanohi sp. nov. J, K, male paratype 2.5 mm NIWA 24020, remainder holotype. A, dorsal view; B, lateral<br />
view; C, head; D, frons; E, pleonite, lateral view; F, pleotelson, setation on posterior margin; G, pleotelson apex; H, sternite 7<br />
showing penial openings; I, frons, anterior view; J, antennule; K, antenna peduncle.<br />
5
Figure 66. Aegiochus kanohi sp. nov. Male paratype 2.5 mm NIWA 24020. A, mandible; B, mandible palp, article 3; C, maxillule;<br />
D, maxilla; E, maxilla apex; F, maxilliped; G, maxilliped articles 2–5.<br />
with 3 articles, extending to posterior <strong>of</strong> pereonite .<br />
Antenna peduncle article 4 .9 times as long as wide,<br />
0.8 times as long as combined lengths <strong>of</strong> articles –3,<br />
inferior margin 0 plumose setae, and 5 short simple<br />
setae (anterodistal angle); article 5 . times as long<br />
as article 4, 2.2 times as long as wide, inferior margin<br />
with 2 palmate setae (at distal angle), anterodistal<br />
angle with cluster <strong>of</strong> 6 short simple setae (simple and<br />
2 pappose); flagellum with 16 articles, extending to<br />
posterior <strong>of</strong> pereonite 3.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like (weakly blade-like, not flat),<br />
6<br />
wider than long, diamond shaped, anterior margin<br />
acute, forming median angle, posterior margin not<br />
abutting clypeus.<br />
Mandible molar process present, small distinct<br />
flat lobe; palp article 2 with 9 distolateral setae (large<br />
biserrate, 3 short simple), palp article 3 with 6 setae.<br />
Maxillule with 6 terminal RS (5 slender, large). Maxilla<br />
mesial lobe with 4 RS (3 serrate, simple); lateral lobe<br />
with 4 RS (3 hooked, straight). Maxilliped endite with<br />
apical seta; palp article 2 with 2 RS (hooked); article 3<br />
with 6 recurved RS (3 recurved, 2 simple, biserrate);
Figure 67. Aegiochus kanohi sp. nov. A, B, holotype, remainder paratype 2.5 mm NIWA 24020. C, male paratype, remainder<br />
holotype. A–D, pereopods , 2, 6 and 7 respectively; E, pereopod 6, dactylus.<br />
article 4 with 4 hooked RS; article 5 with 4 RS (straight,<br />
3 short, long and serrate).<br />
Pereopod 1 basis 3.5 times as long as greatest width;<br />
ischium 0.3 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with RS; merus inferior<br />
margin with 0 RS, superior distal angle with 0 RS (<br />
simple and plumose slender setae); carpus 0.6 as<br />
long as merus, inferior margin with 0 RS; propodus<br />
2.7 times as long as proximal width, inferior margin<br />
with 0 RS, propodal palm simple, without blade or<br />
7
Figure 68. Aegiochus kanohi sp. nov. A, B, holotype, remainder paratype 2.5 mm NIWA 24020. A–D, pleopods –3, 5,<br />
respectively; E, uropod; F, uropod exopod, apex.<br />
process, dactylus abruptly hooked, .4 as long as propodus.<br />
Pereopod 2 ischium inferior margin with 0 RS,<br />
superior distal margin with RS; merus inferior margin<br />
with 2 RS (minute, set as and ), set as two groups,<br />
8<br />
superior distal margin with 0 acute RS ( simple and<br />
plumose slender setae); carpus similar in size to that<br />
<strong>of</strong> pereopod , inferodistal angle with RS. Pereopod 3<br />
similar to pereopod 2. Pereopods 5–7 inferior margins
<strong>of</strong> ischium–carpus with short RS. Pereopod 6 similar to<br />
pereopod 7 (slightly longer with slightly more RS).<br />
Pereopod 7 basis 2.9 times as long as greatest width,<br />
inferior margins with 8 palmate setae; ischium 0.5 as<br />
long as basis, inferior margin with 2 RS (as and ),<br />
superior distal angle with 3 RS, inferior distal angle<br />
with 4 RS; merus .2 as long as ischium, 3.6 times as<br />
long as wide, inferior margin with 2 RS (set as and<br />
), superior distal angle with 6 RS, inferior distal angle<br />
with 5 RS; carpus . as long as ischium, 3.5 times as<br />
long as wide, inferior margin with 3 RS (set as and<br />
2), superior distal angle with 6 RS, inferior distal angle<br />
with 7 RS; propodus 0.9 as long as ischium, 4.7 times<br />
as long as wide, inferior margin with 3 RS (set as and<br />
2), superior distal angle with 3 slender setae (2 slender,<br />
plumose), inferior distal angle with 3 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by 5% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod 2.2 times as long as wide, distally<br />
narrowly rounded, mesial margin weakly oblique,<br />
lateral margin straight, mesial margin strongly convex,<br />
with PMS on distal half, with ~50 PMS; endopod 2.<br />
times as long as wide, distally rounded, lateral margin<br />
weakly concave, with PMS on distal one-third, mesial<br />
margin with PMS on distal one-third, endopod with<br />
~30 PMS; peduncle .6 times as wide as long, mesial<br />
margin with 6 coupling hooks (and 2 PMS). Pleopod 2<br />
exopod with ~65 PMS, endopod with ~38 PMS; appendix<br />
masculina basally swollen (sinuate), .2 times as long<br />
as endopod, distally acute. Pleopod 3 exopod with ~68<br />
PMS, endopod with ~ 8 PMS. Pleopod 4 exopod with<br />
~65 PMS, endopod with ~ 3 PMS. Pleopod 5 exopod<br />
with ~55 PMS. Exopods <strong>of</strong> pleopods –3 each with<br />
distolateral margin not digitate; endopods <strong>of</strong> pleopods<br />
3–5 each with distolateral point; pleopods 2–4 peduncle<br />
distolateral margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 3 RS<br />
(and single slender seta), posterior lobe about one-third<br />
as long as endopod. Endopod apically not bifid, lateral<br />
margin straight, without prominent excision, proximal<br />
lateral margin with RS, distal lateral margin with<br />
RS, mesial margin weakly convex, with 7 RS. Exopod<br />
not extending to end <strong>of</strong> endopod, 3.0 as long as greatest<br />
width, apically not bifid; lateral margin weakly convex,<br />
with RS; mesial margin weakly convex, with 5 RS.<br />
femaLe: Ovigerous females’ BL is twice as long as the<br />
greatest width, with ovate lateral margins; otherwise<br />
similar in appearance to males other than for the sexual<br />
characters. Brood pouch <strong>of</strong> oostegites arising from the<br />
coxae/sternite <strong>of</strong> pereopods –5.<br />
size: Males 9.5– 4.8 mm (mean = 2.2 mm, n = 8);<br />
ovigerous females 3.0– 6.0 mm (mean = 5. mm,<br />
n = 7); non-ovigerous females .6– 6.0 mm (mean =<br />
4.0 mm, n = 5).<br />
Variation: Robust setae: (n = 20) pleotelson RS 4+4 (60%)<br />
or 5+5 (30%) with 4+5 occurring twice; apical pair <strong>of</strong><br />
setae small; the pleotelson apex is <strong>of</strong>ten damaged.<br />
Uropod exopod (n = 37) mesial margin with 4 (27%), 5<br />
(49%) or 6 ( 9%) with 3 twice; lateral margin (27%)<br />
or 2 (68%), one specimen with 3; uropod endopod<br />
mesial (n = 38) margin varied from 5 to 8 RS with 6<br />
(45%) or 7 (36%) the most frequent, lateral margin with<br />
+ on all but one specimen. <strong>The</strong>re is no discernable<br />
difference between males and females, nor does the<br />
number <strong>of</strong> RS increase with the size <strong>of</strong> the specimen.<br />
remarks: <strong>The</strong> united eyes, truncate posterior margins<br />
<strong>of</strong> coxal plates 2 and 3, noticeably slender pereopods,<br />
relatively narrow frontal lamina with the posterior<br />
margin forming a transverse ridge rather than a distinct<br />
blade, distinctive long and sinuate appendix masculina<br />
in the males and the setation <strong>of</strong> robust setae on the<br />
uropods and pleotelson all allow ready identification<br />
<strong>of</strong> A. kanohi.<br />
<strong>The</strong> only similar <strong>New</strong> <strong>Zealand</strong> species with large<br />
eyes is Aegiochus kakai sp. nov., and that species is immediately<br />
distinguished by having far smaller eyes,<br />
a wider frontal lamina, the clear space distal to the<br />
palmate seta on the uropod endopod lateral margin is<br />
straight (concave in A. kakai), the posteroventral angles<br />
<strong>of</strong> pereonite and coxae 2–4 each with prominent and<br />
acute points, and a straight appendix masculina.<br />
Other similar species are Aegiochus coroo (Bruce,<br />
983) from southeastern Australia and Aegiochus synopthalma<br />
(Richardson, 909) from Japan. <strong>The</strong> former<br />
lacks robust setae on the pleotelson, has more robust<br />
pereopods, pleopodal exopods are strongly digitate<br />
and the appendix masculina is shorter and straight. <strong>The</strong><br />
latter species, known only from the female holotype,<br />
has a longer frontal lamina, the uropod endopod lateral<br />
margin has 4 robust setae (compared to 2 in A. kanohi<br />
sp. nov.) as well as having fewer robust setae on the<br />
uropodal exopod lateral margin (8 v. 2).<br />
prey: Not known.<br />
distribution: Recorded from the Chatham Rise, <strong>of</strong>f<br />
eastern South Island, and east <strong>of</strong> Hawkes Bay, North<br />
Island, <strong>New</strong> <strong>Zealand</strong>; also <strong>of</strong>f southern <strong>New</strong> Caledonia;<br />
at depths from to 4 3 to 064 metres.<br />
etymoLogy: Kanohi is a Mäori word for ‘eye’; noun in<br />
apposition.<br />
9
Aegiochus laevis (Studer, 884), comb. nov.<br />
(Figs 69, 70)<br />
Cirolana laevis Studer, 1884*: 21, pl. II, fig. 8.– Hale, 1925:<br />
45.– Nierstrasz, 93 : 57.– Bruce, 98 : 96 .<br />
Aega novi-zealandiae.– Tattersall, 1921: 213, pl. IV, figs 11–14.–<br />
Hurley, 1961: 268 [misidentification].<br />
Aega (Ramphion) laevis.– Brusca, 983: .– Bruce, 983: 763,<br />
figs 5, 6.<br />
Aega laevis.– Bruce, Lew Ton & Poore, 2003: 6 .<br />
materiaL examined: ♀ (ovig. 0.2 mm), <strong>of</strong>f North Cape,<br />
2 March 968, 34°39.00’S, 72° 3.99’E, 2 6 m (NIWA<br />
23763). ♀ (ovig. 3.5 mm), 7 miles east <strong>of</strong> North Cape,<br />
<strong>New</strong> <strong>Zealand</strong>, bottom fauna, 28 m (as 70 fathoms),<br />
Terra Nova stn 96 (BMNH 92 . .29. 49 [Tattersall’s<br />
92 specimen]).<br />
type LoCaLity: ‘Ostlich von Queensland’ (Studer<br />
884).<br />
desCription: Body 2.0 as long as greatest width, dorsal<br />
surfaces smooth, widest at pereonite 5, lateral margins<br />
weakly ovate or ovate. Rostral point folded ventrally<br />
and posteriorly. Eyes moderate, combined widths<br />
50–65% width <strong>of</strong> head, separated by about 32% width<br />
<strong>of</strong> head; each eye made up <strong>of</strong> ~9 transverse rows <strong>of</strong><br />
ommatidia, each row with ~7 ommatidia; eye colour<br />
black. Pereonite 1 and coxae 2–3 each with posteroventral<br />
angle right-angled. Coxae 5–7 with entire oblique<br />
carina; posterior margins concave, posterolateral angle<br />
acute (less than 45°). Pleon with pleonite largely concealed<br />
by pereonite 7; pleonite 4 with posterolateral<br />
margins extending to but not beyond posterior margin<br />
<strong>of</strong> pleonite 5; pleonite 5 with posterolateral angles<br />
overlapped by lateral margins <strong>of</strong> pleonite 4. Pleotelson<br />
0.7 times as long as anterior width, dorsal surface<br />
without longitudinal carina; lateral margins convex,<br />
serrate, posterior margin converging to caudomedial<br />
point, with 2 RS.<br />
Antennule peduncle extending to posterior <strong>of</strong> pereonite<br />
. Antenna flagellum extending to posterior <strong>of</strong><br />
pereonite 3 or pereonite 4.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, triangular,<br />
anterior margin with median point, posterior margin<br />
not abutting clypeus.<br />
Mandible molar process present, minute.<br />
Pereopod 1 basis 2. times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with<br />
0 RS ( long simple seta), superior distal margin with<br />
RS; merus inferior margin with RS, set as distal group,<br />
superior distal angle with 0 RS; carpus 0.8 as long as<br />
merus, inferior margin with RS (large); propodus<br />
* <strong>The</strong> date for this publication is usually cited as 883, but<br />
the library volume held at the ZMUC had a the date <strong>of</strong><br />
publication as 884.<br />
20<br />
.7 times as long as proximal width, inferior margin<br />
with 2 RS, propodal palm simple, without blade or<br />
process, dactylus smoothly curved, .2 as long as<br />
propodus. Pereopod 2 ischium inferior margin with 0<br />
RS, superior distal margin with 2 RS; merus inferior<br />
margin with 6 RS, set as two groups, superior distal<br />
margin with 2 acute RS; carpus similar in size to that<br />
<strong>of</strong> pereopod , inferodistal angle with RS. Pereopod 3<br />
similar to pereopod 2. Pereopods 5–7 inferior margins<br />
<strong>of</strong> ischium–carpus with short RS. Pereopod 6 similar to<br />
pereopod 7. Pereopod 7 basis 2.7 times as long as greatest<br />
width, inferior margins with 6 palmate setae; ischium<br />
0.4 as long as basis, inferior margin with 3 RS (set as ,<br />
2), superior distal angle with 3 RS, inferior distal angle<br />
with 3 RS; merus .0 as long as ischium, 2.2 times as<br />
long as wide, inferior margin with 2 RS (single cluster),<br />
superior distal angle with 0 RS, inferior distal angle<br />
with 6 RS; carpus . as long as ischium, 2.7 times as<br />
long as wide, inferior margin with 3 RS (set as , 2),<br />
superior distal angle with 9 RS, inferior distal angle<br />
with 7 RS; propodus .0 as long as ischium, 3.8 times<br />
as long as wide, inferior margin with 3 RS (set as , 2),<br />
superior distal angle with 2 slender setae (and RS),<br />
inferior distal angle with 3 RS.<br />
Pleopod 1 exopod .7 times as long as wide, distally<br />
broadly rounded, lateral margin straight, mesial margin<br />
strongly convex, with PMS from distal one-third,<br />
with ~50 PMS; endopod .6 times as long as wide,<br />
distally rounded, lateral margin convex, with PMS<br />
from distal half, mesial margin with PMS on distal<br />
margin only, endopod with ~24 PMS; peduncle .9<br />
times as wide as long, mesial margin with 6 coupling<br />
hooks. Exopods <strong>of</strong> pleopods 1–3 each with distolateral<br />
margin digitate; endopods <strong>of</strong> pleopods 3–5 each with<br />
distolateral point; pleopods 2–5 peduncle distolateral<br />
margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 3 RS,<br />
posterior lobe about three-quarters as long as endopod.<br />
Endopod apically not bifid, lateral margin weakly<br />
convex, without prominent excision, proximal lateral<br />
margin with 0 RS, distal lateral margin with 2 RS,<br />
mesial margin weakly convex, with 4 RS. Exopod not<br />
extending to end <strong>of</strong> endopod, 2.3 times as long as greatest<br />
width, apically not bifid; lateral margin convex,<br />
with 8 RS; mesial margin convex, with 4 RS.<br />
remarks: Aegiochus laevis can be identified by the relatively<br />
small eyes, triangular frontal lamina with free<br />
posterior margin, relatively prominent robust setae on<br />
the inferior margins <strong>of</strong> pereopods –3, and the pattern<br />
<strong>of</strong> robust setae on the uropodal rami and posterior margin<br />
<strong>of</strong> the pleotelson. A. laevis is further characterised<br />
by the distolateral margins <strong>of</strong> the pleopod exopods,<br />
particularly pleopods –3, being digitate.<br />
<strong>The</strong> <strong>New</strong> <strong>Zealand</strong> specimens agree entirely with the<br />
redescription <strong>of</strong> the holotype given by Bruce ( 983),
Figure 69. Aegiochus laevis (Studer, 883). NIWA 23763. A, dorsal view; B, lateral view; C, head, dorsal view; D, frons;<br />
E, pleonites, lateral margin; F, pleotelson, posterior margin; G, pleopod ; H, pleopod 2; I, uropod peduncle, distolateral<br />
angle.<br />
2
Figure 70. Aegiochus laevis (Studer, 883). NIWA 23763. A–C, pereopods , 2 and 7 respectively; D, uropod.<br />
with close correspondence <strong>of</strong> the frontal lamina, antennule,<br />
antenna, pereopods, pleopods and uropods. <strong>The</strong><br />
holotype was described as lacking robust setae on the<br />
posterior margin <strong>of</strong> the pleotelson, whereas one <strong>New</strong><br />
<strong>Zealand</strong> specimen has two robust setae; it is common<br />
for such setae to be missing from old specimens and<br />
this is not here regarded as being definitive in the holotype.<br />
<strong>The</strong> North Cape specimen (BMNH) has a ‘rubbed’<br />
pleotelson that lacks robust setae, though indentations<br />
suggest that it too may have had two robust setae. An<br />
abbreviated description is given here to allow ready<br />
identification.<br />
<strong>The</strong>re are four other <strong>New</strong> <strong>Zealand</strong> species with<br />
digitate pleopods — A. insomnis sp. nov., A. nohinohi<br />
22<br />
sp. nov., A. gordoni sp. nov. and A. coroo (Bruce, 983)<br />
(and also Aegiochus bertrandi sp. nov, which is within<br />
the northern reaches <strong>of</strong> the <strong>New</strong> <strong>Zealand</strong> chart area).<br />
Of these, A. coroo has large medially united eyes, and<br />
all except A. nohinohi have relatively weak or minute<br />
robust setae on pereopods –3. Aegiochus nohinohi has a<br />
more slender body shape, larger eyes, the robust setae<br />
on pereopods –3 are smaller and the posterior margin<br />
<strong>of</strong> the pleotelson lacks robust setae.<br />
prey: Not known.<br />
distribution: Northern <strong>New</strong> <strong>Zealand</strong>, northwards to<br />
Queensland, Australia; potentially widespread in the
southwestern or western Pacific; recorded depths <strong>of</strong><br />
26 to 2 6 metres.<br />
Aegiochus nohinohi sp. nov. (Figs 7 –74)<br />
materiaL examined: Holotype. ♀ (non-ovig. 9.0 mm),<br />
Rumble III Sea Mount, 35°44.5 –44.35’S, 78°30.20–<br />
29.75’E, 9 May 200 , 470–260 m, on scoria rubble, coll.<br />
RV Tangaroa (NIWA 24009).<br />
Paratypes. 4 ♂ (4.9, 5.0, 5.2, 6.7 mm), 4 ♀ (non-ovig.<br />
6.4, 6.9 [dissected], 8.6 [dissected], 9.0 mm), manca (4.4<br />
mm), same data as holotype (all at least slightly damaged;<br />
NIWA 240 0). ♀ (non-ovig. 7.0 mm), Rumble<br />
III Sea Mount, 35°44.38–44.35’S, 78°29.85–29.44’E, 9<br />
May 200 , 420–200 m, on scoria rubble, coll. RV Tangaroa<br />
(NIWA 240 ). ♀ (non-ovig. 7. mm), Rumble<br />
III Sea Mount, 35°44.40–44.7 ’S, 78°29.85–30.02’E, 9<br />
May 200 , 96–4 5 m, on scoria rubble, coll. RV Tangaroa<br />
(NIWA 240 2). ♀ (ovig. 7.5 mm), Rumble III Sea<br />
Mount, 35°44.28–43.98’S, 78°29.89–30.03’E, 20 May<br />
200 , 340–300 m, on scoria rubble, coll. RV Tangaroa<br />
(NIWA 240 3). ♂ (5.2 mm), manca (3.4 mm), Rumble<br />
III Sea Mount, 35°44.34–44.24’S, 78°29.74–29.53’E, 23<br />
May 200 , 200–500 m, on scoria rubble, coll. RV Tangaroa<br />
(NIWA 240 4). 2 ♀ (ovig. 0.2; non-ovig. 9.0 mm),<br />
Rumble V Sea Mount, 36°08.70–40’S, 78° 2.07– .8 ’E,<br />
24 May 200 , 40–690 m, on scoria rubble, coll. RV<br />
Tangaroa (NIWA 240 5).<br />
Non-type. ♀ (ovig. 6.5 mm, crushed), Rumble III Sea<br />
Mount, 35°44.49–44.52’S, 78°29.84–29.40’E, 9 May<br />
200 , 426–270 m, on scoria rubble, coll. RV Tangaroa<br />
(NIWA 240 6). 2 ♀ (ovig 7.7; non-ovig. 9.3 mm), coll.<br />
RV Kaharoa (NIWA 240 7). ♂ (6.4 mm), ♀ (ovig. 8.5,<br />
8.6, non-ovig. 6.0 mm), manca (3.8 mm), 43.0667°S,<br />
78.6500°E, September 963, 549 m (NIWA 240 8).<br />
♂ (6.2 mm), manca (5.0 mm), <strong>of</strong>f Three Kings Island,<br />
34° 3.0’S, 74° .5’E, 9 Feb 974, BS 396, 256 m, coll.<br />
RV Acheron (AK 4604).<br />
?♂ (6.9 mm), east <strong>of</strong> Bermagui, <strong>New</strong> South Wales,<br />
Australia, 36°25.2’S, 50° 8.5’E, 5 September 994, 220<br />
m, coll. Southern Surveyor (AM P74738).<br />
desCription: Body 2.8 times as long as greatest width,<br />
dorsal surfaces polished in appearance, widest at<br />
pereonite 6, lateral margins subparallel. Rostral point<br />
present, folded ventrally and posteriorly. Eyes large,<br />
not medially united (just under 50%), separated by<br />
about 6% width <strong>of</strong> head; each eye made up <strong>of</strong> ~7<br />
transverse rows <strong>of</strong> ommatidia, each row with ~9<br />
ommatidia; eye colour red. Pereonite 1 and coxae 2–3<br />
each with posteroventral angle rounded. Coxae 5–7<br />
with entire oblique carina; posterior margins convex,<br />
posterolateral angle blunt (more than 45°). Pleon with<br />
pleonite visible in dorsal view; pleonite 4 with posterolateral<br />
margins not extending to posterior margin<br />
<strong>of</strong> pleonite 5; pleonite 5 with posterolateral angles<br />
free, not overlapped by lateral margins <strong>of</strong> pleonite 4.<br />
Pleotelson 0.8 times as long as anterior width, dorsal<br />
surface without longitudinal carina; lateral margins<br />
convex, serrate (weakly), posterior margin converging<br />
to caudomedial point, with 0 RS.<br />
Antennule peduncle articles 3 and 4 .0 times as<br />
long as combined lengths <strong>of</strong> articles and 2, article 3<br />
2.9 times as long as wide; flagellum with 11 articles,<br />
extending to posterior <strong>of</strong> pereonite . Antenna peduncle<br />
article 4 2.2 times as long as wide, 0.8 times as long as<br />
combined lengths <strong>of</strong> articles –3, inferior margin with<br />
0 plumose setae, and 0 short simple setae (anterodistal<br />
angle with 4 long simple setae); article 5 .3 times as<br />
long as article 4, 2.8 times as long as wide, inferior<br />
margin with 2 pappose setae, anterodistal angle with<br />
cluster <strong>of</strong> 5 short simple setae (long SS, and 2 pappose<br />
setae); flagellum with 14 articles, extending to posterior<br />
<strong>of</strong> pereonite 3.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, posterior<br />
margin concave; anterior margin with median point,<br />
posterior margin not abutting clypeus.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 4 distolateral setae (2 serrate,<br />
2 simple), 3 with 3 setae. Maxillule with 6 terminal RS<br />
( large, 5 slender). Maxilla mesial lobe with 2 RS (<br />
biserrate); lateral lobe with 3 RS. Maxilliped endite with<br />
0 apical setae; palp article 2 with 2 RS; article 3 with 4<br />
recurved RS (all slender); article 4 with 4 hooked RS<br />
(and small slender; unevenly spaced with proximal<br />
3 distal); article 5 articulating with article 4, with 4<br />
hooked/straight RS (2 large, one <strong>of</strong> which is serrate,<br />
and 2 small slender).<br />
Pereopod 1 basis 2.5 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin<br />
with 0 RS, superior distal margin with RS; merus<br />
inferior margin with 0 RS ( long and short simple<br />
setae), superior distal angle with 0 RS ( simple seta);<br />
carpus 0.6 as long as merus; inferior margin with RS<br />
(large); propodus .9 times as long as proximal width,<br />
inferior margin with 2 RS (set as and distally), propodal<br />
palm simple, without blade or process, dactylus<br />
smoothly curved, .2 as long as propodus. Pereopod<br />
2 ischium inferior margin with 0 RS, superior distal<br />
margin with RS; merus inferior margin with 3 RS<br />
(set as and 2 plus distal large simple seta), set as in<br />
two groups, superior distal margin with 0 acute RS (<br />
simple seta); carpus similar in size to that <strong>of</strong> pereopod<br />
or longer than that <strong>of</strong> pereopod , with inferodistal<br />
lobe, inferodistal angle with RS. Pereopod 3 similar<br />
to pereopod 2. Pereopods 5–7 superior margins <strong>of</strong><br />
ischium–carpus without setae. Pereopod 6 similar<br />
to pereopod 7 (with larger RS). Pereopod 7 basis 2.6<br />
times as long as greatest width, inferior margin with<br />
4 palmate setae; ischium 0.5 as long as basis, inferior<br />
margin with 3 RS (set singly), superior distal angle with<br />
23
Figure 71. Aegiochus nohinohi sp. nov. G, H, female paratype (8.6 mm, NIWA 240 0), remainder holotype. A, dorsal<br />
view; B, lateral view; C, head; D, frons, ventral view; E, frons, anterior view; F, penial openings; G, antenna peduncle;<br />
H, antennule peduncle.<br />
2 RS, inferior distal angle with 3 RS; merus .3 as long<br />
as ischium, 2.4 times as long as wide, inferior margin<br />
with 2 RS (single pair), superior distal angle with 7<br />
RS, inferior distal angle with 4 RS; carpus .2 as long<br />
as ischium, 3.0 as long as wide, inferior margin with 3<br />
RS (set as and 2), superior distal angle with 6 RS (2<br />
biserrate), inferior distal angle with 6 RS; propodus .<br />
24<br />
as long as ischium, 4.3 times as long as wide, inferior<br />
margin with 2 RS (set singly), superior distal angle with<br />
slender seta (2 small simple and plumose setae),<br />
inferior distal angle with 3 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7; penial<br />
openings separated by 6% <strong>of</strong> sternal width.
Figure 72. Aegiochus nohinohi sp. nov. Female paratype (8.6 mm, NIWA 240 0). A, mandible; B, mandible palp article 3;<br />
C, maxillule apex; D, maxilla ; E, maxilla apex; F, maxilliped; G, maxilliped articles 2–5; H, uropod exopod apex; I, uropod<br />
endopod apex.<br />
25
Figure 73. Aegiochus nohinohi sp. nov. Female paratype (6.9 mm, NIWA 240 0). A–D, pereopods , 2, 6 and 7 respectively;<br />
E, dactylus unguis, pereopod 6; E, dactylus, pereopod 7.<br />
Pleopod 1 exopod .5 times as long as wide, lateral<br />
margin weakly convex, mesial margin strongly<br />
convex, with ~35 PMS; endopod .6 times as long as<br />
wide, lateral margin convex, mesial margin straight,<br />
endopod with ~ 7 PMS; peduncle .8 times as wide as<br />
long, peduncle mesial margin with 5 coupling hooks.<br />
Pleopod 2 exopod with ~36 PMS, endopod with ~ 8<br />
PMS; appendix masculina .2 times as long as endopod,<br />
distally acute. Pleopod 3 exopod with ~49 PMS, endo-<br />
26<br />
pod with ~ PMS. Pleopod 4 exopod with ~40 PMS,<br />
endopod with ~ 0 PMS. Pleopod 5 exopod with ~37<br />
PMS. Exopods <strong>of</strong> pleopods –3 each with distolateral<br />
margin digitate; endopods <strong>of</strong> pleopods 3–5 each with<br />
distolateral point; pleopods 2–4 peduncle distolateral<br />
margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 3 RS,<br />
posterior lobe about one-half as long as endopod.<br />
Endopod apically deeply bifid, mesial process promi-
Figure 74. Aegiochus nohinohi sp. nov. B holotype, remainder female paratype ([dissected] NIWA 240 0). A–E, pleopods –5<br />
respectively; E, uropod exopod, ventral view; F, uropod.<br />
nent, lateral margin straight, without prominent excision,<br />
lateral margin with RS, mesial margin weakly<br />
convex, with 2 RS (small). Exopod extending to end <strong>of</strong><br />
endopod, 3.6 times as long as greatest width, apically<br />
deeply bifid, mesial process prominent; lateral margin<br />
weakly convex, lateral margin with 6 RS; mesial margin<br />
weakly convex, with 2 RS (small).<br />
femaLe: Similar to males, ovigerous females proportionally<br />
wider (2. times as long as greatest width).<br />
size: Males 4.9–6.7 mm (mean 5.4 mm, n = 5) ; ovigerous<br />
females 6.5– 0.2 mm, (mean 8.9 mm, n = 3), nonovigerous<br />
females 6.4–9.0 mm, (mean 7.7 mm, n = 7);<br />
one manca measured 4.4 mm, and eggs measured<br />
.0– .2 mm.<br />
27
Variation: Most specimens were damaged with uropods<br />
crushed or broken. Robust setae: (n = 3) pleotelson<br />
always without RS. Uropod exopod mesial (n = 9) margin<br />
0 ( 6%), 2 (42%) or 3 (36%), lateral margin 6 (56%)<br />
or 7 (44%); uropod endopod mesial margin (n = 9) with<br />
0 (42%), (37%) or 2 (2 %), lateral margin with only<br />
large RS distal to the pappose seta, twice without RS,<br />
possibly owing to damage. <strong>The</strong>re is no discernable difference<br />
between males and females. <strong>The</strong> robust setae<br />
are small and fine.<br />
<strong>The</strong> small size <strong>of</strong> the species precluded making accurate<br />
direct counts <strong>of</strong> the robust setae on pereopods<br />
and 2 although <strong>of</strong> the specimens examined the pattern,<br />
number and size appears consistent.<br />
remarks: Aegiochus nohinohi sp. nov., at an average<br />
length <strong>of</strong> 5.4 mm for males, about 8 mm for females,<br />
is the smallest species <strong>of</strong> the genus in <strong>New</strong> <strong>Zealand</strong><br />
waters. Aegiochus gordoni sp. nov. is similar in many aspects,<br />
but is distinctly larger (on average 43–66% longer<br />
BL than A. nohinohi). <strong>The</strong>re are numerous morphological<br />
differences between these two species, including<br />
the eyes being smaller and more widely separated in<br />
A. nohinohi, uropod endopod mesial margin with fewer<br />
( or 2) and smaller RS (usually 3 in A. gordoni); pereopods<br />
–3 with more and much larger RS on the merus,<br />
carpus and propodus than does A. gordoni; pereopods<br />
–3 with a relatively shorter dactylus ( .2 as long as<br />
propodus for A. nohinohi v. .4 as long for A. gordoni),<br />
and the pereopods are in general more robust. <strong>The</strong>se<br />
two species are distinguished from all other species in<br />
<strong>New</strong> <strong>Zealand</strong> by their small size, separate eyes, serrate<br />
pleopod exopods and weakly serrate shield-shaped<br />
pleotelson that totally lacks robust setae.<br />
Aegiochus perulis (Menzies & George, 972), known<br />
only from the female holotype from the East Pacific <strong>of</strong>f<br />
Chile, is similar in appearance and size (9.2 mm), with<br />
relatively small and widely separated eyes, a similar<br />
frontal lamina and similar uropod and pleotelson<br />
shape. It differs in the body shape being more ovate<br />
and in having 2 robust setae on the posterior margin<br />
<strong>of</strong> the pleotelson. <strong>The</strong> brief description (Menzies &<br />
George 972) precludes further comparisons.<br />
A single specimen from <strong>of</strong>f Bermagui, <strong>New</strong> South<br />
Wales (AM P74738) is provisionally included under<br />
this name. It agrees well with the <strong>New</strong> <strong>Zealand</strong> material,<br />
differing only in having the antennule and antenna<br />
proportionally longer (extending to the posterior <strong>of</strong><br />
pereonites 2 and 3 respectively).<br />
prey: Not known.<br />
distribution: Recorded from the Chatham Rise to the<br />
east <strong>of</strong> South Island, the Brothers Sea Mounts to the<br />
northeast <strong>of</strong> North Island and <strong>of</strong>f Three Kings Island;<br />
possibly also southeastern Australia.<br />
28<br />
At depths from 360 to 40 metres (the minimum<br />
depth was recorded from a haul that ran from 360 to<br />
755 metres, all other records are from 698 metres or<br />
greater).<br />
etymoLogy: Nohinohi is a Mäori word that means small,<br />
this species being one <strong>of</strong> the smallest <strong>of</strong> those that occur<br />
<strong>New</strong> <strong>Zealand</strong> waters (noun in apposition).<br />
Aegiochus piihuka sp. nov. (Figs 75–78)<br />
materiaL examined: Holotype, ♂ (22 mm), northeast <strong>of</strong><br />
Hawkes Bay, North Island, <strong>New</strong> <strong>Zealand</strong>, 37°0 .39’S,<br />
76°43.09’E, 20 Jul 998, 972– 207 m, Z9 8 , from hexactinellid<br />
sponge (NIWA 23775).<br />
Paratypes. <strong>New</strong> <strong>Zealand</strong>: ♀ (28 mm, dissected),<br />
Z9 8 , same data as holotype (NIWA 23776). ♂ ( 8.5<br />
mm), <strong>of</strong>f Hawkes Bay, 40°0 .5’S, 78°03.3’E, 28 August<br />
986, from trawled sponge, 935 m (NMNZ Cr.5953). ♂<br />
( 7.0 mm), <strong>of</strong>f White Island, 37°23.7’S, 77°39.5–36.6’E,<br />
23 November 98 , 075– 00 m, FV USSR Kalinovo<br />
(NMNZ Cr. 20 9).<br />
Non-type material: Australia, Nsw: 5 ♂ ( 7.0, 7.5,<br />
8.0, 9.0, 9.5 mm), 6 ♀ (ovig. 27, 28, 29, 3 ; non-ovig.<br />
26, 30 mm) east <strong>of</strong> Cape Hawke, NSW, 32°06.02’S,<br />
53°08.09’E, 2 February 983, from sponge, 940–980<br />
m, coll. FRV Kapala (AM P347 3). 2 ♂ (20, 2 mm),<br />
east <strong>of</strong> Broken Bay, NSW, 33°32–39’S, 52°09– 2’E, 23<br />
August 983, 955 m, coll. FRV Kapala (AM P34709). 3 ♂<br />
(20, 2 , 28 mm), east <strong>of</strong> Broken Bay, NSW, 33°39–37’S,<br />
52°06–07’E, 6 December 983, 006 m, coll. FRV<br />
Kapala (AM P34705). ♀ (ovig. 30 mm), many mancas<br />
(9.5– 0.5 mm), east <strong>of</strong> Broken Bay, NSW, 33°39–37’S,<br />
52°06–07’E, 6 December 979, 006 m, coll. FRV Kapala<br />
(AM P34706). Queensland: 3♂ ( 7.0, 8.5, 9.0 mm),<br />
2♀ (ovig. 23, 24 mm), Coral Sea, 7°0 .8’S, 5 °20. ’E,<br />
6 December 985, 800 m, coll. P.J.F. Davie on RV Soela<br />
(QM W 8829). <strong>New</strong> caledonia: ♀ (non-ovig. 34 mm),<br />
2 ° 5.0 ’S, 57°5 .33’E, 4 October 986, 970 m, MU-<br />
SORSTOM V (MNHN unreg). taiwan: ♂ (20 mm),<br />
22°20.98’N, 20°6.73’E, 2 November 200 , 690–700 m,<br />
Otter Trawl (Le Drézén type JUNEAUX), stn CD 32, coll.<br />
RV Ocean Researcher (MTQ W 3680).<br />
Unmeasured: Eastern Australia. 3♀ (ovig.), east <strong>of</strong><br />
Cape Hawke, NSW, 32°09’S, 53°09’E, 8 July 984,<br />
052– 25 m, coll. FRV Kapala (AM P43976). ♂, east<br />
<strong>of</strong> Long Reef Point, NSW, 32°45–4 ’S, 52°00–03’E,<br />
October 984, 5– 005 m, coll. FRV Kapala (AM<br />
P37504). ♂, east <strong>of</strong> Broken Bay, NSW, 33°32–29’S,<br />
52°09– 2’E, 23 August 983, 955 m, coll. FRV Kapala<br />
(AM P34708). ♂, east <strong>of</strong> Broken Bay, NSW, 33°32–24’S,<br />
52°09– 2’E, 23 August 983, 955 m, coll. FRV Kapala<br />
(AM P34707, photographed). 2♀ (ovig), southeast <strong>of</strong><br />
<strong>New</strong>castle, NSW, 33°05–04’S, 52°33–36’E, 5 May<br />
988, from glass sponge, 900–950 m, coll. FRV Kapala<br />
(AM P43980). 2♀ (ovig), east <strong>of</strong> Broken Bay, NSW,
33°47–44’S, 5 °59’– 52°0 ’E, from sponge, 987– 005<br />
m, coll. FRV Kapala (AM P43979). 2♀ (non-ovig), northeast<br />
<strong>of</strong> Tuncurry, NSW, 32°08’S, 53°09’E, 989, with<br />
bits <strong>of</strong> hexactinellid sponge attached, 034– 079 m (AM<br />
P4398 ). 94 specimens, <strong>of</strong>f Tuncurry, NSW, 32°09–05’S,<br />
53°09’E, 2 June 988, ‘from tall sponge’, 066– 00<br />
m, coll. FRV Kapala (AM P43973).<br />
Additional material: vanuatu: MUSORSTOM, 8,<br />
coll. B. Richer de Forges: 3, 5°57.30’S, 67°27.73’E, 5<br />
October 994, stn. CP 080, 799–850 m (MNMN Is.5868);<br />
2, 8°57.70’S, 68°54.40’E, 29 September 994, stn.<br />
CP 037, 058– 086 m, (MNMN Is.5869); 3, 8°0 .00’S,<br />
68°48.20’E, 20 September 994, stn. CP 036, 920–950<br />
m (MNMN Is.5870); 3, 5°52.62’S, 67°20.36’E, 22<br />
September 994, stn. CP 082, 492–520 m, in sponge<br />
(MNMN Is.5871); 3, 6°00.73’S, 66°39.94’E, 0 October<br />
994, stn. CP 29, 0 4– 050 m, (MNMN Is.5872). <strong>New</strong><br />
caledonia: , 2 °42.8 ’S, 66°4 .95’E, 20 March 994,<br />
HALIPRO stn. CP858, 000– 20 m, coll. B. Richer<br />
de Forges (MNHN Is.5873). 2, 25° 7’S, 70°24’E,<br />
November 996, HALIPRO 2, stn. BT25, 000– 348 m,<br />
from hexactinellid sponge, coll. B. Richer de Forges<br />
(MNHN Is.5874). 3, 2 °4 .80’S, 66°40. 0’E, March<br />
993, BATHUS stn. CP65 , 080– 8 m, coll. B.<br />
Richer de Forges (MNHN Is.5875). , 24°44.24’S,<br />
170°0.01’E, 24 Feb 1993, BATHUS 3 St. DW776, 770–830<br />
m, coll. B. Richer de Forges (MNHN Is.5877). 3, 24°43.49’S,<br />
170°07.07’E, 24 November 1993, BATHUS 3 stn. DW778,<br />
750–760 m, coll. B. Richer de Forges (MNHN Is.5878).<br />
Also examined: Comparative material <strong>of</strong> Aegiochus<br />
plebeia, see Appendix 2.<br />
desCription: Body 2.5 times as long as greatest width,<br />
dorsal surfaces sparsely punctate, widest at pereonite<br />
5, lateral margins weakly ovate. Rostral point folded<br />
ventrally and posteriorly. Eyes large, not medially<br />
united, separated by about 8– 0% width <strong>of</strong> head; each<br />
eye made up <strong>of</strong> ~8–9 transverse rows <strong>of</strong> ommatidia,<br />
each row with ~ 8 ommatidia; eye colour dark brown.<br />
Pereonite 1 and coxae 2–3 each with posteroventral angle<br />
with small distinct produced point. Coxae 5–7 with<br />
entire oblique carina; posterior margins straight and<br />
sinuate (coxa 6 sinuate, 7 straight), posterolateral angle<br />
acute (less than 45°). Pleon with pleonite visible in<br />
dorsal view; pleonite 4 with posterolateral margins extending<br />
to but not beyond posterior margin <strong>of</strong> pleonite<br />
5; pleonite 5 with posterolateral angles overlapped by<br />
lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.8 times as long<br />
as anterior width, dorsal surface without longitudinal<br />
carina; lateral margins convex, notched, posterior margin<br />
converging to caudomedial point, with 2 RS.<br />
Antennule peduncle articles 3 and 4 . times as<br />
long as combined lengths <strong>of</strong> articles and 2, article 3<br />
3.3 times as long as wide (narrowing distally); flagellum<br />
with 9 articles, extending to anterior <strong>of</strong> pereonite<br />
. Antenna peduncle article 4 2.0 as long as wide, 0.8<br />
times as long as combined lengths <strong>of</strong> articles –3,<br />
inferior margin 0 plumose setae, and 4 short simple<br />
setae; article 5 .0 times as long as article 4, 2.2 times<br />
as long as wide, inferior margin with 2 palmate setae,<br />
anterodistal angle with cluster <strong>of</strong> 5 short simple setae;<br />
flagellum with 21 articles, extending to posterior <strong>of</strong><br />
pereonite 2.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, posteriorly<br />
rounded, anterior margin acute, forming median angle,<br />
posterior margin not abutting clypeus.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 8 distolateral setae (biserrate),<br />
palp article 3 with 25 setae (simple; distal 3 markedly<br />
longer than remainder). Maxillule with 7 terminal RS<br />
(three largest being weakly hooked). Maxilla mesial<br />
lobe with 3 RS (straight); lateral lobe with 4 RS. Maxilliped<br />
endite with 2 apical setae (minute); palp article<br />
2 with 2 RS; article 3 with 5 recurved RS (straight or<br />
weakly recurved); article 4 with 4 hooked RS; article 5<br />
with 6 RS (as 2 stout and 2 pairs <strong>of</strong> slender).<br />
Pereopod 1 basis 3.0 as long as greatest width; ischium<br />
0.5 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with RS (and one short<br />
simple seta); merus inferior margin with RS (minute),<br />
superior distal angle with 0 RS ( simple seta); carpus<br />
0.7 as long as merus, inferior margin with RS (set on<br />
weak lobe); propodus 2.6 times as long as proximal<br />
width, inferior margin with RS, propodal palm with<br />
large distal lobe (with distal margin at right angles to<br />
axis <strong>of</strong> propodus), dactylus smoothly curved, .5 as<br />
long as propodus. Pereopod 2 ischium inferior margin<br />
with 0 RS, superior distal margin with RS (and<br />
simple seta); merus inferior margin with 4 RS (concave,<br />
distally lobed), set as single row or two groups,<br />
superior distal margin with acute RS; carpus longer<br />
than that <strong>of</strong> pereopod , with inferodistal lobe, inferodistal<br />
angle with 2 RS. Pereopod 3 similar to pereopod<br />
2. Pereopods 5–7 inferior margins <strong>of</strong> ischium–carpus<br />
with short RS. Pereopod 6 similar to pereopod 7 (but<br />
longer). Pereopod 7 basis 3.2 times as long as greatest<br />
width, inferior margins with 2 palmate setae; ischium<br />
0.4 as long as basis, inferior margin with 3 RS (set as ,<br />
and ), superior distal angle with 3 RS, inferior distal<br />
angle with 3 RS; merus .3 as long as ischium, 2.6 times<br />
as long as wide, inferior margin with 5 RS (set as 4<br />
clusters <strong>of</strong> , 2, and ), superior distal angle with 0<br />
RS, inferior distal angle with 5 RS; carpus 0.9 as long<br />
as ischium, 2.9 times as long as wide, inferior margin<br />
with 4 RS (set as , 2 and submarginal), superior<br />
distal angle with 9 RS, inferior distal angle with 6 RS;<br />
propodus .0 as long as ischium, 4. times as long as<br />
wide, inferior margin with 6 RS (set as 3 loose clusters<br />
<strong>of</strong> 2 each), superior distal angle with 3 slender setae,<br />
inferior distal angle with 3 RS.<br />
Penes low tubercles; penial openings separated by<br />
0% <strong>of</strong> sternal width.<br />
29
Figure 75. Aegiochus piihuka sp. nov. A–E, holotype; F, ovigerous female; remainder female NIWA 23776. A, dorsal view;<br />
B, lateral view; C, head; D, frons; E, penial openings; F, dorsal view; G, antenna; H, antennule.<br />
Pleopod 1 exopod 2.0 as long as wide, distally narrowly<br />
rounded, mesial margin weakly oblique, lateral<br />
margin distally concave, mesial margin strongly convex,<br />
with PMS on distal two-thirds; endopod 2.0 as<br />
long as wide, distally rounded, lateral margin weakly<br />
concave, with PMS on distal margin only, mesial margin<br />
with PMS on distal half; peduncle .4 times as wide<br />
as long, mesial margin with 6 coupling hooks. Pleopod<br />
2 appendix masculina basally swollen (weakly), .0 as<br />
long as endopod, distally bluntly rounded. Exopods<br />
<strong>of</strong> pleopods –3 each with distolateral margin not digitate;<br />
endopods <strong>of</strong> pleopods 3–5 each with distolateral<br />
30<br />
point; pleopods 2–4 peduncle distolateral margin with<br />
prominent acute RS.<br />
Uropod peduncle ventrolateral margin with RS,<br />
posterior lobe about two-thirds as long as endopod.<br />
Endopod apically sub-bifid, mesial process prominent,<br />
lateral margin straight, without prominent excision,<br />
proximal lateral margin with RS, distal lateral margin<br />
with 3 RS, mesial margin weakly convex, with 7<br />
RS. Exopod not extending to end <strong>of</strong> endopod, 2.4 times<br />
as long as greatest width, apically sub-bifid, mesial<br />
process prominent; lateral margin convex, with 3 RS;<br />
mesial margin convex, with 5–7 RS.
Figure 76. Aegiochus piihuka sp. nov. A, B, E, H I, male NMNZ Cr.5953, remainder female paratype NIWA 23776; A, mandible;<br />
B, mandible palp article 3; C, maxillule; D, maxillule apex; E, maxillule apex, oblique view; F, maxilla; G, maxilla apex; H,<br />
maxilliped; I, maxilliped articles 2–5; J, female maxilliped; K, female maxilliped articles 2–5.<br />
3
Figure 77. Aegiochus piihuka sp. nov. Holotype. A–D, pereopods , 2, 6 and 7 respectively; E, dactylus unguis, pereopod 6;<br />
F, dactylus, pereopod 7; G, distolateral margin, pereopod 6 merus; H, distolateral margin, pereopod 6 carpus.<br />
32
Figure 78. Aegiochus piihuka sp. nov. F, female paratype NIWA 23776; remainder holotype. A–E pleopods –5 respectively;<br />
F, uropod exopod, ventral view; G, uropod; H, uropod rami, apices; I, pleotelson, dorsal view.<br />
femaLe: Body 2.0 as long as greatest width, lateral<br />
margins ovate; otherwise similar to male.<br />
size: Males 7–28 mm (mean = 20 mm, n = 7); ovigerous<br />
females 23–3 mm (mean = 27 mm, n = 7); nonovigerous<br />
females 26–34 mm (mean = 28 mm, n = 4).<br />
Variation: Robust setae: (n = 4) pleotelson RS 6+6 (50%),<br />
5+6 (36%) and 5+5 ( 4%). Uropod exopod mesial margin<br />
5 (25%), 6 (50%), 7 (2 %) and 8 once, lateral margin<br />
2 (39%) or 3 (57%); uropod endopod mesial margin<br />
varied from 5 to 0 RS with 7 (26%), 8 (34%) or 9 (34%)<br />
the most frequent, lateral margin with +2 ( 0%), +3<br />
33
(64%). 2+3 ( 4%), and 2+2 (7%). <strong>The</strong>re is no discernable<br />
difference between males and females, nor does the<br />
number <strong>of</strong> RS increase with the size <strong>of</strong> the specimen.<br />
<strong>The</strong> most distal RS on the uropodal endopod lateral<br />
margin (see Fig. 78H) is small and is usually lost with<br />
damage to the apex—this RS has not been included in<br />
the numbers given here. <strong>The</strong> most proximal RS on the<br />
uropodal exopod lateral margin is minute and could<br />
<strong>of</strong>ten be missed, and therefore the maximum number<br />
<strong>of</strong> RS could be one more than here stated.<br />
remarks: <strong>The</strong> morphology <strong>of</strong> pereopods –3 is unique.<br />
No other species has a similarly shaped distal lobe on<br />
the propodus, nor the inferodistal lobes <strong>of</strong> the merus<br />
and carpus <strong>of</strong> pereopods 2 and 3. This character is<br />
consistent for males, females and mancas, and serves<br />
to identify the species. <strong>The</strong> specimen from Taiwan<br />
agrees entirely with material from the southwestern<br />
Pacific, notably in the details <strong>of</strong> the propodus <strong>of</strong> pereopods<br />
–3, eye size, coxal shape, shape <strong>of</strong> the frontal<br />
lamina and the counts for the marginal robust setae<br />
on the pleotelson and uropods. <strong>The</strong> distribution <strong>of</strong><br />
this species from <strong>New</strong> <strong>Zealand</strong> and Australia to Japan<br />
suggests that some records <strong>of</strong> Aegiochus plebeia may be<br />
<strong>of</strong> this species.<br />
Aegiochus plebeia (Hansen, 897), from the North<br />
and East Pacific (see Brusca 983), is perhaps the most<br />
similar species. Compared to Aegiochus piihuka sp. nov.<br />
A. plebeia (Appendix 2, Figs 43, 44) has a weak distal<br />
lobe provided with a prominent and large robust seta<br />
on the propodus <strong>of</strong> pereopods –3; has larger eyes,<br />
each with a wider mesial margin; the coxae are more<br />
truncate, scarcely extending posteriorly; and the lateral<br />
margins <strong>of</strong> the pleotelson are more strongly convex<br />
with 8–10 robust setae. Brusca’s (1983) figures and<br />
description <strong>of</strong> pereopod are not in mutual agreement—being<br />
described as having a propodal plate but<br />
figured without such. Brusca’s figures agree well with<br />
those <strong>of</strong> Hansen ( 897) but there is the possibility that<br />
more than one species is being identified under the name<br />
<strong>of</strong> A. plebeia (note—this epithet is frequently misspelled,<br />
e.g. plebeja—Brandt & Poore 2003; Gurjanova 936;<br />
Nierstrasz 93 ; plebia—Wetzer 990). Other similar<br />
species are Aegiochus symmetrica (Richardson, 905b)<br />
(northwestern Pacific) and Aegiochus ventrosa (Sars,<br />
859) (North Atlantic), but both <strong>of</strong> these species lack the<br />
propodal lobe on the anterior pereopods. See Appendix<br />
2 for supplementary description <strong>of</strong> Aegiochus plebeia.<br />
prey: Not known.<br />
distribution: Widespread in the western Pacific. Off<br />
northeastern North Island, <strong>New</strong> <strong>Zealand</strong>; also eastern<br />
Australia, <strong>New</strong> Caledonia, Vanuatu and Taiwan,<br />
frequently recorded from ‘glass sponges’ (Hexactinellidae).<br />
Most records are at depths from 700 to 25<br />
34<br />
metres in the southwestern Pacific, with single record<br />
<strong>of</strong> 492–520 metres <strong>of</strong>f <strong>New</strong> Caledonia and 690–700<br />
metres <strong>of</strong>f Taiwan. Frequency <strong>of</strong> collection suggests<br />
that this widespread species is common in the southwestern<br />
Pacific<br />
etymoLogy: <strong>The</strong> epithet, piihuka, a Mäori word meaning<br />
hook, alluding to the hooked anterior legs.<br />
Aegiochus pushkini (Kussakin & Vasina, 982),<br />
comb. nov. (Figs 79–83)<br />
Aega pushkini Kussakin & Vasina, 1982: 265, figs 7, 8.– Kensley,<br />
200 : 227.<br />
materiaL examined: Holotype, ♀ (non-ovig. 15.3 mm),<br />
Ob’ Bank, southern Indian Ocean, Stn 2634/4 2 (inner<br />
label states 659/4 2). 52° 7.8’S, 4 °4 .9’E, 4 0 m, coll.<br />
Skif (ZIAS RAN No /7 625).<br />
<strong>New</strong> <strong>Zealand</strong>: ♂ ( 6.5 mm [dissected]), southwest<br />
<strong>of</strong> <strong>New</strong> <strong>Zealand</strong>, 53.9 67°S, 58.9 67°E, 25 November<br />
1961, 366 m, stn C734 (NIWA 24008). ♀ (non-ovig. 17.0<br />
mm), manca (crushed), also C734 (NMNZ Cr. 2003).<br />
south Atlantic: ♀ (ovig. 25 mm, non-ovig. 17.5, 16.0<br />
mm), <strong>of</strong>f Bouvet Island, 54°22.49–54’S, 03° 7.58–2 ’E,<br />
25 November 2003, 34– 22 m, coll. ANT2 /2 BENDEx<br />
(ZMH K-4 228).<br />
desCription (<strong>of</strong> new zeaLand speCimen): Body 2.2 times<br />
as long as greatest width, dorsal surfaces smooth and<br />
sparsely punctate, widest at pereonites 5 and 6, lateral<br />
margins weakly ovate. Rostral point folded ventrally<br />
and posteriorly. Eyes large, not medially united, separated<br />
by about 2% width <strong>of</strong> head; each eye made up <strong>of</strong><br />
~ 7 transverse rows <strong>of</strong> ocelli, each row with ~ 0 ocelli;<br />
eye colour pale brown. Pereonite 1 and coxae 2–3 each<br />
with posteroventral angle right-angled. Coxae 5–7 with<br />
entire oblique carina; posterior margins straight posterolateral<br />
angle blunt (more than 45°). Pleon with pleonite<br />
visible in dorsal view; pleonite 4 with posterolateral<br />
margins extending to but not beyond posterior margin<br />
<strong>of</strong> pleonite 5; pleonite 5 with posterolateral angles free,<br />
not overlapped by lateral margins <strong>of</strong> pleonite 4. Pleotelson<br />
0.7 times as long as anterior width, dorsal surface<br />
without longitudinal carina; lateral margins convex,<br />
serrate, posterior margin converging to caudomedial<br />
point, with 0 RS.<br />
Antennule peduncle article 3 and 4 .0 times as long<br />
as combined lengths <strong>of</strong> articles and 2, article 3 3.3<br />
times as long as wide; flagellum with 12 articles (articles<br />
2– with distal margin widest), extending to anterior<br />
<strong>of</strong> pereonite . Antenna peduncle article 4 2. times as<br />
long as wide, inferior margin with plumose seta (?);<br />
article 5 0.9 times as long as article 4, 2.6 times as long as<br />
wide (widest distally), inferior margin with pappose<br />
seta (?); extending to middle <strong>of</strong> pereonite 2.
Figure 79. Aegiochus pushkini (Kussakin & Vasina, 982). Holotype. A, dorsal view; B, lateral view (coxa <strong>of</strong> pereonite 3 damaged);<br />
C, head; D, frons, ventral view; E, pleotelson posterior margin; F, pereopods 3– , left to right, in situ; G, uropod, in situ.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, posterior<br />
margin concave, anterior margin with median point,<br />
posterior margin not abutting clypeus.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 2 distolateral setae (all but<br />
biserrate), palp article 3 with 22 setae. Maxillule with 7<br />
terminal RS ( large, moderate and 5 slender). Maxilla<br />
mesial lobe with 3 RS ( straight, 2 biserrate); lateral<br />
35
Figure 80. Aegiochus pushkini (Kussakin & Vasina, 982). Male, NIWA 24008. A, dorsal view; B, lateral view; C, head; D, frons,<br />
ventral view; E, penial openings; F, coxae 2–4, right side; G, antenna peduncle; H, antennule peduncle.<br />
lobe with 3 RS. Maxilliped endite with apical seta; palp<br />
article 2 with 2 RS (straight); article 3 with 2 recurved<br />
RS (and large serrate and straight setae); article 4<br />
with 4 hooked RS; article 5 with 4 RS (long serrate).<br />
Pereopod 1 basis 2.8 times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with RS; merus inferior<br />
margin with 2 RS, set as two groups, superior distal<br />
angle with 0 RS ( slender seta); carpus .2 as long as<br />
merus; inferior margin with RS; propodus .7 times<br />
36<br />
as long as proximal width, inferior margin with RS<br />
(distal), propodal palm simple, without blade or process,<br />
dactylus smoothly curved, .2 as long as propodus.<br />
Pereopod 2 ischium inferior margin with 0 RS, superior<br />
distal margin with RS; merus inferior margin with<br />
3 RS (set as +2, proximal seta minute), set as two<br />
groups, superior distal margin with 0 acute RS (3 slender<br />
setae); carpus similar in size to that <strong>of</strong> pereopod ,<br />
inferodistal angle with RS (large). Pereopod 3 similar<br />
to pereopod 2 (but with single RS on propodal palm).
Figure 81. Aegiochus pushkini (Kussakin & Vasina, 982). Male, NIWA 24008. A, mandible; B, mandible palp article 3;<br />
C, maxillule; D, maxillule apex; E, maxilla; F, maxilla apex; G, maxilliped; H, maxilliped articles 2–5.<br />
37
Figure 82. Aegiochus pushkini (Kussakin & Vasina, 982). Male, NIWA 24008. A–E, pereopods –3, 6, and 7 respectively.<br />
38
Figure 83. Aegiochus pushkini (Kussakin & Vasina, 982). Male, NIWA 24008. A–D, pleopods –3, 5 respectively;<br />
E, uropod, in situ.<br />
Pereopod 6 similar to pereopod 7. Pereopod 7 basis 3.3<br />
times as long as greatest width, inferior margins with<br />
5 palmate setae; ischium .4 as long as basis, inferior<br />
margin with 3 RS (set as and 2), superior distal angle<br />
with 5 RS, inferior distal angle with 3 RS; merus .4<br />
as long as ischium, 2.8 times as long as wide, inferior<br />
margin with 3 RS (set as and 2), superior distal angle<br />
with 0 RS, inferior distal angle with 6 RS; carpus .3<br />
as long as ischium, 3.2 times as long as wide, inferior<br />
margin with 3 RS (set as and 2), superior distal angle<br />
with 9 RS, inferior distal angle with 5 RS; propodus .<br />
as long as ischium, 3.6 times as long as wide, inferior<br />
margin with 6 RS (set as 2, 2 and 2), superior distal<br />
angle with 2 slender setae (and robust seta), inferior<br />
distal angle with 3 RS.<br />
Penes low tubercles; penial openings separated by<br />
2% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod .7 times as long as wide, distally<br />
broadly rounded, lateral margin straight, mesial margin<br />
strongly convex, with PMS from distal two-thirds,<br />
with ~60 PMS; endopod 2.2 times as long as wide,<br />
distally rounded, lateral margin weakly concave, with<br />
PMS from distal one-third, mesial margin with PMS<br />
from distal one-third, endopod with ~28 PMS; pedun-<br />
39
cle 2.0 as wide as long, mesial margin with 7 coupling<br />
hooks. Pleopod 2 exopod with ~75 PMS, endopod with<br />
~50 PMS (24 short PMS on mesial margin); appendix<br />
masculina basally swollen, 0.96 times as long as endopod,<br />
distally acute. Pleopod 3 exopod with ~78 PMS, endopod<br />
with ~ 8 PMS. Pleopod 4 exopod with ~76 PMS,<br />
endopod with ~ 4 PMS. Pleopod 5 exopod with ~67<br />
PMS. Exopods <strong>of</strong> pleopods –5 each with distolateral<br />
margin not deeply serrate; endopods <strong>of</strong> pleopods 3–5<br />
each with mediodistal point; pleopods 2–5 peduncle<br />
distolateral margin with prominent acute RS.<br />
Uropod peduncle posterior lobe about one-half<br />
as long as endopod. Uropod rami extending beyond<br />
pleotelson. Endopod apically not bifid, lateral margin<br />
straight, without prominent excision, proximal lateral<br />
margin with RS, distal lateral margin with 2 RS, mesial<br />
margin weakly convex, with 7 RS. Exopod not extending<br />
to end <strong>of</strong> endopod, 2.3 times as long as greatest<br />
width, apically not bifid; lateral margin convex, with<br />
3 RS; mesial margin convex, with 5 RS.<br />
femaLe: <strong>The</strong> single female, taken at the same station as<br />
the <strong>New</strong> <strong>Zealand</strong> male, differs in a number <strong>of</strong> minor<br />
characters, and its identity is here regarded as provisional<br />
(see ‘Remarks’).<br />
size: <strong>New</strong> <strong>Zealand</strong> material examined here 6.5– 7.0<br />
mm; specimens from Bouvet Island, 6.0–25 mm.<br />
Variation: Robust setae: pleotelson RS 4+4 (holotype)<br />
and 5+5 (<strong>New</strong> <strong>Zealand</strong>). <strong>The</strong> holotype had only one<br />
uropod, so no assessment was possible for uropodal<br />
robust setae. <strong>The</strong> robust setae <strong>of</strong> the pereopods were<br />
consistent between specimens, as illustrated (Figs 79F,<br />
82). <strong>The</strong> Bouvet Island specimens are in good condition,<br />
and have variable counts for robust setae: pleotelson<br />
RS 4+4, 5+5 and 7+7; uropodal endopod mesial margin<br />
7 (three times), 9 (twice) and 0 (once), lateral margin<br />
+2 (all); uropodal exopod lateral margin with 0– 2<br />
RS, mesial with 5 (twice) or 6 (four times). <strong>The</strong> coxae <strong>of</strong><br />
pereonites 5–7 on the Bouvet Island specimens have a<br />
weakly concave posterior margin, while this is straight<br />
in the <strong>New</strong> <strong>Zealand</strong> specimens and the holotype.<br />
Bouvet Island specimens dorsally brown, laterally<br />
with patches <strong>of</strong> white chromatophores on all or some<br />
<strong>of</strong> pereonites 4–6.<br />
remarks: <strong>The</strong> mesially narrow eyes, rectangular posterior<br />
margins <strong>of</strong> coxal plates 2–4, wide and short frontal<br />
lamina with the posterior margin forming a distinctly<br />
concave blade, close-set penial openings in the males<br />
and the presence (and pattern and number) <strong>of</strong> robust<br />
setae on the uropods and pleotelson all allow identification<br />
<strong>of</strong> this species.<br />
40<br />
Aegiochus pushkini belongs with a group <strong>of</strong> Aegiochus<br />
species characterised by their relatively small size,<br />
the posterior margin <strong>of</strong> the frontal lamina forming a<br />
blade and relatively slender pereopods armed with<br />
small robust setae. Two <strong>of</strong> these, Aegiochus nohinohi sp.<br />
nov. and A. gordoni sp. nov., can be distinguished by<br />
the rami <strong>of</strong> pleopod and 2 being deeply serrate and<br />
lacking robust setae on the pleotelson. <strong>The</strong> remaining<br />
species with similar pereopodal, uropodal and pleotelson<br />
morphology can be separated by having the eyes<br />
united (Aegiochus kakai sp. nov. and A. kanohi sp. nov.)<br />
or by having the distal margin <strong>of</strong> the propodal palm <strong>of</strong><br />
pereopods –3 expanded (A. piihuka sp. nov.).<br />
Other superficially similar species from the southern<br />
Indian Ocean are Aegiochus crozetensis (Kussakin &<br />
Vasina, 982), see p. 237, and A. uschakovi (Kussakin,<br />
967) see p. 24 . <strong>The</strong> former has far larger eyes and a<br />
narrower frontal lamina which has a convex posterior<br />
margin, while the latter also has a narrower frontal<br />
lamina and also has a far wider uropod endopod.<br />
<strong>The</strong> specimens from three widely distant locations<br />
show a degree <strong>of</strong> variation not usually observed within<br />
a species. <strong>The</strong> holotype and male <strong>New</strong> <strong>Zealand</strong> specimen<br />
agree closely, differing principally in the male<br />
having a wider frontal lamina and two distal robust<br />
setae on pereopod 2 merus. <strong>The</strong> female <strong>New</strong> <strong>Zealand</strong><br />
specimen is in good condition and shows rather more<br />
differences that can be ascribed to sexual dimorphism.<br />
<strong>The</strong>se differences include a more strongly serrated<br />
and posteriorly wider pleotelson margin, the antennal<br />
flagellum extending to the posterior <strong>of</strong> pereonite<br />
3 (it is more usual in dimorphic species that the male<br />
has the longer antennal flagellae), coxae 7 are slightly<br />
more acute. <strong>The</strong> specimens from Bouvet Island are in<br />
close agreement with the <strong>New</strong> <strong>Zealand</strong> female, but<br />
show considerable variation in pleotelson robust setae<br />
(from 8 to 4), the large female has the posterior margin<br />
<strong>of</strong> coxae 5–7 weakly concave and the small specimen<br />
is similar to the <strong>New</strong> <strong>Zealand</strong> material; the pereopod<br />
and uropod morphology seems identical in all <strong>of</strong> these<br />
specimens. Without a larger series <strong>of</strong> specimens from<br />
one location it is not possible to further assess the consistency<br />
<strong>of</strong> such differences. In view <strong>of</strong> these differences<br />
I consider the <strong>New</strong> <strong>Zealand</strong> male to be the same as<br />
the holotype, but the remaining specimens to be only<br />
provisionally determined as this species.<br />
prey: Not known.<br />
distribution: Recorded from the southern Indian Ocean<br />
(Kussakin & Vasina 982); in the <strong>New</strong> <strong>Zealand</strong> region<br />
<strong>of</strong>f southern South Island, Macquarie Ridge; <strong>of</strong>f Bouvet<br />
Island (to Norway), South Atlantic; all localities<br />
between 52° and 54° South. At depths from 22 to 4 0<br />
metres.
Aegiochus riwha sp. nov. (Figs 84–87)<br />
materiaL examined: Holotype. ♂ (27 mm), west <strong>of</strong> North<br />
Island, <strong>New</strong> Caledonia Trough, 37°30.55’S, 72° 3.60’E,<br />
23 April 2000, sea mount, 000 m, coll. RV Kaharoa<br />
(NIWA 7933).<br />
Paratype. ♀ (33 mm), west <strong>of</strong> North Island, <strong>New</strong> Caledonia<br />
Trough, 37°30.3 ’S, 72° 3.68’E, 24 April 2000,<br />
sea mount, 060 m, coll. RV Kaharoa (NIWA 7934).<br />
Additional material: tasman sea: Crushed specimen,<br />
west <strong>of</strong> North Island, <strong>New</strong> Caledonia Trough,<br />
37°30.3 ’S, 72° 3.68’E, 24 April 2000, sea mount, 060<br />
m, (NIWA 7935). ♀ (non-ovig. 28 mm), West Norfolk<br />
Ridge, 34° 7.84’S, 68°25.82’E, 2 June 2003, 25 – 268<br />
m, coll. NORFANZ (NIWA 7937). ♀ (non-ovig. 38<br />
mm), West Norfolk Ridge, 34° 4.33’S, 68°2 . 8’E,<br />
3 June 2003, 95– 202 m, coll. NORFANZ (NIWA<br />
7938). pacific <strong>New</strong> <strong>Zealand</strong>: ♀ (crushed ovig.),<br />
Chatham Rise, Shipley Sea Mount, 4 .80 2–8005°S,<br />
80.5065–4967°E, 6 June 2006, 240– 275 m (NIWA<br />
25672). taiwan: ♂ (3 mm), <strong>of</strong>f Taiwan, 22° 6.56’N,<br />
20°6. ’E, 22 November 2001, 736– 040 m, Otter Trawl<br />
(Le Drézén type JUNEAUX), stn CD 34, coll. RV Ocean<br />
Researcher (MTQ W 368 ). <strong>New</strong> caledonia, HALIPRO<br />
2, coll. B. Richer de Forges: 2♀ (34 mm non-ovig,<br />
damaged ~25 mm), 25°20’S, 68°55’E, 2 November<br />
996, stn. BT32, 697– 340 m (MNHN Is.5858); ♀ (ovig,<br />
40 mm), 25°04’S, 68°44’E, 7 November 966, stn.<br />
BT55, 098– 480 (MNHN Is.5859); ♂ (30 mm), ♀ (42<br />
mm, non-ovig.), 25°52’S, 68°44’E, 8 November 996,<br />
stn. BT60, 33– 280 m (MNHN Is.5857).<br />
desCription: Body 2.2 times as long as greatest width,<br />
dorsal surfaces smooth, sparsely punctate (finely<br />
pilose), widest at pereonite 5, lateral margins subparallel.<br />
Rostral point ventrally directed. Eyes large,<br />
not medially united, separated by about 0% width<br />
<strong>of</strong> head; each eye made up <strong>of</strong> ~22 transverse rows <strong>of</strong><br />
ommatidia, each row with ~9 ommatidia; eye colour<br />
black. Pereonite 1 and coxae 2–3 each with posteroventral<br />
angle rounded. Coxae 5–7 with entire oblique carina;<br />
posterior margins convex, posterolateral angle blunt<br />
(more than 45°). Pleon with pleonite visible in dorsal<br />
view; pleonite 4 with posterolateral margins extending<br />
to but not beyond posterior margin <strong>of</strong> pleonite 5; pleonite<br />
5 with posterolateral angles free, not overlapped<br />
by lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.7 times as<br />
long as anterior width, dorsal surface without longitudinal<br />
carina; lateral margins convex, serrate (weakly),<br />
posterior margin with truncate median excision, with<br />
– 2 RS (each set within serration).<br />
Antennule peduncle articles 3 and 4 .2 times as<br />
long as combined lengths <strong>of</strong> articles and 2, article 3<br />
3.4 times as long as wide; flagellum with 17 articles,<br />
4<br />
extending to anterior <strong>of</strong> pereonite . Antenna peduncle<br />
article 4 .3 times as long as wide, 0.8 times as long as<br />
combined lengths <strong>of</strong> articles –3, inferior margin 0 plumose<br />
setae, and 0 short simple setae; article 5 .3 times<br />
as long as article 4, 2.6 times as long as wide, inferior<br />
margin with palmate seta, anterodistal angle with<br />
cluster <strong>of</strong> 3 short simple setae (and 1 palmate); flagellum<br />
with 27–30 articles, extending to pereonite 4.<br />
Frontal lamina flat, as wide as long, diamond shaped,<br />
with lateral margins converging posteriorly, anterior<br />
margin acute, forming median angle, posterior margin<br />
forming narrow stem.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 2 distolateral setae ( long,<br />
remainder short), palp article 3 with 34 setae. Maxillule<br />
with 0 terminal RS ( large, 9 slender). Maxilla mesial<br />
lobe with 3 RS (2 hooked, straight); lateral lobe with 3<br />
RS (hooked). Maxilliped endite with 4 apical setae (flat,<br />
blade-like, with CPS); palp article 2 with 4 RS (small<br />
hooked); article 3 with 8 recurved RS (4 hooked lateral<br />
and 4 long slender distal); article 4 with 4 hooked RS;<br />
article 5 with 4 RS (3 straight, hooked).<br />
Pereopod 1 basis 2.4 times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 2 RS (acute); merus<br />
inferior margin with 2 RS (minute), set as proximal<br />
group, superior distal angle with 0 RS (2 slender<br />
setae); carpus 0.9 as long as merus, inferior margin<br />
with RS (minute); propodus 2. times as long as<br />
proximal width, inferior margin with 0 RS, propodal<br />
palm simple, without blade or process, dactylus<br />
smoothly curved, .0 as long as propodus. Pereopod<br />
2 ischium inferior margin with 0 RS, superior distal<br />
margin with RS; merus inferior margin with 4 RS,<br />
set as single row (widely spaced series), superior distal<br />
margin with acute RS; carpus similar in size to that<br />
<strong>of</strong> pereopod , inferodistal angle with 2 RS. Pereopod 3<br />
similar to pereopod 2. Pereopods 5–7 inferior margins<br />
<strong>of</strong> ischium–carpus with short RS. Pereopod 6 similar to<br />
pereopod 7. Pereopod 7 basis 2.3 times as long as greatest<br />
width, inferior margins with 2 palmate setae; ischium<br />
0.5 as long as basis, inferior margin with 3 RS (set as<br />
and 2), superior distal angle with 3 RS, inferior distal<br />
angle with 4 RS; merus . as long as ischium, 2.0 as<br />
long as wide, inferior margin with 4 RS (set as 3 and ),<br />
superior distal angle with 0 RS, inferior distal angle<br />
with 8 RS; carpus .0 as long as ischium, 2.2 times as<br />
long as wide, inferior margin with 3 RS (set as and 2),<br />
superior distal angle with 4 RS, inferior distal angle<br />
with 8 RS; propodus 0.9 as long as ischium, 3. times<br />
as long as wide, inferior margin with 3 RS (set as 2 and<br />
), superior distal angle with acute RS (plus 3 slender<br />
setae), inferior distal angle with 2 RS.<br />
Penes low tubercles; penial openings separated by<br />
6% <strong>of</strong> sternal width.
Figure 84. Aegiochus riwha sp. nov. I, J male paratype, NIWA 7934; remainder holotype. A, dorsal view; B, lateral view;<br />
C, head; D, frons; E, frons, anterior view; F, pleotelson; G, pleotelson, posterior margin; H, sternite 7 showing penial openings;<br />
I, antennule; J, antenna peduncle.<br />
Pleopod 1 exopod .5 times as long as wide, distally<br />
broadly rounded, lateral margin weakly convex,<br />
mesial margin strongly convex, with PMS on distal<br />
one-third; distally rounded, lateral margin strongly<br />
concave, with PMS on distal one-third, mesial margin<br />
42<br />
with PMS on distal one-third; peduncle .6 times as<br />
wide as long, mesial margin with 9 coupling hooks.<br />
Pleopod 2 appendix masculina distally widest, 0.9 times<br />
as long as endopod, distally bluntly rounded. Exopods<br />
<strong>of</strong> pleopods –3 each with distolateral margin not digi-
Figure 85. Aegiochus riwha sp. nov. Male paratype, NIWA 7934. A, mandible; B, mandible palp, article 3; C, maxillule;<br />
D, maxillule apex; E, maxilla; F, maxilla apex; G, maxilliped; H, maxilliped articles 2–5.<br />
tate; endopods <strong>of</strong> pleopods 3–5 each with distolateral<br />
point; pleopods 2–4 peduncle distolateral margin with<br />
prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about three-quarters as long as endopod.<br />
Uropod rami extending beyond pleotelson. Endopod<br />
apically deeply bifid, mesial process prominent, lateral<br />
margin sinuate (very weakly sinuate), without<br />
prominent excision, proximal lateral margin with 0 RS,<br />
distal lateral margin with 2 RS, mesial margin strongly<br />
convex, with 9– 0 RS. Exopod extending beyond end <strong>of</strong><br />
endopod (slightly), 2.7 times as long as greatest width,<br />
apically deeply bifid, mesial process prominent; lateral<br />
margin weakly convex, with 2– 3 RS; mesial margin<br />
straight (or very weakly concave, distally convex),<br />
with 5 RS.<br />
43<br />
femaLe: Similar to male.<br />
size: Specimens from the <strong>New</strong> <strong>Zealand</strong> region: males<br />
27–33 mm, non-ovigerous females 28–33 mm; largest<br />
size a 42 mm female from <strong>New</strong> Caledonia.
Figure 86. Aegiochus riwha sp. nov. C, male paratype, NIWA 7934; remainder holotype. A–C, pereopods , 2 and 7,<br />
respectively; D, pereopod , inferior margin <strong>of</strong> merus and carpus.<br />
Variation: <strong>The</strong>re were only two intact specimens<br />
available at time <strong>of</strong> description, therefore the details<br />
here are <strong>of</strong> range only. Robust setae on the pleotelson<br />
ranged from 0 or . Uropod endopod mesial margin<br />
8– 2 RS, lateral margin always 2 RS; uropod exopod<br />
mesial margin 5 or 6 RS, lateral margin 2 or 3 RS. In<br />
both specimens there were 3 minute RS set within the<br />
apical notch.<br />
remarks: Aegiochus riwha sp. nov. is most similar to<br />
A. beri, but has coxae that are more blunt, more robust<br />
pereopods with a shorter dactylus ( .0 times as long<br />
as propodus v. .4 times as long in A. beri), ventral<br />
surface <strong>of</strong> frontal lamina is flat (angled in A. beri), the<br />
uropod rami and pleotelson margins are weakly serrate<br />
(strongly serrate in A. beri), the appendix masculina is<br />
weakly spatulate and as long as the endopod (terminally<br />
acute and shorter than the endopod in A. beri)<br />
and the apices <strong>of</strong> the uropodal rami are more widely<br />
bifid that in A. beri (narrowly bifid). In addition A. riwha<br />
has all dorsal body surfaces covered by brown chro-<br />
44<br />
matophores (A. beri is without such both in holotype<br />
and Lord Howe material), although without access to<br />
fresh or freshly preserved material it is not possible to<br />
place much reliance on colour difference. Given the<br />
proximity <strong>of</strong> the locations for the two species and the<br />
possibility <strong>of</strong> their being sympatric in part, these differences<br />
are considered to be <strong>of</strong> species-specific value.<br />
<strong>The</strong> difference in coxal shape readily and easily<br />
distinguishes Aegiochus riwha from Aegiochus beri. <strong>The</strong><br />
anteriorly truncate notch on the pleotelson posterior<br />
margin separates these two species from all other<br />
southwestern Pacific and Southern Ocean species.<br />
Aegiochus quadratisinus (Richardson, 903) (Bruce 983)<br />
is known only from Hawai’i but, while similar to A.<br />
riwha, has smaller eyes and the anterior margins <strong>of</strong> the<br />
frontal lamina are concave.<br />
<strong>The</strong> maxilla appears to have the mesial lobe fused to<br />
the larger lateral lobe, and I could not distinguish the<br />
point <strong>of</strong> join under stereomicroscopy nor compound<br />
microscopy using DIC light interference.
Figure 87. Aegiochus riwha sp. nov. C–E, male paratype, NIWA 7934; remainder holotype. A–D, pleopods –3, 5 respectively;<br />
E, uropod; F, uropod endopod, ventral view; G, exopod apex; H, endopod apex.<br />
prey: <strong>The</strong>re are no records.<br />
distribution: Northwestern <strong>New</strong> <strong>Zealand</strong>, and the<br />
southern West Norfolk Ridge; also <strong>of</strong>f <strong>New</strong> Caledonia,<br />
and in the northwestern Pacific from <strong>of</strong>f Taiwan.<br />
Potentially at depths <strong>of</strong> 697– 480 metres.<br />
etymoLogy: Riwha is a Mäori word meaning cleft or<br />
notch, in reference to the excision in the pleotelson<br />
(noun in apposition).<br />
45
Aegiochus tara sp. nov. (Figs 88–90)<br />
materiaL examined: Holotype, ♀ (non-ovig. 3 mm),<br />
West Norfolk Ridge, 33°42.45’S, 67°27.03’E, 28 May<br />
2003, 45 – 478 m, coll. NORFANZ, RV Tangaroa<br />
(NIWA 7947).<br />
Paratype. ♀ (non-ovig. 22 mm), Tasmania, 48 km east<br />
<strong>of</strong> Cape Toureville, 42°00.25’S, 48°43.55’E, 30 October<br />
980, 264 m, gravel with sandy mud, G.C.B. Poore &<br />
co. (NMV J277 2).<br />
Other material: Comparative material <strong>of</strong> Aegiochus<br />
plebeia, see Appendix 2.<br />
desCription: Body .6 times as long as greatest width,<br />
dorsal surfaces polished in appearance and sparsely<br />
punctate, widest at pereonite 5, lateral margins weakly<br />
ovate. Rostral point folded ventrally and posteriorly.<br />
Eyes large, not medially united, separated by about<br />
4% width <strong>of</strong> head; each eye made up <strong>of</strong> ~ 2 transverse<br />
rows <strong>of</strong> ommatidia, each row with ~22 ommatidia;<br />
eye colour dark brown. Pereonite 1 and coxae 2–3 each<br />
with posteroventral angle acute, posteriorly produced.<br />
Coxae 5–7 with incomplete oblique carina; posterior<br />
margins concave, posterolateral angle acute (less than<br />
45°). Pleon with pleonite visible in dorsal view; pleonite<br />
4 with posterolateral margins extending to but not<br />
beyond posterior margin <strong>of</strong> pleonite 5; pleonite 5 with<br />
posterolateral angles free, not overlapped by lateral<br />
margins <strong>of</strong> pleonite 4. Pleotelson 0.9 times as long as<br />
anterior width, dorsal surface without longitudinal<br />
carina; lateral margins sinuate (posteriorly weakly<br />
concave), smooth, posterior margin converging to<br />
caudomedial point, with 4 RS (estimated).<br />
Antennule peduncle articles 3 and 4 . times as long<br />
as combined lengths <strong>of</strong> articles and 2, article 3 3.3<br />
times as long as wide (narrowing distally); flagellum<br />
with 9 articles, extending to posterior <strong>of</strong> pereonite .<br />
Antenna peduncle article 4 2.5 times as long as wide,<br />
.2 times as long as combined lengths <strong>of</strong> articles –3,<br />
inferior margin 0 plumose setae, and short simple<br />
seta; article 5 .0 times as long as article 4, 3.0 times<br />
as long as wide, inferior margin with 0 palmate setae,<br />
anterodistal angle with cluster <strong>of</strong> 2 short simple setae;<br />
flagellum with 20 articles (terminal article/s missing),<br />
extending to middle <strong>of</strong> pereonite 3.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, posteriorly<br />
rounded, anterior margin acute, forming median angle,<br />
posterior margin not abutting clypeus.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 7 distolateral setae (simple and<br />
biserrate; with distal submarginal row 7 simple setae),<br />
palp article 3 with 25 setae (simple; distal most short,<br />
next 3 markedly longer than remainder). Maxillule with<br />
6 terminal RS (three largest being weakly hooked).<br />
46<br />
Maxilla mesial lobe with 3 RS (2 simple, serrate);<br />
lateral lobe with 7 RS (4 hooked, 3 short and straight).<br />
Maxilliped endite with 0 apical setae; palp article 2 with<br />
3 RS; article 3 with 8 recurved RS (straight or weakly<br />
recurved); article 4 with 4 hooked RS; article 5 with 4<br />
RS (two longest <strong>of</strong> which are biserrate).<br />
Pereopod 1 basis 2.8 times as long as greatest width<br />
(narrowing distally); ischium 0.3 times as long as basis,<br />
inferior margin with 0 RS, superior distal margin with<br />
RS; merus inferior margin with 0 RS, superior distal<br />
angle with 0 RS ( simple seta); carpus 0.7 as long as<br />
merus, inferior margin with 0 RS; propodus 2.9 times as<br />
long as proximal width, inferior margin with RS, propodal<br />
palm with small distal lobe, dactylus smoothly<br />
curved, .0 as long as propodus. Pereopod 2 ischium<br />
inferior margin with 0 RS, superior distal margin with<br />
RS; merus inferior margin with 3 RS (all small), set as<br />
two groups, superior distal margin with 0 acute RS (2<br />
simple setae); carpus similar in size to that <strong>of</strong> pereopod<br />
, inferodistal angle with RS (large weakly curved).<br />
Pereopod 3 similar to pereopod 2. Pereopods 5–7 inferior<br />
margins <strong>of</strong> ischium–carpus with short RS. Pereopod 6<br />
similar to pereopod 7. Pereopod 7 basis 4.4 times as long<br />
as greatest width, inferior margins with 23 palmate<br />
setae; ischium 0.4 as long as basis, inferior margin with<br />
RS, superior distal angle with 4 RS, inferior distal<br />
angle with 5 RS; merus .7 as long as ischium, 3.5 times<br />
as long as wide, inferior margin with 5 RS (set as 2, 2<br />
and ), superior distal angle with 4 RS, inferior distal<br />
angle with 6 RS; carpus .3 as long as ischium, 3.7 times<br />
as long as wide, inferior margin with 3 RS (set singly),<br />
superior distal angle with RS, inferior distal angle<br />
with 5 RS; propodus .0 as long as ischium, 4.6 times<br />
as long as wide, inferior margin with 4 RS (set as ,<br />
and 2), superior distal angle with slender seta (and<br />
RS), inferior distal angle with 2 RS.<br />
Pleopod 1 exopod .7 times as long as wide, distally<br />
narrowly rounded, mesial margin weakly oblique,<br />
lateral margin weakly convex, mesial margin strongly<br />
convex, with PMS on distal one-third; lateral margin<br />
strongly concave, with PMS on distal one-third, mesial<br />
margin with PMS on distal half; peduncle .7 times as<br />
wide as long, mesial margin with 9 coupling hooks.<br />
Exopods <strong>of</strong> pleopods –3 each with distolateral margin<br />
not digitate; endopods <strong>of</strong> pleopods 3–5 each with<br />
distolateral point; pleopods 2–4 peduncle distolateral<br />
margin with prominent acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about one-half as long as endopod.<br />
Uropod rami extending beyond pleotelson. Endopod<br />
apically not bifid, lateral margin straight, without<br />
prominent excision, proximal lateral margin with RS,<br />
distal lateral margin with 2 RS, mesial margin weakly<br />
convex, with 7 RS. Exopod extending to end <strong>of</strong> endopod,<br />
3.0 as long as greatest width, apically not bifid; lateral
Figure 88. Aegiochus tara sp. nov. Holotype. A, dorsal view; B, lateral view; C, head; D, frons, ventral view; E, pleotelson;<br />
F, pleonites, lateral margins; G, antennule; H, antenna peduncle.<br />
47
Figure 89. Aegiochus tara sp. nov. Holotype. A, mandible; B, mandible palp article 3; C, mandible incisor; D, robust seta,<br />
manidible palp article 2; E, maxillule; F, maxillule apex; G, maxilla; H, maxilla apex; I, maxilliped; J, maxilliped articles 2–5;<br />
K, largest seta, maxilliped palp article 5.<br />
margin weakly convex, with 2 RS; mesial margin<br />
weakly convex, with 4 RS.<br />
maLe: Not known.<br />
size: Female 3 mm.<br />
Variation: Most pleotelson robust setae are missing<br />
although the sockets are clearly distinct. <strong>The</strong> apices <strong>of</strong><br />
the uropodal rami and the pleotelson are all damaged,<br />
and while counts appeared the same for all margins,<br />
the numbers can be regarded only as estimates.<br />
remarks: Although similar to the sympatric Aegiochus<br />
piihuka sp. nov., Aegiochus tara sp. nov. is readily iden-<br />
48<br />
tified by the large but separate eyes, small distal lobe<br />
on the propodus <strong>of</strong> pereopods –3, that lobe bearing<br />
a prominent robust seta, the acute and laterally expressed<br />
coxal plates, the sinuate margins to the pleotelson,<br />
the apex <strong>of</strong> which is relatively strongly produced,<br />
and also the proximal one-third <strong>of</strong> the lateral margin<br />
<strong>of</strong> the uropodal exopod is devoid <strong>of</strong> setae.<br />
Aegiochus plebeia (Hansen, 890), a species <strong>of</strong> uncertain<br />
distribution (see p. 238), is also similar. Examination<br />
<strong>of</strong> the types shows that A. tara has a much narrower<br />
uropod exopod, terminally more acute pleotelson apex,<br />
smaller distal robust seta on the propodus <strong>of</strong> pereopods<br />
–3 and the coxae are strongly splayed and acute (not<br />
splayed and posteriorly produced as in A. plebeia).
Figure 90. Aegiochus tara sp. nov. Holotype. A, pereopod ; B, pereopod 2; C, pereopod 7; D, uropod exopod, ventral view;<br />
E, pleopod ; F, pleopod 2; G, uropod.<br />
Aegiochus ventrosa (Sars, 859), a North Atlantic species<br />
(see ‘Remarks’ for A. plebeia, see p. 238) is another<br />
similar species. A. ventrosa has been figured both with<br />
and without a prominent robust seta and weak propodal<br />
lobe on the palm <strong>of</strong> the propodus <strong>of</strong> pereopods –3,<br />
but pereopods –3 are more robust (propodus twice as<br />
long as wide) than those <strong>of</strong> Aegiochus tara (three times<br />
as long as wide).<br />
prey: Not known.<br />
distribution: Tasman Sea; east <strong>of</strong> Cape Toureville,<br />
Tasmania and the West Norfolk Ridge to the west <strong>of</strong><br />
northern <strong>New</strong> <strong>Zealand</strong>; 264– 478 metres.<br />
etymoLogy: <strong>The</strong> epithet, tara, is a Mäori word meaning<br />
spine or spiny in relation to marine animals.<br />
49
Aegiochus vigilans (Haswell, 88 ), comb. nov.<br />
(Fig. 9 )<br />
restriCted synonymy:<br />
Aega ommatophylax Stebbing, 905: 2, pls IV, VA.<br />
Aega dubia Richardson, 1910: 12, fig. 2.<br />
Aega (Rhamphion) vigilans.– Bruce, 1993: 762, figs 3, 4 (citations<br />
therein).<br />
Aega giganteoculata Nunomura, 1988a: 19, figs 1, 2 (new<br />
synonymy).<br />
type LoCaLity: Holborn Island, near Port Dennison,<br />
Queensland (Springthorpe & Lowry, 994: 64).<br />
materiaL examined: Manca (7.0 mm), <strong>of</strong>f Great Barrier<br />
Island, North Island, January 2006, old longline gear<br />
at ~500 m, coll. Steve Lowe (NIWA 23779). ♀ (nonovig.<br />
3 mm), <strong>New</strong> Caledonia, 2 °03.680–03.997’S,<br />
60°44.766–44.874’E, 2 October 2005, DW2636, 254–<br />
27 m, coll. B. Richer de Forge (MNH Is.59 0).<br />
Also examined: ♀ (non-ovig. 6.5 mm), northwest<br />
<strong>of</strong> Bluff Point, Geraldton, Western Australia, 27°40’S,<br />
50<br />
3°03’E, 22 March 963, 28 m, CSIRO stn 3 (WAM<br />
2293-86). 4, 93 km west <strong>of</strong> Dongara, Western Australia,<br />
20°07’S, 3°57’E, 9 February 976, 0 m, stn 30<br />
(WAM 2282-86). Paratypes <strong>of</strong> Aega giganteocula, two<br />
lots, heavily dissected (body no longer intact), <strong>of</strong>f Itoman,<br />
Okinawa, Japan, July 985, coll. Hideo Sekiguchi<br />
(TSM Cr7649, Cr7650). , <strong>of</strong>f Singapore, 3 May 95 ,<br />
39 m, coral, Galathea stn 355 (ZMUC unreg).<br />
remarks: A detailed redescription <strong>of</strong> this widely distributed<br />
species was given by Bruce ( 983). As the<br />
sole specimen from <strong>New</strong> <strong>Zealand</strong> waters is a manca, a<br />
detailed redescription is not given, particularly as mature<br />
adult males possess two large forward-projecting<br />
processes on pereonite , and the rostrum is also produced,<br />
giving these animals a striking ‘three-horned’<br />
appearance, abundantly different from all other species<br />
in the genus. Females and immature specimens can<br />
be identified by the huge, black, united eyes filling<br />
the head in dorsal view, the characteristic scalloping<br />
<strong>of</strong> the posterior margin <strong>of</strong> the pleotelson, shape <strong>of</strong> the<br />
Figure 91. Aegiochus vigilans (Haswell, 88 ). From Stebbing ( 905). A, dorsal view; B, head, lateral view; C–E, pereopods ,<br />
2 and 7 respectively; F, pleotelson and uropods, ventral view; G, pleopod ; H, pleopod 2; I, uropod.
uropods, and the elongate and flat (‘cirolanid-like’)<br />
frontal lamina. Comparison with specimens from tropical<br />
Australia (Australian Museum, Western Australian<br />
Museum) confirm the identity <strong>of</strong> this immature specimen,<br />
the most southerly record for the species.<br />
Aegiochus vigilans shows a number <strong>of</strong> unique character<br />
states these being, in addition to the strongly<br />
dimorphic ornamentation <strong>of</strong> the mature males, a flat<br />
and elongate frontal lamina, cirolanid-like spination <strong>of</strong><br />
the mandibular molar process, large blunt robust seta<br />
at the inferodistal margin <strong>of</strong> the carpus <strong>of</strong> pereopods<br />
2 and 3, an extremely long appendix masculina (more<br />
than twice as long as the endopod), and paired narrow<br />
penial processes; the maxilliped palp differs from others<br />
in that article 4 is comparatively small.<br />
Examination <strong>of</strong> paratypic material <strong>of</strong> Aega giganteocula<br />
Nunomura, 998, shows that it agrees entirely<br />
with the description <strong>of</strong> Aegiochus vigilans given by<br />
Bruce ( 983); I have no hesitation in making the<br />
synonymy.<br />
distribution: One record from northeastern <strong>New</strong> <strong>Zealand</strong>.<br />
Commonly recorded within the tropics from India<br />
to eastern Australia and the Philippines (Bruce 983);<br />
from shallow water on coral reefs to a depth <strong>of</strong> 27 m,<br />
possibly deeper. Collections examined at the Muséum<br />
national d’Histoire naturelle in Paris indicate that the<br />
species is common in <strong>New</strong> Caledonian waters.<br />
Aegiochus sp.<br />
materiaL examined: ♂ (damaged, 9.5 mm), Parengarenga<br />
Harbour, 2 Feb 974, Zostera and sand, 6–9<br />
m, coll. RV Acheron (AK 4607).<br />
remarks: This specimen, unfortunately in very poor<br />
condition, appears most similar to Aegiochus nohinohi<br />
sp. nov. Both species have relatively small eyes, similar<br />
pattern <strong>of</strong> setation on the pereopods, serrate pleopodal<br />
endopods, and the pleotelson without robust setae. A<br />
number <strong>of</strong> differences suggest that this specimen may<br />
be a distinct species: the frontal lamina appears to be a<br />
ventrally flat triangle (similar to that <strong>of</strong> Aegiochus pushkini),<br />
pereopods 2 and 3 have prominent robust setae<br />
on the inferior margin <strong>of</strong> the merus, and the uropod<br />
exopod and endopod each have two prominent robust<br />
setae on the mesial margins. Given the very different<br />
habitats <strong>of</strong> the two species (Parengarenga Harbour is<br />
a largely enclosed marine estuary, while A. nohinohi is<br />
known from oceanic shelf habitats, 360 to 40 metres)<br />
it seems very unlikely that this specimen is A. nohinohi.<br />
Unfortunately the specimen is too badly damaged to<br />
describe, and additional specimens would be necessary<br />
to adequately characterise the species.<br />
5<br />
Aegiochus sp.<br />
materiaL examined: ♀ (ovig. 2.9 mm), Rumble 3 sea<br />
mount, 78°29.79’E, 35°44.30’S, 20 May 200 , scoria<br />
rubble, 939–250 m, coll. RV Tangaroa (NIWA 34807).<br />
remarks: This specimen is very similar to Aegiochus<br />
kanohi sp. nov., but differs notably in the shape <strong>of</strong> the<br />
frontal lamina, which does not have a blade-like posterior<br />
margin, pereopod 2 which has one robust seta<br />
on the propodal palm, and in having robust setae on<br />
the pleotelson posterior margins. <strong>The</strong>se are diagnostic<br />
characters, but without more specimens, including<br />
mature males, the species cannot adequately be characterised,<br />
nor differentiated from other similar sympatric<br />
species. Until further material is available it remains<br />
identified as Aegiochus sp.<br />
Epulaega gen. nov.<br />
type speCies: Aega fracta Hale, 940, here designated.<br />
speCies inCLuded: In addition to type species— Epulaega<br />
derkoma sp. nov., Epulaega nodosa (Schioedte & Meinert,<br />
879), comb. nov., Epulaega lethrina (Bruce, 983) comb.<br />
nov. and E. monilis (Barnard, 9 4) comb. nov.<br />
diagnosis: Rostrum minute, not visible in dorsal view.<br />
Eyes large, united. Antennule and antenna peduncle<br />
articles not expanded or produced. Maxillule with single<br />
large broad-based and several small RS. Maxilliped<br />
palp 5-articled, article shorter than wide, article 5<br />
minute. Penial processes medially fused, with separate<br />
openings.<br />
desCription: Body moderately to strongly vaulted,<br />
about 2 to 4 times as long as wide. Eyes large, medially<br />
united. Rostral point ventral or anteroventral, minute.<br />
Coxae 4–7 longer than respective segment, posteriorly<br />
produced. Pleon not abruptly narrower than pereon;<br />
pleonites all visible, pleon not widest posteriorly, pleonite<br />
5 laterally overlapped by pleonite 4; pleonites 3–5<br />
posteriorly produced to an acute point.<br />
Frontal lamina present, ovate, not in contact with labrum.<br />
Mandible with uni- or bicuspid incisor. Maxillule<br />
with 1 large flat terminal broad-based RS, several small<br />
RS. Maxilla lateral lobe with 3–5 terminal hooked RS,<br />
endite with 2–3 hooked RS. Maxilliped palp 5-articled,<br />
article shorter than wide, articles 3 and 4 each with<br />
2–6 stout recurved RS, article 5 minute, vestigial.<br />
remarks: All species <strong>of</strong> Epulaega gen. nov. have large<br />
medially united eyes, lack an obvious rostrum in dorsal<br />
view, maxilliped palp article 5 is a vestigial lobe,<br />
and the penial papillae are medially fused. <strong>The</strong> latter<br />
three states are defining apomorphies for the genus.
Most species are loosely associated with sponges, but<br />
Epulaega lethrina is unambiguously but not exclusively<br />
a commensal <strong>of</strong> coral reef fishes (principally Serranidae<br />
and Lethrinidae) on the Great Barrier Reef (Bruce<br />
983). On present records the genus has an Indo-West<br />
Pacific distribution to approximately 42° South in <strong>New</strong><br />
<strong>Zealand</strong>.<br />
etymoLogy: From the Latin epulo (guest at a banquet or<br />
feast), in combination with Aega to indicate the family<br />
affinity. Gender feminine.<br />
Key to the new <strong>Zealand</strong> species <strong>of</strong> epulAegA<br />
Posterior margin <strong>of</strong> pereonites without nodules;<br />
dorsal surfaces <strong>of</strong> pleotelson smooth ....................<br />
...................................................E. derkoma sp. nov.<br />
– Posterior margin <strong>of</strong> pereonites with small nodules;<br />
dorsal surface <strong>of</strong> pleotelson with scattered small<br />
spines .............................................E. fracta (p. 56)<br />
Epulaega derkoma sp. nov. (Figs 92–95)<br />
materiaL: Holotype: ♀ (ovig. 7.7 mm), South Norfolk<br />
Basin, 25 July 975, 32° 0.80’S, 67°2 . 9’E, 356 m, DR<br />
Stn I96 (NIWA 23862).<br />
desCription: Body .4 times as long as greatest width,<br />
dorsal surfaces polished in appearance, widest at pereonite<br />
5, lateral margins weakly ovate. Rostral point<br />
ventrally directed. Eyes large, medially united (line <strong>of</strong><br />
separation present), anterior clear field 17% length <strong>of</strong><br />
head, posterior clear field 20% length <strong>of</strong> head; each eye<br />
made up <strong>of</strong> ~9 transverse rows <strong>of</strong> ommatidia, each row<br />
with ~7–8 ommatidia; eye colour dark brown (chestnut).<br />
Pereonite 1 and coxae 2–3 each with posteroventral<br />
angle right-angled; posterior margins <strong>of</strong> pereonites not<br />
ornamented (posterior <strong>of</strong> pereonites 2 and 3 with weak<br />
submarginal transverse ridge). Coxae 5–7 with entire<br />
oblique carina (weakly defined) posterior margins<br />
convex, posterolateral angle blunt (more than 45°).<br />
Pleon with pleonite largely concealed by pereonite 7;<br />
pleonites with lateral margin <strong>of</strong> pleonites 4–5 plate-like<br />
and flattened; pleonite 4 with posterolateral margins<br />
not extending to posterior margin <strong>of</strong> pleonite 5; pleonite<br />
5 with posterolateral angles free, not overlapped by<br />
lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.7 times as long<br />
as anterior width, dorsal surface without longitudinal<br />
carina; lateral margins convex, smooth, posterior margin<br />
converging to caudomedial point, with RS rubbed<br />
<strong>of</strong>f, only remaining.<br />
Antennule peduncle articles 3 and 4 . times as long<br />
as combined lengths <strong>of</strong> articles and 2, article 3 3.4<br />
times as long as wide; flagellum with 9 articles (article<br />
0.9 times as long as peduncle), extending to anterior<br />
<strong>of</strong> pereonite . Antenna peduncle article 4 2.3 times as<br />
52<br />
long as wide, 0.9 times as long as combined lengths <strong>of</strong><br />
articles –3, inferior margin with plumose seta and<br />
2 short simple setae (distal); article 5 .4 times as long<br />
as article 4, 4.3 times as long as wide, inferior margin<br />
with pappose seta, anterodistal angle with cluster<br />
<strong>of</strong> 1 short simple seta (and 1 pappose seta); flagellum<br />
with 2 articles, extending to posterior <strong>of</strong> pereonite 3.<br />
Frontal lamina flat, longer than greatest width, lateral<br />
margins converging posteriorly, anterior margin<br />
rounded, without small median point, posterior margin<br />
not abutting clypeus.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 0 distolateral setae (in two<br />
tiers, 4 biserrate, 4 stout simple, 2 slender simple),<br />
palp article 3 with 20 setae (terminal less than twice<br />
as long as remainder). Maxillule with 6 terminal RS (<br />
large, 3 slender, 2 minute). Maxilla mesial lobe with 2<br />
RS (circumplumose); lateral lobe with 2 RS (curved).<br />
Maxilliped palp article 2 fused to article 3; article 3 with 3<br />
straight RS; article 4 with 2 straight RS; article 5 wholly<br />
fused to article 4, with RS.<br />
Pereopod 1 basis 3.9 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
RS (small), superior distal margin with RS; merus<br />
inferior margin with RS (plus 2 slender setae), set as<br />
distal group, superior distal angle with RS (small);<br />
carpus 0.4 as long as merus; inferior margin with 0 RS;<br />
propodus 2. times as long as proximal width, inferior<br />
margin with 0 RS, propodal palm simple, without<br />
blade or process, dactylus smoothly curved, .0 as<br />
long as propodus. Pereopod 2 ischium inferior margin<br />
with RS (large blunt), superior distal margin with<br />
RS; merus inferior margin with 5 RS (large, blunt;<br />
set as , 2 and 2), set as three groups, superior distal<br />
margin with 2 acute RS; carpus similar in size to that<br />
<strong>of</strong> pereopod , inferodistal angle with 0 RS. Pereopod 3<br />
similar to pereopod 2. Pereopod 6 similar to pereopod<br />
7. Pereopod 7 basis 3.0 as long as greatest width, inferior<br />
margins with 4 palmate setae; ischium 0.4 as long as<br />
basis, inferior margin with 2 RS (set as and ), superior<br />
distal angle with 3 RS, inferior distal angle with 3<br />
RS; merus 0.9 as long as ischium, .7 times as long as<br />
wide, inferior margin with 2 RS (set as and ), superior<br />
distal angle with 6 RS, inferior distal angle with 3<br />
RS; carpus .0 as long as ischium, 2.0 times as long as<br />
wide, inferior margin with RS, superior distal angle<br />
with 8 RS (5 biserrate), inferior distal angle with 4 RS;<br />
propodus . as long as ischium, 4.0 times as long as<br />
wide, inferior margin with 2 RS (set as single cluster),<br />
superior distal angle with slender seta, inferior distal<br />
angle with 2 RS.<br />
Pleopod 1 exopod 2.0 times as long as wide, lateral<br />
margin weakly convex, mesial margin strongly convex,<br />
with PMS from distal half, with ~34 PMS; endopod 2.<br />
times as long as wide, distally rounded, lateral margin<br />
straight, with PMS from on distal margin only, mesial
Figure 92. Epulaega derkoma sp. nov. Holotype. A, dorsal view; B, lateral view; C, head; D, pleonites, lateral view; E, frons;<br />
F, antenna; G, antennule peduncle.<br />
margin with PMS from distal one-third, endopod with<br />
~ 7 PMS; peduncle .8 times as wide as long, mesial<br />
margin with 6 coupling hooks. Pleopod 2 exopod with<br />
~46 PMS, endopod with ~ 9 PMS. Pleopod 3 exopod<br />
with ~53 PMS, endopod with ~ 2 PMS. Pleopod 4 exopod<br />
with ~52 PMS, endopod with ~ 2 PMS. Pleopod 5<br />
exopod with ~48 PMS. Exopods <strong>of</strong> pleopods –3 each<br />
with distolateral margin not digitate; endopods <strong>of</strong><br />
53
Figure 93. Epulaega derkoma sp. nov. Holotype. A, mandible; B, mandible palp article 3; C, maxillule; D, maxillule apex;<br />
E, maxilla; F, maxilla apex; G, maxilliped; H, maxilliped articles 2–5.<br />
pleopods 3–5 each with distolateral point; pleopods<br />
2–5 peduncle distolateral margin with prominent<br />
acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS<br />
(and 2 plumose slender setae), posterior lobe about<br />
two-thirds as long as endopod. Endopod apically not<br />
bifid, lateral margin straight, without prominent excision,<br />
proximal lateral margin with 0 RS, distal lateral<br />
54<br />
margin with RS, mesial margin strongly convex,<br />
with 3 RS. Exopod not extending to end <strong>of</strong> endopod,<br />
3.2 times as long as greatest width, apically not bifid;<br />
lateral margin weakly convex, with 7 RS; mesial margin<br />
weakly convex, with RS.<br />
maLe: Not known.
Figure 94. Epulaega derkoma sp. nov. Holotype. A–D, pereopods –3 and 7, respectively (pereopod 3 basis omitted;<br />
pereopod 7 dactylus missing).<br />
Variation: <strong>The</strong>re being but a single specimen, variation<br />
remains unknown.<br />
remarks: Epulaega derkoma sp. nov. is a distinctive species,<br />
readily identified by the large and united eyes,<br />
antennule flagellum article 1 being as long as peduncle<br />
article 3, flattened and plate-like lateral margins <strong>of</strong> the<br />
epimera, short dactylus on pereopods –3, and pereopods<br />
2 and 3 with conspicuously large robust setae.<br />
<strong>The</strong> maxilliped <strong>of</strong> ovigerous females is usually similar<br />
to that <strong>of</strong> males, with the robust setae usually being<br />
more slender. As with most species <strong>of</strong> the <strong>Aegidae</strong> the<br />
shape and pattern and number <strong>of</strong> robust setae on the<br />
pleotelson and uropods is also diagnostic.<br />
55<br />
prey: Not known.<br />
distribution: Known only from the South Norfolk Basin,<br />
north <strong>of</strong> North Island, <strong>New</strong> <strong>Zealand</strong>.<br />
etymoLogy: Adapted from the Greek derkomai, meaning<br />
to see clearly (in allusion to the huge eyes).
Figure 95. Epulaega derkoma sp. nov. Holotype. A–D, pleopods –3, 5, respectively; E, uropod.<br />
Epulaega fracta (Hale, 940), comb. nov. (Figs 96–99)<br />
Aega fracta Hale, 1940: 296, fig. 4.– Bruce, Lew Ton & Poore,<br />
2002: 6 .<br />
Aega (Ramphion) fracta.– Brusca, 983: .<br />
materiaL examined: Holotype, ♂ ( 4.9 mm), <strong>of</strong>f the<br />
Tasmanian coast, Australia, coll. FIS Endeavour (AM<br />
E6747). Specimen may have been dried at some point<br />
as pin holes are evident.<br />
56<br />
Non-type material. ♂ ( 5.5 mm), <strong>of</strong>f Great Barrier<br />
Island, North Island, October 2004, from Hyperoglyphe<br />
antarctica, longline at ~500 m, coll. Steve Lowe (NIWA<br />
34806). ♂ ( 4.4 mm), Conway Rise, <strong>of</strong>f Kaikoura, 3 May<br />
987, in Symplectella, 400 m, coll. E. Barbarel (NMNZ<br />
Cr. 20 5). ♀ (non-ovig 5.5 mm), Conway Rise, <strong>of</strong>f<br />
Kaikoura, 2 September 984, in ‘organ pipe’ sponge, 400<br />
m, coll. Ted Forbes on F.V. Bar-K-Lin (NMNZ Cr.4970).<br />
Imm. ♂ ( 0.6 mm), manca (6.0 mm), J/ 6/28/84, inside
Figure 96. Epulaega fracta (Hale, 940). A, holotype AM E6747, remainder NMNZ Cr.9260. A, dorsal view, holotype;<br />
B, dorsal view, NMNZ Cr.9260; C, lateral view; D, head; E, frons; F, pleonites, lateral view; G, sternite 7 showing penial openings;<br />
pleotelson; H, spines, dorsal surface <strong>of</strong> pleotelson; I, antenna peduncle; J antennule; K, antenna flagellum, article 11.<br />
large orange sponge (NMNZ Cr. 200 ). ♂ ( 8.0 mm),<br />
<strong>New</strong> <strong>Zealand</strong>, stn J679 [station could not be traced]<br />
(NIWA 23764). ♂ ( 7.0 ), <strong>of</strong>f Great Barrier Island, North<br />
Island, January 2006, old longline gear at ~500 m, coll.<br />
Steve Lowe (NIWA 23765). Queensland: ♀ (ovig. 3.0<br />
mm), Marion Plateau, 22° 4. – 0. ’S 58°3 .7–29. ’E,<br />
303-333 m, 9 November 985, stn 0685/ 5/6, from<br />
Hexactinella, coll. Soela (NTM Cr 4926).<br />
57
Figure 97. Epulaega fracta (Hale, 940). NMNZ Cr.9260. A, mandible; B, mandible palp, article 3; C, maxillule apex; D, maxilla<br />
apex; E, maxilla; F, maxilliped; G; maxilliped palp article 5.<br />
Also examined: 24 specimens (5 mature adults) <strong>of</strong><br />
Aega monilis Barnard, 9 4, 3 °35’S, 6°30’E, 24 January<br />
995, 33 m, in sponge, coll. SAFRI Africana (SafM<br />
A43 7).<br />
type LoCaLity: ‘Off Tasmanian coast’ (Hale 940), southeastern<br />
Australia.<br />
desCription: Body 2.3 times as long as greatest width,<br />
dorsal surfaces sparsely punctate, widest at pereonite<br />
5, lateral margins subparallel. Rostral point ventrally<br />
directed. Eyes large, medially united, anterior clear field<br />
14% length <strong>of</strong> head, posterior clear field 44% length <strong>of</strong><br />
head; each eye made up <strong>of</strong> ~ 3 transverse rows <strong>of</strong> ommatidia,<br />
each row with ~7 ommatidia; eye colour black.<br />
Pereonite 1 and coxae 2–3 each with posteroventral angle<br />
rounded; pereonite 2 with median curved transverse<br />
nodulose ridge; posterior margins <strong>of</strong> pereonites with<br />
58<br />
small nodules. Coxae 5–7 with entire oblique carina;<br />
posterior margins convex, posterolateral angle blunt<br />
(more than 45°). Pleon with pleonite visible in dorsal<br />
view; pleonites with small nodules along posterior<br />
margin; pleonite 4 with posterolateral margins extending<br />
clearly beyond posterior margin <strong>of</strong> pleonite 5;<br />
pleonite 5 with posterolateral angles free, not overlapped<br />
by lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.7<br />
times as long as anterior width, dorsal surface without<br />
longitudinal carina, with short acute spines; lateral<br />
margins convex, smooth, posterior margin with distinct<br />
short median point, with 4–6 RS.<br />
Antennule peduncle articles 3 and 4 . times as<br />
long as combined lengths <strong>of</strong> articles and 2, article 3<br />
3.8 times as long as wide; flagellum with 12 articles,<br />
extending to posterior <strong>of</strong> pereonite . Antenna peduncle<br />
article 4 2. times as long as wide, 0.7 times as long<br />
as combined lengths <strong>of</strong> articles –3, inferior margin 0
Figure 98. Epulaega fracta (Hale, 940). NMNZ Cr.9260. A–D, pereopods , 2, and 7, respectively.<br />
plumose setae, and 0 short simple setae; article 5 .3<br />
times as long as article 4, 2.8 times as long as wide, inferior<br />
margin with 0 palmate setae, anterodistal angle<br />
with cluster <strong>of</strong> 8 short simple setae (and palmate);<br />
flagellum with 18 articles, extending to pereonite 4.<br />
Frontal lamina flat, as wide as long, diamond shaped,<br />
anterior margin acute, forming median angle, posterior<br />
margin forming narrow stem.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 23 distolateral setae (stiff),<br />
palp article 3 with 22 setae. Maxillule with 5 terminal<br />
RS ( large, 4 slender, plus 2 stubs). Maxilla mesial lobe<br />
with 3 RS ( straight, 2 biserrate); lateral lobe with 3 RS<br />
(recurved). Maxilliped endite with 0 apical setae (endite<br />
large); palp article 2 with 2 RS (curved, slender); article<br />
3 with 4 RS (all slender, weakly recurved); article<br />
4 with 4 hooked RS; article 5 partly fused to article<br />
4, small subcircular lobe, with RS (biserrate; plus<br />
simple seta).<br />
Pereopod 1 basis 3.0 as long as greatest width; ischium<br />
0.4 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 2 RS (one stub-like);<br />
merus inferior margin with 3 RS, set as two groups<br />
(<strong>of</strong> 2 and ), superior distal angle with 0 RS ( short<br />
simple seta); carpus 0.9 as long as merus, inferior<br />
margin with 0 RS; propodus .3 times as long as proximal<br />
width, inferior margin with RS (opposite base<br />
<strong>of</strong> dactylus), propodal palm simple, without blade<br />
or process, dactylus smoothly curved, .0 as long as<br />
propodus. Pereopod 2 ischium inferior margin with<br />
RS, superior distal margin with 2 RS; merus inferior<br />
margin with 5 RS, set as two groups, superior distal<br />
59
Figure 99. Epulaega fracta (Hale, 940). NMNZ Cr.9260. A–D, pleopods –3, 5 respectively; E, uropod; F, pleotelson posterior<br />
margin.<br />
margin with acute RS; carpus similar in size to that<br />
<strong>of</strong> pereopod , inferodistal angle with 0 RS. Pereopod 3<br />
similar to pereopod 2. Pereopods 5–7 inferior margins<br />
<strong>of</strong> ischium–carpus with short RS. Pereopod 6 similar to<br />
pereopod 7 (slightly larger). Pereopod 7 basis 2.8 times<br />
as long as greatest width, inferior margins with 8 palmate<br />
setae; ischium 0.4 as long as basis, inferior margin<br />
with 3 RS (set as and 2), superior distal angle with 4<br />
60<br />
RS, inferior distal angle with 4 RS; merus . as long<br />
as ischium, .9 times as long as wide, inferior margin<br />
with 4 RS (set as , and 2), superior distal angle with<br />
RS, inferior distal angle with 5 RS; carpus .2 as long<br />
as ischium, 2.3 times as long as wide, inferior margin<br />
with 3 RS (set as , and ), superior distal angle with<br />
4 RS (simple and biserrate setae), inferior distal angle<br />
with 7 RS; propodus .2 as long as ischium, 4.3 times
as long as wide, inferior margin with 3 RS (set as and<br />
2), superior distal angle with 2 slender setae, inferior<br />
distal angle with 3 RS.<br />
Penes medially united; penial process 0.4 times as<br />
long as basal width.<br />
Pleopod 1 exopod .4 times as long as wide, distally<br />
broadly rounded, lateral margin straight, mesial<br />
margin strongly convex, with PMS on distal half,<br />
with ~58 PMS; endopod .9 times as long as wide,<br />
distally subtruncate, lateral margin weakly concave,<br />
with PMS on distal margin only, mesial margin with<br />
PMS on distal one-third, endopod with ~40 PMS; peduncle<br />
.7 times as wide as long, mesial margin with<br />
8 coupling hooks. Pleopod 2 exopod with ~88 PMS,<br />
endopod with ~60 PMS; appendix masculina basally<br />
swollen, 0.7 times as long as endopod, distally acute.<br />
Pleopod 3 exopod with ~ 00 PMS, endopod with ~27<br />
PMS. Pleopod 4 exopod with ~90 PMS, endopod with<br />
~25 PMS. Pleopod 5 exopod with ~72 PMS. Exopods <strong>of</strong><br />
pleopods –3 each with distolateral margin not digitate;<br />
endopods <strong>of</strong> pleopods 3–5 each with distolateral<br />
point; pleopods 2–4 peduncle distolateral margin with<br />
prominent acute RS.<br />
Uropod peduncle posterior lobe about three-<br />
quarters as long as endopod. Uropod rami extending<br />
to pleotelson apex, apices acute. Endopod apically not<br />
bifid, lateral margin straight, without prominent ex-<br />
cision, proximal lateral margin with 0 RS, distal lateral<br />
margin with 2 RS, mesial margin sinuate, with 7 RS.<br />
Exopod not extending to end <strong>of</strong> endopod, 3.0 times as<br />
long as greatest width, apically not bifid; lateral margin<br />
weakly convex, with 9 RS; mesial margin straight,<br />
distally convex, with 4 RS.<br />
femaLe: More ovate than male; lacks the transverse<br />
ridge on pereonite 2; lacks dorsal nodules except on<br />
the pleotelson, where they are present though smaller<br />
than in the male.<br />
size: Adults measure between 4.4 and 8.0 mm; the<br />
single manca was 6.0 mm.<br />
Variation: <strong>The</strong> small number (4) <strong>of</strong> mature adult specimens<br />
means that it is not possible to precisely detail<br />
the variation. <strong>The</strong> pleotelson has from about 4 to 6 RS.<br />
<strong>The</strong> uropod endopod lateral margin has 2 distal RS,<br />
the mesial margin 6 or 7 RS for adults (5 in the manca);<br />
the exopod lateral margin has 9 or 0 RS (8 and 9 in<br />
the manca), the mesial margin 4 RS (3 in the manca).<br />
<strong>The</strong> RS on the merus <strong>of</strong> pereopods –3 appear to be<br />
constant but could not be readily discerned in the two<br />
smallest specimens.<br />
Body shape varies a little, with the adult male from<br />
<strong>New</strong> <strong>Zealand</strong> having subparallel lateral margins (2.4<br />
times as long as wide), while the male holotype and<br />
non-ovigerous female were both wider and more ovate<br />
(2.3 times as long as wide).<br />
6<br />
remarks: This is a distinctive species, males being readily<br />
recognised by the fine nodules along the posterior<br />
margins <strong>of</strong> the posterior pereonites and pleonites, presence<br />
<strong>of</strong> numerous small spines on the dorsal surface<br />
<strong>of</strong> the pleotelson and medially fused penial processes.<br />
Females can be recognised by the presence <strong>of</strong> reduced<br />
spines on the pleotelson and the characteristic setation<br />
and shape <strong>of</strong> the uropod rami and posterior margin <strong>of</strong><br />
the pleotelson.<br />
<strong>The</strong> shape <strong>of</strong> the eyes is unusual within the genus<br />
in that they are noticeably anterior in position (rather<br />
than ventral), appearing somewhat bulbous and almost<br />
entirely filling the anterior margin <strong>of</strong> the head in<br />
dorsal view. <strong>The</strong> propodus <strong>of</strong> pereopod is distinctive<br />
in having a prominent and acute robust seta opposing<br />
the base <strong>of</strong> the dactylus. In most <strong>Aegidae</strong> there is no<br />
such seta at this position.<br />
Epulaega monilis (Barnard, 9 4) is closely similar<br />
to Epulaega fracta, but can be distinguished by lacking<br />
spines on the dorsal surface <strong>of</strong> the pleotelson in<br />
both males and females, the nodules on the posterior<br />
margins <strong>of</strong> the pleonites are larger (particularly the<br />
median nodules) and extend to the anterior pereonites,<br />
and the uropodal exopod is as long as the endopod (a<br />
little shorter in E. fracta).<br />
prey: One specimen from Hyperoglyphe antarctica (Carmichael,<br />
8 9) [bluenose and matiri (<strong>New</strong> <strong>Zealand</strong>) or<br />
Antarctic butterfish, Centrolophidae].<br />
distribution: Southwestern Pacific, <strong>of</strong>f Tasmania and<br />
southern Queensland; <strong>of</strong>f Great Barrier Island, North<br />
Island and Kaikoura, South Island, both on the east<br />
coast <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>; at a recorded depth <strong>of</strong> 400–500<br />
metres.<br />
Genus Rocinela Leach, 8 8<br />
Rocinela Leach, 8 8: 349.– Desmarest, 825:304.– Milne<br />
Edwards, 840: 243.– Dana, 852: 304*.– Bate &<br />
Westwood, 867: 289.– Schioedte & Meinert: 879b: 380.–<br />
Gerstaecker, 882: 227.– Haswell, 882: 285.– Stebbing,<br />
893: 348; 905: 23.– Sars, 897: 65.– Richardson, 898:<br />
8; 905a: 90.– Barnard, 9 4: 368; 936: 59.– Hale,<br />
925: 82.– Menzies, 962: 8.– Menzies & Glynn,<br />
968: 45.– Menzies & George, 972: 2.– Kensley, 978:<br />
59.– Kussakin, 979: 25 .– Menzies & Kruczynski, 983:<br />
62.– Brusca, 980: 229.– Bruce, 983: 778.– Brusca &<br />
Iverson, 985: 42.– Brusca & France, 992: 236.– Kensley &<br />
Schotte, 989: 9.– Bruce, Lew Ton & Poore, 2002: 63.<br />
Acherusia Lucas, 849: 78.– Dana, 852: 304* (type species<br />
Acherusia dumerilii Lucas, 849).<br />
Acherousia.– Schioedte & Meinert, 879b: 380 (lapsus).<br />
Rocinella.– Bate, 878 : 65 (lapsus).<br />
Not Rocinela.– Bovallius 885: 4 (= Syscenus Harger, 880).<br />
type speCies: Rocinela danmoniensis Leach, 8 8, by<br />
monotypy.<br />
* <strong>The</strong>re is a pagination error in this publication, with page<br />
304 printed as 204.
diagnosis: Body weakly dorsally vaulted. Head with<br />
posterolateral margins contained by anterolateral<br />
angles <strong>of</strong> pleonite ; rostral point blunt, overriding<br />
antenna and antennule peduncles; eyes present, <strong>of</strong>ten<br />
large, sometimes united, occupying more than 50%<br />
width <strong>of</strong> the head. Pleonite not abruptly narrower<br />
than pereonite 7. Frontal lamina present, small, narrow.<br />
Antennule much shorter than antenna, usually shorter<br />
than antenna peduncle. Maxilliped palp 3-articled;<br />
endite present. Pleopods 3–4 endopods without PMS.<br />
Uropodal peduncle mesial margin produced; rami<br />
lamellar, plane <strong>of</strong> exopod at strongly oblique angle to<br />
plane <strong>of</strong> endopod; pleotelson posteriorly rounded.<br />
desCription: Body moderately vaulted, about 2 to 4<br />
times as long as wide. Head with eyes, <strong>of</strong>ten large, may<br />
meet at midpoint; anterior margin with broad median<br />
(rostral) point. Coxae <strong>of</strong> pereonites 4–7 longer than<br />
respective segment, posteriorly produced. Pleon not<br />
abruptly narrower than pereon; pleonites all visible,<br />
not posteriorly widest, pleonites 2–4 with free lateral<br />
margins, pleonite 5 laterally overlapped by pleonite<br />
4; pleonites 3–5 laterally produced to an acute point.<br />
Pleotelson large, about as long or longer pleon, usually<br />
with PMS and RS. Pleonal sternite absent.<br />
Antenna peduncle articles 4 and 5 usually with<br />
long setae.<br />
Mandible with uni- or bicuspid incisor; molar process<br />
present, conspicuous; two scaled lobes present at<br />
base <strong>of</strong> incisor. Maxillule with 5–8 flat terminal robust<br />
setae; mesial lobe reduced or absent. Maxilla lateral<br />
lobe with 2–5 terminal hooked RS, mesial with –3<br />
straight or hooked RS. Maxilliped 3-articled, article 2<br />
with 2 or 3 stout recurved RS and article 3 each with<br />
or 2 stout recurved RS; endite present.<br />
Pleopods 3–5 endopods smaller than exopods,<br />
usually with thickened ridge; coupling setae present<br />
on peduncles <strong>of</strong> pleopods –4. Pleopods not extending<br />
beyond lateral margins <strong>of</strong> pleon.<br />
remarks: Rocinela is rather uniform in appearance,<br />
typically flat-bodied, relatively wide with a prominent<br />
wide and flat rostrum (or this could be interpreted as<br />
the rostrum absent and the anterior part <strong>of</strong> the head<br />
being produced and forming a process). Most species<br />
have large or very large eyes, in a few species the eyes<br />
meeting medially. Many species have a flat lobe or<br />
blade on the palm <strong>of</strong> pereopod –3 propodus, this blade<br />
always being provided with robust setae. Rocinela appears<br />
to have highest diversity in high latitudes with<br />
only 2 <strong>of</strong> the 42 species occurring within the tropics.<br />
<strong>The</strong> high-latitude diversity <strong>of</strong> the genus is maintained<br />
by the nine species present in <strong>New</strong> <strong>Zealand</strong> waters.<br />
<strong>The</strong>se figures are probably due to under reporting as<br />
museum collections in Australia and those held at the<br />
Muséum national d’Histoire naturelle in Paris do have<br />
62<br />
significant numbers <strong>of</strong> undescribed species, notably<br />
from the tropical western Pacific region.<br />
Rocinela is one <strong>of</strong> the very few isopod genera,<br />
other than some cirolanids, known to attack humans<br />
(Garzon-Ferreira 990).<br />
Key to the new <strong>Zealand</strong> species <strong>of</strong> RocinelA<br />
. Pereopod propodus slender, 4.4 times as long as<br />
proximal width, without distinct propodal blade;<br />
eyes small, separated by 40% head width ..........2<br />
– Pereopod propodus robust, less than 2.0 times<br />
as long as proximal width, with distinct propodal<br />
blade or lobe; eyes large separated by less than<br />
30% head width .....................................................3<br />
2. Rostrum subtruncate, anterior margin <strong>of</strong> the head<br />
‘stepped’; pereopod propodal palm with 2 small<br />
distal RS, with small distal lobe; dactylus weakly<br />
curved, 0.8 times as long as propodus.. ................<br />
......................................... Rocinela leptopus (p. 74)<br />
– Rostrum smoothly narrowed, rounded; pereopod<br />
propodal palm with minute RS, without distal<br />
lobe; dactylus distally curved, .0 times as long<br />
as propodus .......................Rocinela runga (p. 89)<br />
3. Pereopod propodal blade wide, approximately<br />
as long (0.9– . ) as palm, with more than 8 marginal<br />
RS, ...................................................................4<br />
– Pereopod propodal blade narrow, 0.5–0.7 as<br />
long as propodal palm, with 6 or less marginal<br />
RS ..............................................................................5<br />
4. Rostrum tri-cornered; eyes moderate, separated<br />
by about 3 % width <strong>of</strong> head; pereopod propodal<br />
blade with 8 or 9 marginal RS; mesial surface with<br />
numerous stiff simple setae ....................................<br />
.......................................... Rocinela garricki (p. 69)<br />
– Rostrum evenly narrowing to subtruncate anterior<br />
margin; eyes large, separated by about 2.5% width<br />
<strong>of</strong> head; pereopod propodal blade with 2 or 3<br />
marginal RS; mesial surface with single simple<br />
seta .......................................Rocinela pakari (p. 78)<br />
5. Pereopod blade about 0.7 as long as propodal<br />
palm, with 5 or 6 marginal RS, with abundant<br />
slender setae on mesial surface ..............................<br />
........................................... Rocinela satagia (p. 93)<br />
– Pereopod blade about 0.5 as long as propodal<br />
palm, with less than 6 marginal RS, without<br />
abundant slender setae on mesial surface ......... 6<br />
6. Rostrum subtruncate; eyes large, separated by<br />
about 4% width <strong>of</strong> head; pereopod propodal<br />
blade with 4 marginal RS ........................................<br />
.............................................Rocinela bonita (p. 63)<br />
– Rostrum anteriorly truncate, turned upwards;<br />
eyes moderate, separated by about 28% width <strong>of</strong><br />
head; pereopod propodal blade with 3 marginal<br />
RS ........................................ Rocinela resima (p. 84)
Rocinela bonita sp. nov. (Figs 00– 04)<br />
materiaL examined: Holotype, ♀ (3 mm, non-ovig.),<br />
Bounty Trough, 44°26.89’S, 74°54.79’E, 25 October<br />
979, Stn S 44, 676 m, epibenthic sled (NIWA 2388 ).<br />
Paratypes. 3 ♂ (35, 22 mm and large male pleon and<br />
pleotelson), ♀ (22 mm, non-ovig.), 7 mancas ( –22<br />
mm, further 8 not measured), same data as holotype<br />
(NIWA 23882).<br />
desCription: Body 2.0 times as long as greatest width,<br />
dorsal surfaces smooth and polished in appearance,<br />
widest at pereonite 5, lateral margins ovate. Rostral<br />
point anteriorly subtruncate. Eyes not medially united,<br />
separated by about 4% width <strong>of</strong> head; each eye made<br />
up <strong>of</strong> ~ 7 transverse rows <strong>of</strong> ommatidia, each row<br />
with ~ 0 ommatidia; eye colour dark brown. Pereonite<br />
1 and coxae 2–3 each with posteroventral angle<br />
rounded; coxae 5–7 with incomplete oblique carina<br />
(on coxae 5–7). Pleon with pleonite largely concealed<br />
by pereonite 7; pleonite 4 with posterolateral margins<br />
extending clearly beyond posterior margin <strong>of</strong> pleonite<br />
5; pleonite 5 with posterolateral angles free, not overlapped<br />
by lateral margins <strong>of</strong> pleonite 4. Pleotelson 0.8<br />
times as long as anterior width, anterior dorsal surface<br />
without 2 sub-medial depressions, dorsal surface with<br />
short setae; lateral margins convex, posterior margin<br />
evenly rounded, with 6 RS.<br />
Antennule peduncle article 3 .0 times as long as<br />
combined lengths <strong>of</strong> articles and 2, 3.0 as long as wide;<br />
flagellum with 6 articles, extending to posterior margin<br />
<strong>of</strong> eye. Antenna peduncle article 3 .5 times as long as<br />
article 2, . times as long as wide; article 4 .5 times<br />
as long as article 3, .6 times as long as wide, inferior<br />
margin with 2 plumose setae, and 8 simple setae; article<br />
5 .3 times as long as article 4, 2.9 times as long as wide,<br />
inferior margin with 7 setae (2 plumose), anterodistal<br />
angle with cluster <strong>of</strong> 1 short simple seta; flagellum with<br />
7 articles, extending to posterior <strong>of</strong> pereonite .<br />
Frontal lamina longer than greatest width, anteriorly<br />
acute.<br />
Mandible molar process distinct flat lobe; palp article<br />
2 with 4 short marginal distolateral setae and 2 long<br />
distolateral setae; palp article 3 with 30 setae. Maxillule<br />
with 6 RS ( large, 5 slender, serrate). Maxilla mesial<br />
lobe with 2 RS; lateral lobe with 2 RS. Maxilliped palp<br />
article distomesial angle with 3 RS (short; 2 curved,<br />
straight); article 2 with 3 hooked RS; article 3 with 2<br />
hooked RS.<br />
Pereopod 1 basis 2.7 times as long as greatest width;<br />
ischium 2.7 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 7 setae ( robust seta);<br />
merus inferior margin with 3 RS (set as +2), set as two<br />
groups, superior distal angle with 6 setae; carpus 0.7<br />
as long as merus, inferior margin with RS (minute);<br />
propodus .6 times as long as proximal width, propodal<br />
palm with blade, propodal blade 0.5 times as wide as<br />
palm, inferior margin with 4 RS; dactylus 2.0 times as<br />
long as propodus. Pereopods 2 and 3 similar to pereopod<br />
(but RS on merus longer). Pereopod 6 similar to pereopod<br />
7. Pereopod 7 basis 3.9 times as long as greatest<br />
width, inferior margins with 0 palmate setae; ischium<br />
0.7 as long as basis, inferior margin with 7 RS (set as ,<br />
3, 2 and ), superior distal angle with 5 RS (and 2 setae),<br />
inferior distal angle with 3 RS; merus 0.6 times as long<br />
as ischium, 2.3 times as long as wide, inferior margin<br />
with 3 RS (set as and 2), superior distal angle with 3<br />
RS, inferior distal angle with 4 RS; carpus 0.6 as long<br />
as ischium, 2.8 times as long as wide, inferior margin<br />
with 3 RS (set as and 2), superior distal angle with 2<br />
RS, inferior distal angle with 6 RS; propodus 0.5 as long<br />
as ischium, 4.0 as long as wide, inferior margin with<br />
2 RS (set singly), superior distal angle with 4 slender<br />
setae, inferior distal angle with RS.<br />
Pleopod 1 exopod .9 times as long as wide, lateral<br />
margin straight, mesial margin weakly convex, with<br />
PMS from distal two-thirds; endopod 2.6 times as long<br />
as wide, lateral margin weakly concave, with PMS<br />
from on distal margin only, mesial margin with PMS<br />
from distal one-third; peduncle mesial margin with 6<br />
coupling hooks. Pleopods 2–5 peduncle distolateral<br />
margin with acute RS.<br />
Uropod peduncle ventrolateral margin with 0 RS,<br />
posterior lobe about one-third as long as endopod. Exopod<br />
at angle <strong>of</strong> about 35° to endopod, rami extending<br />
to pleotelson apex, marginal setae in two tiers. Endopod<br />
lateral margin weakly sinuate, lateral margin with 8<br />
RS, mesial margin weakly convex, with 4 RS. Exopod<br />
not extending to end <strong>of</strong> endopod, 2.4 times as long as<br />
greatest width; lateral margin weakly convex, with 2<br />
RS; mesial margin sinuate, proximally concave, with<br />
RS; distal margin rounded.<br />
maLe: Pleopod 2 appendix masculina with straight margins,<br />
0.9 times as long as endopod, distally narrowly<br />
rounded. Otherwise similar to female, but with a more<br />
elongate body shape (as do immature specimens and<br />
mancas); the rostrum is more strongly produced than<br />
in the female, the robust setae on the pereopod palm<br />
<strong>of</strong> pereopods –3 are more slender and acute, and the<br />
lateral margins <strong>of</strong> the uropodal rami are more densely<br />
setose.<br />
size: Males 22 and 35 mm; females 22 and 3 mm; mancas<br />
–22 mm (mean = 8.3 mm, n = 8); the large pleon<br />
and pleotelson indicate that this species will reach a<br />
greater size than indicated here.<br />
Variation: Robust setae: Pleotelson (n = 2) RS 3– 7,<br />
with 4, 5 and 6 each at 25%. Uropod exopod (n = 24)<br />
mesial margin with (7 %) most frequent, 0 and 2 each<br />
occurring three times; lateral margin – 4 with 2<br />
63
Figure 100. Rocinela bonita sp. nov. Holotype; C, F, male paratype 35 mm, others as indicated. A, dorsal view; B, lateral view;<br />
C, dorsal view, male; D, head; E, frons; F, head, paratype; G, head, manca, 20 mm; H, coxa 7; I, pleonites 4 and 5, lateral margins;<br />
J, sternite 7; K, penial processes.<br />
64
Figure 101. Rocinela bonita sp. nov. Male paratype 35 mm, others as indicated. A, mandible; B, antenna; C, antennule;<br />
D, mandible palp article 3; E, maxillule; F, maxillule apex; G, maxillule apex, non-ovig female, 22 mm; H, maxilla apex;<br />
I, maxilla; J, maxilla apex, non-ovig female, 22 mm; K, maxilliped; L, maxilliped palp; M, maxilliped palp articles 2 and 3,<br />
non-ovig female, 22 mm.<br />
65
Figure 102. Rocinela bonita sp. nov. Holotype, others as indicated. A–E, pereopods –3, 6, and 7, respectively;<br />
F, pereopod ischium, mesial surface; G, distal margin <strong>of</strong> carpus and propodus, mesial surface; H, I, male 35 mm: H, pereopod ;<br />
I, pereopod 2.<br />
66
Figure 103. Rocinela bonita sp. nov. Male paratype 35 mm. A–E, pleopods –5 respectively; F, pleopod peduncle mesial<br />
and lateral margins.<br />
(38%), 3 (33%) and 4 (20%) most frequent. Uropod<br />
endopod mesial margin varied from 2 to 5, with 4 (76%)<br />
the most frequent, 3 and 5 each occurring twice and 2<br />
once; lateral margin with 6–8, with 7 (48%) and 8 (43%)<br />
most frequent, 6 occurring twice.<br />
<strong>The</strong> setation <strong>of</strong> the pereopod palm is highly consistent<br />
with 4 robust setae on pereopods –3 with only<br />
one exception with 3 robust setae; no specimen had<br />
more than four robust setae on the palm. <strong>The</strong> robust<br />
setae count on the inferior margin <strong>of</strong> the merus was<br />
consistently +2, although some <strong>of</strong> these were missing<br />
and there may be some variation.<br />
<strong>The</strong>re is no discernable difference in the number <strong>of</strong><br />
robust setae between males and females, nor in relation<br />
to size—the smallest male ( 5.5 mm) and female<br />
( 6.5 mm) had similar counts for robust setae: pleotelson<br />
RS ( ), pereopods –3 (8 and 9) and uropods<br />
(endopod mesial with 2 and 3, lateral with 3).<br />
Eye size varies with size, small specimens having<br />
proportionally larger eyes than adult specimens; the<br />
eyes are separated by 2.5%, 5% and 24% in the manca<br />
(Fig. 00G), female holotype (Fig. 00D) and adult male<br />
(Fig. 00F) respectively.<br />
remarks: Rocinela bonita sp. nov. can be identified by<br />
the large but well-separated eyes, relatively narrow<br />
rostrum, narrow sub-rectangular blade on the palm <strong>of</strong><br />
pereopods –3 that is consistently provided with four<br />
robust setae (stout in juveniles and females, slender in<br />
the mature male), evenly rounded pleotelson, pleotel-<br />
67
Figure 104. Rocinela bonita sp. nov. A, E, holotype, remainder male paratype. A, pleotelson and uropod; B, uropod; C, uropod<br />
endopod ventral view; D, uropod endopod, apex; E, pleotelson posterior margin.<br />
son anterior dorsal surface without submedian depressions,<br />
posterior margin <strong>of</strong> the pleotelson with 4– 6<br />
robust setae, uropodal endopod extending posterior<br />
to the exopod, broadly rounded uropod exopod and<br />
the uropod rami not extending significantly beyond<br />
the posterior margin <strong>of</strong> the pleotelson. In females the<br />
rostrum is anteriorly rounded, in the male it is more<br />
strongly produced and anteriorly subtruncate.<br />
Rocinela resima sp. nov. is the most similar species<br />
in <strong>New</strong> <strong>Zealand</strong> waters. R. bonita can be distinguished<br />
by pereopods –3 having more strongly produced propodal<br />
blade with four robust setae, longer and more<br />
slender robust setae on the merus, and the uropodal<br />
endopod lateral margin being sinuate with a narrowly<br />
68<br />
rounded distal point, whereas in R. resima the uropodal<br />
endopod lateral margin is evenly convex, the distal<br />
margin being broadly rounded.<br />
Rocinela juvenalis Menzies & George, 972 appears<br />
similar, but that species differs in having the uropodal<br />
exopod exceeding the posterior <strong>of</strong> the endopod, pereopod<br />
propodus palm being wider and in being much<br />
smaller in size (the holotype and presumably adult<br />
female <strong>of</strong> R. juvenalis measured mm, the mancas <strong>of</strong><br />
R. bonita sp. nov. measure –22 mm).<br />
Rocinela modesta Hansen, 897 has a similar pereopod<br />
morphology (Brusca & France 992) but has an<br />
anteriorly rounded rostrum, smaller eyes, four robust<br />
setae on the inferior margin <strong>of</strong> the merus <strong>of</strong> pereopod
3, more strongly produced uropod peduncle posterior<br />
lobe (half as long as endopod versus one-third as long<br />
in R. bonita) and an evenly convex uropodal endopod<br />
lateral margin (sinuate in R. bonita).<br />
Rocinela cornuta Richardson, 898 is a poorly known<br />
species, superficially similar to both the present species<br />
and to Rocinela hawaiiensis Richardson, 903. Brusca and<br />
France ( 992) illustrated the dorsal view for the species<br />
but descriptive data otherwise rests with the original<br />
description. R. bonita differs in having a less ovate<br />
body shape and the propodal blade <strong>of</strong> pereopods –3<br />
provided with four (rather than three) robust setae.<br />
Rocinela major Brocchi, 877, is the only species<br />
that could not be specifically excluded as no figures<br />
exist and the description is not adequate by modern<br />
standards <strong>of</strong> species description. <strong>The</strong> species should<br />
be regarded as species inquirenda as, in addition to<br />
the lack <strong>of</strong> descriptive data, the location <strong>of</strong> the types<br />
is uncertain. If there is found to be no type material<br />
the species will have to be relegated to nomen dubium.<br />
Kensley ( 976) considered it as probably the same species<br />
as Epulaega monilis Barnard, 9 4. All other species<br />
are excluded by varying combinations <strong>of</strong> differences<br />
among the differential characters.<br />
prey: Not known.<br />
distribution: Known only from the type locality,<br />
Bounty Trough, eastwards from the mid-coast <strong>of</strong> South<br />
Island; 676 metres.<br />
etymoLogy: From the Latin origin for bounty (as in the<br />
Bounty Trough) — bonitas, meaning good, plentiful.<br />
Rocinela garricki Hurley, 957 (Figs 05– 09)<br />
Rocinela garricki Hurley, 1957: 11, figs 39–49; 1961: 268.– Hicks<br />
et al., 99 : 6.<br />
materiaL examined: Holotype: ♂ (~ 5 mm, previously<br />
dissected, pleotelson missing), Cook Strait, 4 °3 .5’S,<br />
74°48.0’E, 9 January 956, 28– 46 m, beam trawl,<br />
station BOL, Vuz 43 (NMNZ Cr.365 ).<br />
Other material: ♂ ( 5.5 mm, dissected), Camp Bay,<br />
Endeavour Inlet, Queen Charlotte Sound, South Island,<br />
4 °08’S, 74°08.45’E, 0 May 967, <strong>of</strong>f jetty, stn.<br />
Z 5 3, coll. Maria van Dooren (NIWA 23855). ♀ (22<br />
mm, ovig.; poor condition, dried out at some point),<br />
southern Bounty Trough, 46.0°S, 70.72°E, 75 m, 8 October<br />
962, NIWA stn B568 (NIWA 23854). ♀ (~22 mm,<br />
broken, poor condition), <strong>of</strong>f northeastern South Island,<br />
42.7533°S, 73.50 7°E, 4 November 979, canyon coral,<br />
79 m (NIWA 23849). ♀ (non-ovig. 20, 9.5, 6.5 mm),<br />
<strong>New</strong> <strong>Zealand</strong>, without locality, Z2, ex groper, 22F,<br />
/63 (poor condition, possibly dried or in formalin<br />
for a long time) (NIWA 23852). ♂ ( 9.5 mm), Dunedin,<br />
South Island, 2 January 957, wharves, at night light,<br />
coll. R.K. Dell & J. Moreland (NMNZ Cr. 2004). Manca<br />
(5.5 mm), Cape Turakirae, 8 June 966, 42 m (NMNZ<br />
Cr. 2006).<br />
Not measured: ♀ (ovig.), <strong>New</strong> <strong>Zealand</strong>, no locality,<br />
Z2*, in poor condition (NIWA 23853). ♂ (~23 mm,<br />
pleotelson damaged), midway between Cape Jackson<br />
& Mana Island, North Island (Cook Strait), 4 °02’S,<br />
74°33’E, 6 March 976, 256– 86 m, RV Acheron (NMNZ<br />
Cr. 2005). ♂, <strong>of</strong>f Kaikoura, South Island, 42.4384°S,<br />
74.7600°E, 20 June 96 , 00 m (NIWA 23848). ♀<br />
(ovig., very damaged), south <strong>of</strong> <strong>The</strong> Snares, 48.0033°S,<br />
66.9500°E, 2 October 962, 55 m (NIWA 23847).<br />
♀ (ovig., poor condition), Chatham Rise, 44°30’S,<br />
76°00’W, 7 October 964, 92 m (NIWA 23850). ♂<br />
(poor condition) ( 6.5 mm), <strong>New</strong> <strong>Zealand</strong>, no locality<br />
data, stn Z6 4 (NIWA 2385 ).<br />
desCription: Body 2.3 times as long as greatest width,<br />
dorsal surfaces smooth, widest at pereonite 5, lateral<br />
margins ovate. Rostrum basally expanded, anteriorly<br />
rounded and tri-cornered. Eyes not medially united,<br />
separated by about 3 % width <strong>of</strong> head; each eye made<br />
up <strong>of</strong> ~ transverse rows <strong>of</strong> ommatidia, each row<br />
with ~8 ommatidia; eye colour pale brown. Coxae 2–3<br />
each with posteroventral angle rounded; 5–7 without<br />
oblique carina. Pleon with pleonite visible in dorsal<br />
view; pleonite 4 with posterolateral margins extending<br />
clearly beyond posterior margin <strong>of</strong> pleonite 5; pleonite<br />
5 with posterolateral angles rounded. Pleotelson 0.9<br />
times as long as anterior width, anterior dorsal surface<br />
without 2 sub-median depressions, dorsal surface with<br />
short setae (posteriorly); lateral margins convex, posterior<br />
margin evenly rounded, with RS.<br />
Antennule peduncle article 3 0.7 times as long as<br />
combined lengths <strong>of</strong> articles and 2, 2.9 times as long as<br />
wide; flagellum with 6 articles (articles 1 and 2 longest),<br />
extending to anterior <strong>of</strong> pereonite . Antenna peduncle<br />
article 3 .4 times as long as article 2, .2 times as long<br />
as wide; article 4 .3 times as long as article 3, .6 times<br />
as long as wide, inferior margin with 0 plumose setae,<br />
and 2 simple setae (stiff); article 5 .5 times as long as<br />
article 4, 2.8 times as long as wide, inferior margin with<br />
seta (palmate), anterodistal angle with cluster <strong>of</strong> 3<br />
short simple setae; flagellum with 11 articles, extending<br />
to posterior <strong>of</strong> pereonite 2.<br />
Frontal lamina longer than greatest width, anteriorly<br />
acute.<br />
Mandible molar process distinct flat lobe; palp article<br />
2 with 10 marginal distolateral setae (finely biserrate),<br />
69<br />
* Station ‘Z2’ (= Z0002) lists eight lots, mostly <strong>Aegidae</strong>,<br />
according to coordinates from the Fiji region. One lot<br />
includes an unpublished manuscript name <strong>of</strong> ‘timaruensis’.<br />
<strong>The</strong> data are clearly wrong for the material and the<br />
material is regarded as being <strong>New</strong> <strong>Zealand</strong>, no locality.
Figure 105. Rocinela garricki Hurley, 957. Holotype. A, dorsal view; B, head; dorsal view; C, pleonites 3 and 5, lateral margin;<br />
D, pereopod , in situ; E, pereopod 2, in situ.<br />
and 3 long distolateral setae; palp article 3 with 5 setae<br />
(terminal 2 short, conical). Maxillule with 5 RS ( large;<br />
4 slender, <strong>of</strong> which 2 weakly serrate). Maxilla mesial<br />
lobe with hooked RS; lateral lobe with 2 hooked RS.<br />
Maxilliped palp article distomesial angle with RS<br />
(straight); article 2 with 3 hooked RS; article 3 with<br />
hooked RS.<br />
Pereopod 1 basis 2.3 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 5 setae ( robust seta);<br />
merus inferior margin with 3 RS (set as + 2), set as two<br />
rows, superior distal angle with 6 setae (all stiff); carpus<br />
0.7 times as long as merus, inferior margin with RS;<br />
propodus .6 times as long as proximal width, propodal<br />
palm with blade, propodal blade 0.9 times as wide as<br />
palm, with numerous setae, inferior margin with 8 RS;<br />
dactylus .6 times as long as propodus. Pereopods 2 and<br />
3 similar to pereopod (RS on merus longer). Pereopod<br />
70<br />
6 similar to pereopod 7. Pereopod 7 basis 4.0 as long as<br />
greatest width, inferior margins with 2 palmate setae;<br />
ischium 0.8 as long as basis, inferior margin with 6 RS<br />
(set singly), superior distal angle with 4 RS, inferior<br />
distal angle with 7 RS; merus 0.5 times as long as ischium,<br />
2. times as long as wide, inferior margin with<br />
RS, superior distal angle with 5 RS, inferior distal<br />
angle with 6 RS; carpus 0.6 times as long as ischium,<br />
2.9 times as long as wide, inferior margin with 2 RS (set<br />
singly), superior distal angle with 7 RS, inferior distal<br />
angle with 2 RS; propodus 0.5 as long as ischium, 3.9<br />
times as long as wide, inferior margin with 3 RS (set<br />
singly), superior distal angle with 3 slender setae (<br />
palmate), inferior distal angle with 3 RS.<br />
Penes opening flush with surface <strong>of</strong> sternite 7.<br />
Pleopod 1 exopod 2.6 times as long as wide, lateral<br />
margin weakly convex, with PMS on distal one-third;<br />
endopod 2.6 times as long as wide, lateral margin
Figure 106. Rocinela garricki Hurley, 957. Male, 5.5 mm, Camp Bay, NIWA 23855. A, dorsal view; B, lateral view;<br />
C, pleonites –5, lateral margin; D, head, dorsal view; E, antennule; F, antenna peduncle.<br />
straight, with PMS on distal margin only, mesial margin<br />
with PMS on distal two-thirds; peduncle mesial<br />
margin with 6 coupling hooks. Pleopod 2 appendix<br />
masculina with straight margins, 0.5 times as long as<br />
endopod (basally fused), distally bluntly rounded.<br />
Pleopods 2–5 peduncle distolateral margin without<br />
acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about one-third as long as endopod. Exopod<br />
at angle <strong>of</strong> about 35° to endopod, rami extending<br />
to pleotelson apex, marginal setae in two tiers. Endopod<br />
lateral margin weakly convex, lateral margin with 3 RS,<br />
mesial margin straight and distally rounded, with 3 RS.<br />
Exopod not extending to end <strong>of</strong> endopod, 2.4 times as<br />
long as greatest width; lateral margin weakly convex<br />
(weakly serrate), with 8 RS; mesial margin convex, with<br />
0 RS; distal margin with distinct distal point.<br />
7<br />
size: Males from 5.5 to 9.5 mm; ovigerous females<br />
6.5–22.0 mm (mean 20 mm, n = 5); single manca 5.5<br />
mm.<br />
femaLe: Generally similar to the male, with the rostrum<br />
less strongly produced.<br />
Variation: <strong>The</strong> figures here include the holotype, therefore<br />
the uropod and pleotelson details are taken from<br />
the original description. Pleotelson (n = 8) RS 9– 2, with<br />
0 and most frequent, each at 38%. Uropod exopod<br />
(n = 4) mesial margin without RS in all specimens;<br />
lateral margin 7–9 RS with 8 (57%) most frequent, 7<br />
occurring 3 times and 9 once. Uropod endopod mesial<br />
margin (n = 3) varied from 0–5 to with 2 (3 %) and 3<br />
(39%) the most frequent, 0 occurring twice, 4 and 5 each<br />
occurring once; lateral margin (n = 4) with 2–5 with
Figure 107. Rocinela garricki Hurley, 957. Male, 5.5 mm, Camp Bay, NIWA 23855. A, mandible; B, mandible molar and<br />
incisor; C, mandible palp article 3; D, maxillule apex; E, maxillule; F, maxilla; G, maxilla apex; H, maxilliped; I, maxilliped<br />
palp articles 2 and 3; scales, mesial margin palp article ; K, frons.<br />
3 (36%) and 4 (43%) most frequent, 2 and 5 occurring<br />
once and twice respectively.<br />
<strong>The</strong> setation <strong>of</strong> the palms <strong>of</strong> pereopods –3 is highly<br />
consistent with 8 RS being the most frequent; pereopod<br />
palm (n = 4) with 8 (60%) or 9 (40%) RS, pereopod 2<br />
palm (n = 4) with 7 (27%) or 8 (73%) RS and pereopod<br />
3 (n = 3) palm 8 (93%), 7 RS occurring once. <strong>The</strong> RS<br />
on the inferior margin <strong>of</strong> the merus was consistently<br />
+2, although some <strong>of</strong> these were missing and there<br />
may be some variation.<br />
<strong>The</strong>re is no discernable difference in the number <strong>of</strong><br />
robust setae between males and females, nor in relation<br />
to size—the smallest male ( 5.5 mm) and female<br />
( 6.5 mm) had similar counts for robust setae: pleotelson<br />
RS ( ), pereopods –3 (8 and 9) and uropods<br />
72<br />
(endopod mesial with 2 and 3, lateral with 3 and 4,<br />
exopod mesial without, lateral 8). <strong>The</strong> characteristic<br />
shape <strong>of</strong> the rostrum is only evident in larger presumably<br />
mature specimens.<br />
remarks: A combination <strong>of</strong> characters serves to readily<br />
identify Rocinela garricki: the eyes are relatively widely<br />
separated, mature specimens have a distinctly tri-<br />
cornered rostrum, the anterior margin <strong>of</strong> which is <strong>of</strong>ten<br />
bent ventrally, the anterior pereopods usually have 8<br />
or 9 robust setae, the propodal blade is provided with<br />
numerous stiff simple setae (in contrast to the more<br />
usual single seta), the uropodal exopod has 8 robust<br />
setae on the lateral margin, none on the mesial margin,<br />
the uropodal endopod is relatively broad with 2 or 3
Figure 108. Rocinela garricki Hurley, 957. Male, 5.5 mm, Camp Bay, NIWA 23855. A–C, pereopods , 2 and 7 respectively;<br />
D, pereopod 7, distal margin <strong>of</strong> carpus, mesial RS; E, pereopod 7, distal margin <strong>of</strong> carpus, lateral RS.<br />
setae on the mesial margin, 3 or 4 setae on the lateral<br />
margin and the posterior margin <strong>of</strong> the pleotelson usually<br />
has 0 or robust setae. In addition, in mature<br />
specimens the anterior part <strong>of</strong> the head is depressed<br />
and there is an oblique longitudinal ridge that runs<br />
along the anteromesial margin <strong>of</strong> the eye.<br />
<strong>The</strong> most similar species is the potentially sympatric<br />
Rocinela satagia sp. nov., which has in common with<br />
R. garricki a tri-cornered rostrum and the blade <strong>of</strong> the<br />
anterior pereopods with numerous setae. Several differences<br />
allow separation <strong>of</strong> the two species, Rocinela<br />
satagia having eye-ridges on the posteromesial part <strong>of</strong><br />
the eyes, the eyes being close-set, the pereopod blades<br />
are smaller with no more than 6 robust setae and a<br />
narrower uropodal endopod.<br />
One large specimen, in poor condition, from ‘North<br />
Otago’ (a 32 mm non-ovig. female; NMNZ Cr.4966)<br />
is provisionally identified as R. garricki, but excluded<br />
from the material examined as it is in poor condition<br />
with most pereopods incomplete.<br />
Hicks et al. ( 99 ) listed one syntype (NMNZ<br />
Cr.365 ) held at Te Papa. Hurley ( 957) examined<br />
only one specimen and, as the Victoria University<br />
label states ‘type’, that specimen is here regarded as<br />
the holotype. <strong>The</strong> specimen itself is heavily dissected,<br />
with the pleotelson and pereopods from one side all<br />
missing. <strong>The</strong> five slides mentioned in Hurley’s (1957)<br />
description have not been located and are presumed<br />
lost.<br />
distribution: Cook Strait and <strong>of</strong>f the eastern coast <strong>of</strong><br />
South Island to Dunedin; most locations are inshore<br />
and shallow, with recorded depths from the surface<br />
(at a night light) to 256 metres.<br />
etymoLogy: Named for Mr J. A. Garrick, presumably a<br />
productive collector at that time (Hurley 957).<br />
73
Figure 109. Rocinela garricki Hurley, 957. Male, 5.5 mm, Camp Bay, NIWA 23855. A–D, pleopods –3, 5 respectively; E,<br />
uropod; F, uropod exopod, ventral view; G, uropod exopod apex.<br />
Rocinela leptopus sp. nov. (Figs 0– 3)<br />
materiaL examined: Holotype: ♀ (34 mm, ovig.), <strong>of</strong>f<br />
Pegasus Bay, South Island, 43° 4’S, 75°39’E, 27 September<br />
976, <strong>of</strong>f steep wall, coral, 006–5 2 m, stn.<br />
S5559 (NMNZ Cr. 20 0).<br />
desCription: Body .8 times as long as greatest width,<br />
dorsal surfaces smooth or sparsely punctate, widest<br />
at pereonite 5, lateral margins ovate. Rostrum simple,<br />
74<br />
anteriorly subtruncate. Eyes not medially united, separated<br />
by about 40% width <strong>of</strong> head; each eye made up<br />
<strong>of</strong> ~ 2 transverse rows <strong>of</strong> ommatidia, each row with<br />
~7 ommatidia; eye colour black. Coxae 2–3 each with<br />
posteroventral angle rounded; 5–7 without oblique<br />
carina. Pleon with pleonite largely concealed by<br />
pereonite 7; pleonite 4 with posterolateral margins<br />
extending clearly beyond posterior margin <strong>of</strong> pleonite<br />
5; pleonite 5 with posterolateral angles acute. Pleotelson<br />
0.8 times as long as anterior width, anterior dorsal
Figure 110. Rocinela leptopus sp. nov. Holotype. A, dorsal view; B, lateral view; C, head, dorsal view ; D, frons;<br />
E, pleonites 2–5, lateral margin; F, antennule; G, antenna peduncle.<br />
surface without 2 sub-median depressions, dorsal<br />
surface smooth; lateral margins convex, posterior<br />
margin narrowly rounded, with 4 RS (distalmost RS<br />
ventrally directed).<br />
Antennule peduncle article 3 . times as long as<br />
combined lengths <strong>of</strong> articles and 2, 3.5 times as long<br />
as wide; flagellum with 6 articles, extending to anterior<br />
<strong>of</strong> pereonite . Antenna peduncle article 3 .4 times as<br />
long as article 2, . times as long as wide; article 4 .7<br />
times as long as article 3, .9 times as long as wide, inferior<br />
margin with 0 plumose setae, and simple seta<br />
(possibly with more setae as some are clearly missing);<br />
article 5 .2 times as long as article 4, 3.0 times as long<br />
as wide, inferior margin with 3 setae, anterodistal angle<br />
with cluster <strong>of</strong> 4 short simple setae; flagellum with 13<br />
articles, extending to middle <strong>of</strong> pereonite .<br />
Frontal lamina longer than greatest width, anteriorly<br />
acute.<br />
Mandible molar process distinct flat lobe; palp article<br />
2 with 2 marginal distolateral setae, and 2 long distolateral<br />
setae; palp article 3 with 24 setae. Maxillule with 6<br />
RS ( large, 5 slender, serrate). Maxilla mesial lobe with<br />
2 hooked RS; lateral lobe with 2 hooked RS. Maxilliped<br />
palp article distomesial angle with RS; article 2 with<br />
3 hooked RS; article 3 with 2 hooked RS.<br />
Pereopod 1 basis 3.2 times as long as greatest width;<br />
ischium 0.6 times as long as basis, inferior margin with 0<br />
RS, superior distal margin with 2 setae; merus inferior<br />
75
Figure 111. Rocinela leptopus sp. nov. Holotype. A, mandible; B, mandible incisor; C, mandible palp article 3; D, scales,<br />
mandible mesial margin; E, maxilliped palp articles – 3;E, maxillule apex; ; F, maxilla apex.<br />
margin with 2 RS, set as two groups, superior distal<br />
angle with 5 setae ( RS, 4 simple); carpus 0.6 times as<br />
long as merus, inferior margin with RS; propodus 4.4<br />
times as long as proximal width, propodal palm with<br />
small distal lobe, inferior margin with 2 RS (distal);<br />
dactylus 0.8 times as long as propodus (weakly curved).<br />
Pereopods 2 and 3 not similar to pereopod (far more<br />
robust, with +2 RS on inferior margin <strong>of</strong> merus; propodus<br />
2.0 times as long as proximal width, with 3 RS;<br />
dactylus about as long as propodus, weakly curved).<br />
Pereopod 6 similar to pereopod 7. Pereopod 7 basis 3.4<br />
times as long as greatest width, inferior margins with<br />
palmate setae; ischium 0.7 as long as basis, inferior<br />
margin with 7 RS (set as , 2, 2 and 2), superior distal<br />
angle with 5 RS, inferior distal angle with 6 RS; merus<br />
0.5 times as long as ischium, .6 times as long as wide,<br />
inferior margin with RS, superior distal angle with<br />
6 RS, inferior distal angle with 7 RS; carpus 0.6 times<br />
as long as ischium, 2.3 times as long as wide, inferior<br />
margin with 3 RS (set singly), superior distal angle<br />
76<br />
with 0 RS (and 2 setae), inferior distal angle with 6<br />
RS; propodus 0.5 as long as ischium, 3.6 times as long<br />
as wide, inferior margin with 5 RS (set as , , and<br />
2), superior distal angle with 6 slender setae, inferior<br />
distal angle with 2 RS.<br />
Pleopod 1 exopod 2.3 times as long as wide, lateral<br />
margin straight, mesial margin weakly convex, with<br />
PMS on distal two-thirds; peduncle mesial margin with<br />
6 coupling hooks.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about three-quarters as long as endopod.<br />
Exopod at angle <strong>of</strong> about 35° to endopod, rami<br />
extending to pleotelson apex, marginal setae in two<br />
tiers. Endopod lateral margin weakly convex, lateral<br />
margin with 7 RS, mesial margin straight, with 3 RS.<br />
Exopod not extending to end <strong>of</strong> endopod, 4.0 times as<br />
long as greatest width; lateral margin straight, with 8<br />
RS; mesial margin straight, distally convex, with 0 RS;<br />
distal margin with indistinct apex.
Figure 112. Rocinela leptopus sp. nov. Holotype. A–C, pereopods , 2 and 7 respectively; D, pereopod , distomesial margin<br />
<strong>of</strong> carpus; E, pereopod , dactylus and distal margin <strong>of</strong> propodus; F, pereopod 2, inferior margin <strong>of</strong> propodal palm;<br />
G, pereopod 7, distal margin <strong>of</strong> carpus, mesial RS; H, pereopod 7, dactylus and distal margin <strong>of</strong> propodus.<br />
size: <strong>The</strong> single specimen measures 34 mm.<br />
remarks: Rocinela leptopus sp. nov. is readily identified<br />
by the widely separated eyes, subtruncate rostrum,<br />
‘stepped’ anterior margin <strong>of</strong> the head, slender pereopod<br />
with an elongate propodus, pereopods –3 propodal<br />
palm without a blade, the propodal palm <strong>of</strong> pereopod<br />
lacking robust setae, that <strong>of</strong> pereopods 2 and 3 with<br />
widely spaced short robust setae and the dactylus <strong>of</strong><br />
pereopods –3 being only weakly curved.<br />
Nearly all other species <strong>of</strong> Rocinela have pereopods<br />
–3 with a propodal blade and the propodal blade or<br />
palm with prominent robust setae. <strong>The</strong> Caribbean<br />
Rocinela signata Schioedte and Meinert, 879a lacks<br />
robust setae on the pereopod palm, as does Rocinela<br />
media Nierstrasz, 93 , known only from Indonesia, but<br />
in both these species the propodus is short and robust.<br />
Only Rocinela runga sp. nov., from the relatively nearby<br />
Antipodes Islands, has an elongate and unarmed propodus,<br />
and that species may be distinguished by the<br />
77
Figure 113. Rocinela leptopus sp. nov. Holotype. A, pleopod ; B, pleopod 2; C, left uropod, dorsal view; D, uropodal exopod,<br />
ventral view.<br />
longer propodus on pereopods –3, the broad, anteriorly<br />
rounded rostrum and the pleonite lateral margins<br />
being less produced and acute.<br />
<strong>The</strong> holotype, while in good condition, is somewhat<br />
brittle, the pleopods breaking up on dissection. In situ<br />
examination indicates that they are similar to others<br />
<strong>of</strong> the genus.<br />
prey: Not known.<br />
distribution: Known only from the type locality, <strong>of</strong>f<br />
the northeastern coast <strong>of</strong> South Island.<br />
etymoLogy: Adapted from the Greek words leptos (thin,<br />
slender, delicate) and pous (foot) alluding to the slender<br />
first pereopods.<br />
Rocinela pakari sp. nov. (Figs 4– 8)<br />
materiaL examined: Holotype, ♀ (28 mm, non-ovig.),<br />
Chatham Rise, 25 October 979, 44°26.89’S, 74°54.79’E,<br />
676 m (NIWA 23888).<br />
78<br />
Paratypes. 4♀ (each 20 mm, non-ovig.), 8 mancas<br />
( 3, 4 mm, 6 unmeasured), same data as holotype<br />
(NIWA 23889).<br />
Additional material: ♀ (39 mm, non-ovig, poor condition,<br />
all but one anterior pereopod missing), (Fisheries<br />
Research Division) (NIWA 23890). ♀ (33 mm, ovig,<br />
poor condition, uropods and pleotelson largely absent),<br />
Chatham Rise, 44°34.00’S, 74°06.49’E, 29 October 979,<br />
863–9 0 m (NIWA 2389 ).<br />
desCription: Body 2.5 times as long as greatest width,<br />
widest at pereonite 4, lateral margins subparallel.<br />
Rostral point anteriorly subtruncate. Eyes not medially<br />
united, separated by about 2.5% width <strong>of</strong> head; each<br />
eye made up <strong>of</strong> ~ 6 transverse rows <strong>of</strong> ommatidia, each<br />
row with ~ ommatidia; eye colour black. Pereonite 1<br />
and coxae 2–3 each with posteroventral angle rounded;<br />
coxae 5–7 without oblique carina. Pleon with pleonite<br />
largely concealed by pereonite 7, or visible in dorsal<br />
view; pleonite 4 with posterolateral margins extending<br />
clearly beyond posterior margin <strong>of</strong> pleonite 5; pleonite
Figure 114. Rocinela pakari sp. nov. A–E, holotype, F, G, I, J and K, 20 mm paratype, others as indicated. A, dorsal view;<br />
B, lateral view; C, head; dorsal view, male; D, frons; E, pleonites 4 and 5, lateral margins; F, dorsal view; head, paratype;<br />
G, coxa 7; H, head, 33 mm ovigerous female, S 64; I, head; J, antennule; K, antenna.<br />
79
Figure 115. Rocinela pakari sp. nov. Paratype 20 mm. A, mandible; B, mandible molar and incisor; C, mandible palp article 3;<br />
D, maxillule; E, maxillule apex; F, maxilla apex; G, maxilla; H, maxilliped; I, maxilliped palp articles 2 and 3.<br />
5 with posterolateral angles acute. Pleotelson 0.8 times<br />
as long as anterior width, anterior dorsal surface with 2<br />
sub-medial depressions, dorsal surface smooth; lateral<br />
margins weakly convex, posterior margin narrowly<br />
rounded, with 5 robust setae.<br />
Antennule peduncle article 3 0.9 times as long as<br />
combined lengths <strong>of</strong> articles and 2, 3.3 times as long<br />
as wide; flagellum with 5 articles (article 1 elongate),<br />
80<br />
extending to anterior <strong>of</strong> pereonite . Antenna peduncle<br />
article 3 .7 times as long as article 2; article 4 .3 times<br />
as long as article 3, .4 times as long as wide, inferior<br />
margin with 0 plumose setae, and 3 simple setae; article<br />
5 .5 times as long as article 4, 2.5 times as long as wide,<br />
inferior margin with 3 setae (palmate), anterodistal angle<br />
with cluster <strong>of</strong> 4 short simple setae; flagellum with<br />
9 articles, extending to middle <strong>of</strong> pereonite 2.
Figure 116. Rocinela pakari sp. nov. Holotype. A–C, pereopods , 2 and 7 respectively; D, robust setae on pereopod blade;<br />
E, pereopod ischium, mesial surface; F, pereopod 7 distal margin <strong>of</strong> carpus, mesial surface; G, pereopod , 33 mm ovigerous<br />
female, NIWA 2389 .<br />
Frontal lamina longer than greatest width, anteriorly<br />
acute.<br />
Mandible molar process distinct flat lobe; palp article<br />
2 with 0 short, marginal distolateral setae, and<br />
3 long distolateral setae; palp article 3 with 25 setae.<br />
Maxillule with 6 RS ( large, 5 slender, serrate). Maxilla<br />
mesial lobe with 2 hooked RS (small); lateral lobe with<br />
2 hooked RS. Maxilliped palp article distomesial angle<br />
8<br />
with 2 robust setae (straight); article 2 with 3 hooked<br />
RS; article 3 with 2 hooked RS.<br />
Pereopod 1 basis 2.4 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with seta (and 2 plumose<br />
setae); merus inferior margin with 3 RS, set as two<br />
groups, superior distal angle with 6 setae (plumose);<br />
carpus 0.7 as long as merus, inferior margin with RS;
Figure 117. Rocinela pakari sp. nov. Paratype 20 mm. A–D, pleopods –3, 5 respectively; E, pleopod 3 peduncle lateral margin;<br />
F, uropod; G, uropod exopod, ventral view.<br />
propodus .7 times as long as proximal width, propodal<br />
palm with blade, propodal blade . times as wide as<br />
palm, inferior margin with 3 RS; dactylus .8 times as<br />
long as propodus. Pereopods 2 and 3 similar to pereopod<br />
(RS on merus longer). Pereopod 6 similar to pereopod<br />
82<br />
7. Pereopod 7 basis 3.7 times as long as greatest width,<br />
inferior margins with 6 palmate setae; ischium 0.7 as<br />
long as basis, inferior margin with 7 RS (set as , 2, 2<br />
and 2), superior distal angle with 4 RS, inferior distal<br />
angle with 4 RS; merus 0.6 times as long as ischium, 2.2
Figure 118. Rocinela pakari sp. nov. A, holotype, uropod in situ; B–D, ovigerous female, 33 mm: B, maxilliped; C, maxilliped<br />
palp; D, plumose setae, distal margin <strong>of</strong> lamina vibrans.<br />
times as long as wide, inferior margin with 4 RS (set as<br />
and 3), superior distal angle with 5 RS, inferior distal<br />
angle with 4 RS; carpus 0.6 times as long as ischium,<br />
2.9 times as long as wide, inferior margin with 5 RS (set<br />
as , 2 and 2), superior distal angle with 9 RS, inferior<br />
distal angle with 5 RS; propodus 0.6 as long as ischium,<br />
4. times as long as wide, inferior margin with 3 RS<br />
(set singly), superior distal angle with 3 slender setae<br />
( palmate), inferior distal angle with 2 RS.<br />
Pleopod 1 exopod 2.3 times as long as wide, lateral<br />
margin weakly convex, mesial margin weakly convex,<br />
with PMS from distal one-third; endopod 2.6 times<br />
as long as wide, lateral margin weakly concave, with<br />
PMS on distal margin only, mesial margin with PMS<br />
from distal one-third; peduncle mesial margin with 6<br />
coupling hooks. Pleopods 2–5 peduncle distolateral<br />
margin with acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about one-third as long as endopod. Exopod<br />
at angle <strong>of</strong> about 35° to endopod, rami extending<br />
to pleotelson apex, marginal setae in two tiers. Endopod<br />
lateral margin convex, lateral margin with 8 RS, mesial<br />
margin strongly convex, with 4 RS. Exopod extending<br />
to end <strong>of</strong> endopod, 2.3 times as long as greatest width;<br />
lateral margin convex, with 4 RS; mesial margin sinuate,<br />
proximally concave, distally convex, with 4 RS;<br />
distal margin rounded.<br />
83<br />
maLe: Not known<br />
size: Females 20 and 39 mm; mancas 3– 4 mm.<br />
Variation: Based on measured types only. Robust setae:<br />
Pleotelson (n = 7) RS 3– 7, with 4 (twice) and 5<br />
(three times) most frequent. Uropod exopod (n = 4)<br />
mesial margin with 0–4 with (2 %) and 2 (43%) most<br />
frequent, 0 and 4 each occurring twice, 3 once; lateral<br />
margin 2– 4 with 3 (50%) and 4 (36%) most frequent.<br />
Uropod endopod (n = 4) mesial margin varied<br />
from 4 to 6 with 4 (50%) and 5 (36%) most frequent, 6<br />
occurring twice (one specimen); lateral margin with<br />
6–8 with 6 (2 %) and 7 (64%) most frequent, 8 occurring<br />
twice.<br />
<strong>The</strong> setation <strong>of</strong> the pereopod palm is highly consistent<br />
with robust setae on pereopods –3, ranging from
to 4, with the following numbers (n = 4): pereopod<br />
with 2 (43%) or 3 (50%), once (smallest manca);<br />
pereopod 2 with 2 (50%), (three times), or 3 and<br />
4, each twice; pereopod 3 2 (64%) or 3 (three times)<br />
and 4 once; pereopod never had more than 3 robust<br />
setae on the propodal palm and no specimen had<br />
less than robust setae on any pereopod palm. <strong>The</strong><br />
robust setae on the inferior margin <strong>of</strong> the merus were<br />
consistently +2.<br />
<strong>The</strong>re was no difference between adults and<br />
mancas in the number <strong>of</strong> robust setae on the anterior<br />
pereopods. <strong>The</strong> rostrum is longer and straight in adult<br />
specimens whereas in mancas and small specimens it<br />
is bent slightly to the ventral.<br />
<strong>The</strong> single damaged ovigerous female had oo-<br />
stegites on sternites –5. <strong>The</strong> robust setae on the propodal<br />
palm <strong>of</strong> pereopods –3 are notably more slender<br />
and longer than those <strong>of</strong> immature specimens or<br />
non-ovigerous females; the number <strong>of</strong> robust setae on<br />
the palm <strong>of</strong> pereopods –3 is 3, as the most frequent<br />
number for the other specimens.<br />
remarks: Rocinela pakari sp. nov. can be identified by<br />
the wide propodal blade on pereopods –3 which is<br />
provided with – 4 robust setae in conjunction with<br />
well-separated eyes and a gently narrowed pleotelson<br />
posterior margin; in mature specimens the rostrum is<br />
relatively short, anteriorly rounded, and the lateral<br />
margins are very weakly stepped.<br />
Only four species <strong>of</strong> Rocinela have pereopods –3<br />
with propodal blade as wide as the palm and provided<br />
with more than eight robust setae. Of those that do,<br />
three have eyes that meet in the middle (these being<br />
R. affinis, R. kapala and R. oculata) and all <strong>of</strong> these have<br />
fewer than 0 robust setae on the pereopod palm; the<br />
fourth species, Rocinela niponia Richardson, 909, a<br />
species in need <strong>of</strong> redescription, has separate eyes,<br />
fewer robust setae on the pereopod blade ( 0, 8 and<br />
8 on pereopods to 3 respectively) and more robust<br />
setae (5) on the inferior margin <strong>of</strong> the merus.<br />
prey: Not known.<br />
distribution: Known only from the Chatham Rise,<br />
eastwards from the mid-coast <strong>of</strong> South Island.<br />
etymoLogy: Pakari is a Mäori word that means strong<br />
(noun in apposition).<br />
Rocinela resima sp. nov. (Figs 9– 22)<br />
materiaL examined: Holotype, ♂ (29 mm), Christabel<br />
sea mount, northeastern Macquarie Ridge, 5 °04.34’S,<br />
64°36.37’E, 4 April 2003, 065– 030 m, rubble, (NIWA<br />
23883).<br />
Paratypes: ♂ (20 mm, dissected), Chatham Rise,<br />
42°43.95’S, 79°53.9 ’W, 8 April 200 , 076–990 m<br />
(NIWA 23884). ♀ (non-ovig. 34 mm), 42°46.99’S,<br />
84<br />
79°59.64’E, 2 April 200 , 000–870 m (NIWA 23885).<br />
♂? (24 mm), Chatham Rise, 42° 46.07’E, 79°55.3 ’W,<br />
20-April 200 , 955–890 m (NIWA 23886).<br />
Non-type material. ♀ (30 mm, non-ovig.), same data<br />
as holotype (NIWA 23887).<br />
Additional material. Chatham Rise. ♀ (24 mm, nonovig.),<br />
42.7597–7557°S, 79.0 05–0 2°W, 28 May 2006,<br />
765–845 m (NIWA 25655). ♀ (2 mm, ovig.), 42.7885–<br />
7992°S, 79.9985–9982°E, 30 May 2006, 020– 054<br />
m (NIWA 25665). 2♀ (2 mm, one damaged, ovig.),<br />
42.7 70–7 65°S, 79.0420–0440°E, 3 May 2006, 957–985<br />
m (NIWA 25666).<br />
desCription: Body 2. times as long as greatest width,<br />
dorsal surfaces smooth, sparsely punctate, widest at<br />
pereonite 5, lateral margins weakly ovate. Rostrum<br />
turned upwards, anteriorly truncate (margins thickened).<br />
Eyes not mesially united, separated by about<br />
28% width <strong>of</strong> head; each eye made up <strong>of</strong> ~ 3 transverse<br />
rows <strong>of</strong> ommatidia, each row with ~9 ommatidia;<br />
eye colour dark brown (bronze). Coxae 2–3 each with<br />
posteroventral angle acute, posteriorly produced; 5–7<br />
without oblique carina. Pleon with pleonite largely<br />
concealed by pereonite 7; pleonite 4 with posterolateral<br />
margins extending to, but not beyond, posterior margin<br />
<strong>of</strong> pleonite 5; pleonite 5 with posterolateral angles<br />
rounded. Pleotelson .2 times as long as anterior width,<br />
anterior dorsal surface without 2 sub-median depressions,<br />
dorsal surface with short setae; lateral margins<br />
convex, posterior margin with distinct median point,<br />
with 4 RS.<br />
Antennule peduncle article 3 0.9 times as long as<br />
combined lengths <strong>of</strong> articles and 2, 3. times as long as<br />
wide; flagellum with 6 articles, extending to anterior <strong>of</strong><br />
pereonite . Antenna peduncle article 3 2.3 times as long<br />
as article 2, .2 times as long as wide; article 4 .2 times<br />
as long as article 3, .4 times as long as wide, inferior<br />
margin with 0 plumose setae, and 2 simple setae (stiff);<br />
article 5 .6 times as long as article 4, 2.3 times as long<br />
as wide, inferior margin with 2 setae, anterodistal angle<br />
with cluster <strong>of</strong> 6 short simple setae; flagellum with 17<br />
articles, extending to posterior <strong>of</strong> pereonite 2.<br />
Frontal lamina longer than greatest width, anteriorly<br />
acute.<br />
Mandible molar process distinct flat lobe; palp article<br />
2 with marginal distolateral setae, and 3 long distolateral<br />
setae; palp article 3 with 20 setae (all distally<br />
bifurcate except distalmost 2 setae). Maxillule with 6 RS<br />
( large, 5 slender, serrate). Maxilla mesial lobe with 2<br />
hooked RS; lateral lobe with 2 hooked RS. Maxilliped<br />
palp article distomesial angle with 0 RS (with long<br />
seta and less long seta mid-margin); article 2 with<br />
3 hooked RS (2 distal, proximal and straight, stiff<br />
seta); article 3 with 2 hooked RS.<br />
Pereopod 1 basis 2.9 times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with
Figure 119. Rocinela resima sp. nov. A–F, holotype; G, female NIWA 23885; H–J, male paratype NIWA 23884. A, dorsal view;<br />
B, lateral view; C, head; dorsal view; D, frons; E, pleonites 2–5 and uropod peduncle, lateral margins; F, sternite 7 showing<br />
penial papillae; G, head, pereonite ; H, antennule; I, setae, distal margin antennule peduncle article ; J, antenna.<br />
85
Figure 120. Rocinela resima sp. nov. Paratype NIWA 23884. A, mandible; B, mandible palp article 3; C, maxillule apex;<br />
D, maxillule; E, maxilla; F, maxilla apex; G, maxilliped; H, maxilliped palp articles 2 and 3; I, maxilliped palp articles 2 and<br />
3, NIWA 23885.<br />
0 RS, superior distal margin with 7 setae (including<br />
acute RS); merus inferior margin with 3 RS, set as two<br />
rows (<strong>of</strong> and 2), superior distal angle with 7 setae<br />
(long); carpus 0.6 times as long as merus, inferior margin<br />
with RS; propodus .6 times as long as proximal<br />
width, propodal palm with blade, propodal blade 0.5<br />
times as wide as palm, with single seta, inferior margin<br />
with 3 RS; dactylus . times as long as propodus. Pereopods<br />
2 and 3 similar to pereopod (but larger, meral<br />
RS larger). Pereopod 6 similar to pereopod 7. Pereopod<br />
7 basis 3.5 times as long as greatest width, inferior<br />
margins with 8 palmate setae; ischium 0.7 as long as<br />
basis, inferior margin with 5 RS (set as , , 2 and ),<br />
superior distal angle with 6 RS, inferior distal angle<br />
with 6 RS; merus 0.5 times as long as ischium, .9 times<br />
86<br />
as long as wide, inferior margin with RS, superior<br />
distal angle with 7 RS, inferior distal angle with 6 RS;<br />
carpus 0.6 times as long as ischium, 2.4 times as long<br />
as wide, inferior margin with 2 RS (set as and ),<br />
superior distal angle with 0 RS, inferior distal angle<br />
with 4 RS; propodus 0.5 as long as ischium, 3.5 times<br />
as long as wide, inferior margin with 3 RS (set as and<br />
2), superior distal angle with 6 slender setae, inferior<br />
distal angle with 3 RS.<br />
Penes low tubercles; penial openings separated by<br />
6% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod 2.2 times as long as wide, lateral<br />
margin straight, mesial margin weakly convex, with<br />
PMS on distal two-thirds; endopod 2.6 times as long<br />
as wide, lateral margin straight, with PMS on on dis-
Figure 121. Rocinela resima sp. nov. A, pereopod , holotype; B, pereopod 2, holotype; C, pereopod 7, holotype ; D, pereopod<br />
, NIWA 23885; E, pereopod 2, NIWA 23885; F, pereopod , holotype; G, robust setae, carpus mesial margin, pereopod 7.<br />
tal margin only, mesial margin with PMS on distal<br />
two-thirds; peduncle mesial margin with 7 coupling<br />
hooks. Pleopod 2 appendix masculina with straight<br />
margins, 0.8 times as long as endopod, distally bluntly<br />
rounded. Pleopods 2–5 peduncle distolateral margin<br />
with acute RS.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about one-half as long as endopod.<br />
Exopod at angle <strong>of</strong> about 35° to endopod, rami extending<br />
to pleotelson apex. Endopod lateral margin convex,<br />
with 6 RS; mesial margin weakly convex, with 2 RS.<br />
Exopod not extending to end <strong>of</strong> endopod, 3.0 as long<br />
as greatest width; lateral margin weakly convex, with<br />
0 RS; mesial margin sinuate, proximally concave,<br />
distally convex, with 0 RS; distal margin with indistinct<br />
apex.<br />
87
G<br />
Figure 122. Rocinela resima sp. nov. Paratype NIWA 23884. A–C, pleopods –3, respectively; D, holotype, uropod; E, holotype,<br />
uropodal endopod, ventral view; F, uropod, female paratype NIWA 23887; G, uropod endopod apex, female paratype NIWA<br />
23885.<br />
femaLe: Non-ovigerous specimens similar to the male<br />
but: body more elongate (2.5 times as long as wide),<br />
rostrum shorter, anteriorly narrowly rounded; all<br />
females and specimens <strong>of</strong> indeterminate sex with<br />
uropod exopod extending to just short <strong>of</strong> endopod<br />
apex; marginal setae less dense. Pereopods –3 palm<br />
with 2 large and small RS or 3 RS <strong>of</strong> approximately<br />
equal size (as in males); dactylus longer ( .4– .7 times<br />
88<br />
as long as propodus) than in males ( . times as long<br />
as propodus); eyes larger, separated by 6% width <strong>of</strong><br />
head; maxillule RS more slender than in mature male;<br />
maxilliped lacking mesial plumose setae, with more<br />
strongly hooked RS on palp articles and 2.<br />
size: Males 20–29 mm; female 34 mm.
Variation: Based on the four type specimens. Robust<br />
setae: Pleotelson RS 4– 7. Uropod exopod mesial<br />
margin with 0 (all); lateral margin 0– 2. Uropod endopod<br />
mesial margin varied from 2–4; lateral margin<br />
(n = 0) with 5–8.<br />
<strong>The</strong> setation <strong>of</strong> the pereopod palm is consistent with<br />
3 robust setae on pereopods –3; variation occurs in<br />
the distal robust seta on the palm which in the mature<br />
males is equal in size to the other robust setae, but in<br />
the immature male and some <strong>of</strong> the females it is less<br />
than half the size <strong>of</strong> the other robust setae (this can vary<br />
within the individual). <strong>The</strong> robust setae on the inferior<br />
margin <strong>of</strong> the merus were consistently +2.<br />
remarks: Rocinela resima sp. nov. can be identified by<br />
the ovate body shape, strongly produced and upturned<br />
rostrum, relatively widely separated eyes, pereopods<br />
–3 with three robust setae on a small, rounded propodal<br />
blade, and relatively narrow uropodal rami with<br />
posteriorly rounded uropodal endopod. Males have<br />
upturned and truncate rostrum, and relatively short<br />
and robust dactyli on pereopods –3.<br />
In <strong>New</strong> <strong>Zealand</strong> waters Rocinela bonita sp. nov. is<br />
the most similar species. R. resima can be distinguished<br />
from that species by a number <strong>of</strong> characters including<br />
pereopods –3 having a less produced and more<br />
rounded propodal blade with three robust setae (v.<br />
sub-rectangular with four robust setae in R. bonita),<br />
shorter and more robust dactylus in mature males<br />
( .0 times as long as propodus v. 2.0 times as long as<br />
propodus in R. bonita), shorter and more stout robust<br />
setae on the merus, shorter robust setae on the inferior<br />
margins <strong>of</strong> pereopod 7, and the uropodal endopod<br />
with an evenly convex lateral margin and smoothly<br />
rounded distal margin (v. sinuate lateral margin, apex<br />
with distinct apical point), and a narrower uropodal<br />
exopod (3.0 times as long as wide v. 2.4 times as long<br />
as wide in R. bonita).<br />
<strong>The</strong>re are two northern Pacific species, both showing<br />
some similarity to Rocinela resima sp. nov. Rocinela<br />
hawaiiensis Richardson, 903 is known from only two<br />
specimens (from Hawai’i and Pacific Mexico) and the<br />
adult male has not been described (Brusca & France<br />
992). <strong>The</strong> female <strong>of</strong> R. resima differs in having the distalmost<br />
robust seta on the propodal palm <strong>of</strong> pereopods<br />
–3 small, whereas in R. hawaiiensis all three robust<br />
setae are <strong>of</strong> equal length, and the pleotelson posterior<br />
margin <strong>of</strong> R. resima has a distinct median point while in<br />
R. hawaiiensis it is evenly rounded (Brusca & France<br />
992). <strong>The</strong> other similar species is Rocinela cornuta<br />
Richardson, 898, known from Alaska and Arctic waters<br />
(Kussakin 1979; Rafi 1985), a poorly known species<br />
for which few descriptive data are available. While<br />
the anterior margin <strong>of</strong> the head is similarly produced<br />
in mature males <strong>of</strong> both species, R. resima lacks the<br />
anterolateral projections on pereonite and has only<br />
three robust setae on the propodal palm <strong>of</strong> pereopods<br />
–3 rather than the four in R. cornuta; in addition<br />
Richardson (1898) figured the uropods <strong>of</strong> R. cornuta<br />
as extending well beyond the posterior margin <strong>of</strong> the<br />
pleotelson, whereas in R. resima the uropods reach only<br />
to that margin. Kussakin (1979) gave additional figures<br />
for the species, which correspond to neither those <strong>of</strong><br />
Richardson ( 898) nor any other species.<br />
89<br />
prey: Not known.<br />
distribution: All records from <strong>of</strong>f southeastern South<br />
Island in the region <strong>of</strong> the Chatham Rise and south<br />
to Christabel Sea Mount on the northern Macquarie<br />
Ridge; at depths <strong>of</strong> 870– 076 m.<br />
etymoLogy: Adapted from the Latin resimus (turnedup<br />
nose; simus = pug-nosed) and alluding to the<br />
prominent, somewhat upturned rostrum in the adult<br />
males.<br />
Rocinela runga sp. nov. (Figs 23– 25)<br />
materiaL examined: Holotype, ♀ (35 mm, non-ovig.),<br />
49°38. 0–04’S, 78°47.5 –26’E, <strong>of</strong>f Antipodes Islands,<br />
23 April 2003, 03– 08 m (NIWA 23845).<br />
desCription: Body 2.0 times as long as greatest width,<br />
dorsal surfaces smooth and sparsely punctate, widest<br />
at pereonite 5, lateral margins weakly ovate. Rostrum<br />
simple, anteriorly rounded. Eyes not medially united,<br />
separated by about 40% width <strong>of</strong> head; each eye made<br />
up <strong>of</strong> ~ 2 transverse rows <strong>of</strong> ommatidia, each row with<br />
~9 ommatidia; eye colour dark brown. Pereonite 1 and<br />
coxae 2–3 each with posteroventral angle rounded;<br />
coxae 5–7 with incomplete oblique carina (weak). Pleon<br />
with pleonite largely concealed by pereonite 7; pleonite<br />
4 with posterolateral margins extending to, but<br />
not beyond, posterior margin <strong>of</strong> pleonite 5; pleonite<br />
5 with posterolateral angles acute. Pleotelson . times<br />
as long as anterior width, anterior dorsal surface with<br />
2 sub-median depressions, dorsal surface with short<br />
setae; lateral margins weakly convex, posterior margin<br />
narrowly rounded, with 6– 8 RS (many missing).<br />
Antennule peduncle article 3 0.8 times as long as<br />
combined lengths <strong>of</strong> articles and 2 (in situ), 3.5 times<br />
as long as wide; flagellum with 6 articles, extending<br />
to anterior <strong>of</strong> pereonite . Antenna peduncle article 3<br />
2.8 times as long as article 2, .3 times as long as wide;<br />
article 4 .5 times as long as article 3, .9 times as long<br />
as wide, inferior margin with 0 plumose setae, and<br />
simple setae; article 5 .3 times as long as article 4,<br />
3.2 times as long as wide, inferior margin with 4 setae<br />
(minute, widely spaced), anterodistal angle with cluster<br />
<strong>of</strong> 2 short simple setae (and 2 plumose setae); extending<br />
to posterior <strong>of</strong> pereonite 2.
Figure 123. Rocinela runga sp. nov. Holotype. A, dorsal view; B, lateral view; C, head, dorsal view; D, frons; E, pleonites,<br />
oblique lateral view; F, antennule, in situ (dorsal view); G, antenna, in situ (ventral view).<br />
Frontal lamina as wide as long, anteriorly acute.<br />
Mandible molar process distinct flat lobe; palp article<br />
2 with 2 marginal distolateral setae (all with distinctly<br />
bifurcate tips), and 2 long distolateral setae; palp article<br />
3 with 22 setae (all distally bifurcate except distalmost<br />
seta). Maxillule with 6 RS ( large, 5 slender, 2 <strong>of</strong> which<br />
serrate). Maxilla mesial lobe with 2 hooked RS; lateral<br />
lobe with 2 hooked RS. Maxilliped palp article distomesial<br />
angle with 2 RS ( short, straight, hooked);<br />
article 2 with 2 hooked RS; article 3 with 2 hooked RS<br />
(article 3 proximally fused to article 2).<br />
Pereopod 1 basis 3.2 times as long as greatest width;<br />
ischium 0.6 times as long as basis, inferior margin with 0<br />
RS, superior distal margin with 7 setae (and acute RS);<br />
merus inferior margin with 2 RS (minute), set as two<br />
90<br />
groups, superior distal angle with 6 setae (all simple);<br />
carpus 0.7 times as long as merus, inferior margin with<br />
0 RS; propodus 4.4 times as long as proximal width, propodal<br />
palm simple, without blade or process, inferior<br />
margin with RS (distal; minute); dactylus .0 times as<br />
long as propodus (curved distally). Pereopods 2 and 3<br />
not similar to pereopod (more robust, with +2 RS on<br />
inferior margin <strong>of</strong> merus; propodus 2.6 times as long<br />
as proximal width, with 2 small RS; dactylus slightly<br />
longer ( .03) than propodus, weakly curved). Pereopod<br />
6 similar to pereopod 7 (but longer). Pereopod 7 basis 3.7<br />
times as long as greatest width, inferior margins with<br />
7 palmate setae; ischium 0.7 as long as basis, inferior<br />
margin with 7 RS (set as , 2, 2 and 2), superior distal<br />
angle with 4 RS, inferior distal angle with 6 RS; merus
Figure 124. Rocinela runga sp. nov. Holotype. A, mandible; B, mandible molar and incisor; C, mandible palp article 3;<br />
D, robust seta, mandible palp article 2; E, maxillule; F, maxillule apex; G, maxilla; H, maxilla apex; I, maxilliped; J, maxilliped,<br />
palp articles –3.<br />
0.6 times as long as ischium, .9 times as long as wide,<br />
inferior margin with 2 RS (paired), superior distal angle<br />
with 9 RS, inferior distal angle with 5 RS; carpus<br />
0.6 times as long as ischium, 2.4 times as long as wide,<br />
inferior margin with RS (and minute proximal seta),<br />
superior distal angle with 9 RS (4 short), inferior distal<br />
angle with 8 RS; propodus 0.6 as long as ischium, 3.6<br />
times as long as wide, inferior margin with 5 RS (set as<br />
, 2 and 2), superior distal angle with 5 slender setae,<br />
inferior distal angle with 3 RS.<br />
Pleopods swollen and distended, not described; examined<br />
in situ, appearing similar to those <strong>of</strong> Rocinela<br />
leptopus sp. nov.<br />
9<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about three-quarters as long as endopod.<br />
Exopod at angle <strong>of</strong> about 35° to endopod, rami not extending<br />
to pleotelson apex, marginal setae in two tiers.<br />
Endopod lateral margin weakly convex, lateral margin<br />
with 4 RS, mesial margin distally rounded, with 4 RS.<br />
Exopod not extending to end <strong>of</strong> endopod, 3.4 times as<br />
long as greatest width; lateral margin weakly convex,<br />
with 8 RS; mesial margin straight, distally convex, with<br />
0 RS; distal margin with indistinct apex.
Figure 125. Rocinela runga sp. nov. Holotype. A–C, pereopods , 2 and 7 respectively; D, pereopod , merus, distal inferior<br />
margin; E, pereopod , propodus, distal inferior margin, F, pereopod 2 ischium, mesial angle; G, pereopod 7, distal margin <strong>of</strong><br />
carpus, mesial RS; H, uropod; I, uropodal exopod, ventral view.<br />
Variation: <strong>The</strong> uropodal endopod mesial margin had<br />
4 and 6 robust setae, lateral margin 4 and 5 robust<br />
setae.<br />
remarks: Rocinela runga sp. nov. can be identified by<br />
the ovate body shape, relatively small and widely<br />
separated eyes, smoothly narrowed rostrum, very<br />
long (longer than pereopod 7) and slender pereopod<br />
92<br />
, pereopods –3 without a propodal blade and by the<br />
relatively narrow uropodal rami.<br />
Rocinela leptopus sp. nov. is the only other similar<br />
species, with a similar pereopodal, pleotelson and<br />
uropod morphology. <strong>The</strong> two species are readily separated<br />
by R. runga having a more elongate pereopod<br />
, pereopod dactylus distally curved (proximally<br />
curved in R. leptopus), more slender pereopods 2 and
3, and uropodal rami that are relatively wider (exopod<br />
3.4 times as long as wide) than in R. leptopus (exopod<br />
4.0 times as long as wide) and which fail to exceed the<br />
posterior margin <strong>of</strong> the pleotelson (just exceeding the<br />
posterior margin <strong>of</strong> the pleotelson in R. leptopus.<br />
prey: Not known.<br />
distribution: Known only from <strong>of</strong>f the Antipodes<br />
Islands, eastern Campbell Plateau, southeast <strong>of</strong> <strong>New</strong><br />
<strong>Zealand</strong>.<br />
etymoLogy: Runga is a Mäori word meaning south<br />
(location) alluding to the southern location (noun in<br />
apposition).<br />
Rocinela satagia sp. nov. (Figs 26– 29)<br />
materiaL examined: Holotype, ♂ (25 mm) Chatham Rise,<br />
43°49.605’S, 78°29.284’E, 9 October 200 , 454 m, coll.<br />
RV Tangaroa (NIWA 23856).<br />
Paratypes: 2♀ (non-ovig. 2 [dissected], 8.5 mm,),<br />
same data as holotype (NIWA 23857). ♀ (2 mm,<br />
non-ovig.), Chatham Rise, 43.7033°S, 79.9 7°E, no<br />
date, stn. Q4a, 398 m, medium Agassiz trawl (NIWA<br />
23858). ♂ (20 mm), eastern Chatham Rise, 44°09.60’S,<br />
79° 4.20’W, 7 March 978, 320 m, stn Q20 (NIWA<br />
23859). ♂ (2 mm), <strong>of</strong>f East Otago coast, South Island,<br />
45°45.4’S, 7 °05.0’E, 6 August 955, 584 m, canyon<br />
B, M.V. Alert (NMNZ Cr. 2007). ♂ ( 8.5 mm), ♀ (ovig<br />
24 mm), manca (7.5 mm), eastern Chatham Rise,<br />
43°44.92’–44°0 .60’S, 79°00.34–0 .60’W, 8 September<br />
989, 397–399 m, stn V365, (NIWA 23860). Manca ( 2.0<br />
mm), eastern Chatham Rise, 44°05.50’S, 79°06.00’W,<br />
February 968, 322 m, stn G0327 (NIWA 2386 ).<br />
Other material: ♀ ( 9 mm, ovig., poor condition), c. 43<br />
km southeast <strong>of</strong> Cape Campbell, 4 °55.9’S, 4 °43.2’E,<br />
4 January 979, 454–424 m, stn BS668 (= NZOI stn<br />
R26), RV Tangaroa (NIWA 23880). ♀ ( 8 mm, ovig.),<br />
43.5328–5348°S, 79.6280–6257°E, 6 June 2006, 375–38<br />
m (NIWA 25669). ♀ ( 3.0 mm, non-ovig), Pegasus<br />
Canyon, Pegasus Bay, 43° 4’S, 73°39’E, 29 September<br />
976, BS559, 006–5 2 m, coral, coll. RV Acheron<br />
(NMNZ Cr. 2008). Manca? (8.5 mm, poor condition),<br />
Pegasus Canyon, Pegasus Bay, 43°30.0’S, 73°3 .3’E, 27<br />
September 976, BS558, 446 m, mud, coll. RV Acheron<br />
(NMNZ Cr. 2009).<br />
desCription: Body 2.0 times as long as greatest width,<br />
dorsal surfaces smooth, widest at pereonite 5, lateral<br />
margins weakly ovate. Rostrum basally expanded, tricornered.<br />
Eyes not medially united, separated by about<br />
27% width <strong>of</strong> head; each eye made up <strong>of</strong> ~ 0 transverse<br />
rows <strong>of</strong> ommatidia, each row with ~8 ommatidia; eye<br />
colour dark brown. Coxae 2–3 each with posteroventral<br />
angle right-angled (coxae 3 rounded); 5–7 without<br />
oblique carina. Pleon with pleonite largely concealed<br />
by pereonite 7; pleonite 4 with posterolateral margins<br />
extending clearly beyond posterior margin <strong>of</strong> pleonite<br />
5; pleonite 5 with posterolateral angles rounded. Pleotelson<br />
0.8 times as long as anterior width, anterior dorsal<br />
surface without 2 sub-median depressions, dorsal surface<br />
smooth; lateral margins convex, posterior margin<br />
narrowly rounded, with 8– 0 RS.<br />
Antennule peduncle article 3 0.8 times as long as<br />
combined lengths <strong>of</strong> articles and 2, 3.4 times as long<br />
as wide; flagellum with 6 articles, extending to anterior<br />
<strong>of</strong> pereonite . Antenna peduncle article 3 .3 times as<br />
long as article 2, .2 times as long as wide; article 4 .5<br />
times as long as article 3, .8 times as long as wide,<br />
inferior margin with 0 plumose setae, and simple<br />
setae; article 5 .5 times as long as article 4, 2.9 times<br />
as long as wide, inferior margin with 2 setae (palmate),<br />
anterodistal angle with cluster <strong>of</strong> 3 short simple setae;<br />
flagellum with 14 articles, extending to middle <strong>of</strong><br />
pereonite 2.<br />
Frontal lamina longer than greatest width, anteriorly<br />
rounded.<br />
Mandible molar process distinct flat lobe; palp article<br />
2 with 7 marginal distolateral setae (finely biserrate),<br />
and 3 long distolateral setae; palp article 3 with 6 setae<br />
(terminal 2 longest). Maxillule with 5 RS ( large, 4<br />
slender). Maxilla mesial lobe with 2 hooked RS; lateral<br />
lobe with hooked RS. Maxilliped palp article distomesial<br />
angle with 3 RS (slender, straight); article 2 with<br />
3 hooked RS; article 3 with hooked RS.<br />
Pereopod 1 basis 2.8 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 2 setae (and RS);<br />
merus inferior margin with 3 RS (set as + 2), set as two<br />
groups, superior distal angle with 4 setae (including<br />
RS); carpus 0.6 times as long as merus, inferior margin<br />
with RS; propodus .7 times as long as proximal width,<br />
propodal palm with blade, propodal blade 0.7 times<br />
as wide as palm, with numerous setae, inferior margin<br />
with 5 RS; dactylus .3 times as long as propodus. Pereopods<br />
2 and 3 similar to pereopod . Pereopod 6 similar<br />
to pereopod 7. Pereopod 7 basis 3.9 times as long as<br />
greatest width, inferior margins with 2 palmate setae<br />
(most rubbed away); ischium 0.8 as long as basis,<br />
inferior margin with 6 RS (set as , , , 2, ), superior<br />
distal angle with 6 RS, inferior distal angle with 5<br />
RS; merus 0.5 times as long as ischium, 2.0 as long as<br />
wide, inferior margin with 2 RS (set singly), superior<br />
distal angle with 7 RS, inferior distal angle with 5 RS;<br />
carpus 0.5 times as long as ischium, 2.4 times as long<br />
as wide, inferior margin with 2 RS (set singly), superior<br />
distal angle with RS, inferior distal angle with 7 RS;<br />
propodus 0.4 as long as ischium, 3.5 times as long as<br />
wide, inferior margin with 2 RS (set singly), superior<br />
distal angle with 3 slender setae ( palmate), inferior<br />
distal angle with 3 RS.<br />
93
Figure 126. Rocinela satagia sp. nov. A, C, D, H–I, holotype, remainder paratype 8.5 mm. A, dorsal view; B, dorsal view,<br />
paratype; C, head, dorsal view; D, frons; E, lateral view; F, pleonites 4 and5, lateral margins; G, head, lateral view; I, sternite 7<br />
showing penial papillae; J antenna peduncle; K, antennule.<br />
94
Figure 127. Rocinela satagia sp. nov. Paratype 8.5 mm. A, mandible; B, mandible molar and incisor; C, mandible palp<br />
article 3; D, maxillule; E, maxillule apex; F, maxilla; G, maxilla apex; H, maxilliped palp articles –3; I, maxilliped scales.<br />
Penes low tubercles; penial openings separated by<br />
5% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod 2.4 times as long as wide, lateral<br />
margin weakly convex, mesial margin weakly convex,<br />
with PMS on distal two-thirds; endopod 2.9 times as<br />
long as wide, lateral margin straight; peduncle mesial<br />
margin with 6 coupling hooks. Pleopod 2 appendix<br />
masculina with sinuate margins, 0.8 times as long as<br />
endopod, distally acute. Pleopods 2–5 peduncle distolateral<br />
margin without acute RS.<br />
Uropod peduncle posterior lobe about one-half as<br />
long as endopod. Exopod at angle <strong>of</strong> about 35° to endopod,<br />
rami extending to pleotelson apex, marginal<br />
setae in two tiers. Endopod lateral margin weakly con-<br />
vex, with –3 RS, mesial margin straight or distally<br />
rounded, with 3 RS. Exopod not extending to end <strong>of</strong><br />
endopod, 2.6 times as long as greatest width; lateral<br />
margin weakly convex, with 9 RS; mesial margin convex,<br />
with 0 RS; distal margin rounded.<br />
size: Males 9–25.0 mm; ovigerous females 9–24 mm,<br />
non-ovigerous females 3–2 mm; mancas 7.5– 2<br />
mm.<br />
Variation: Pleotelson frequently damaged or rubbed<br />
(n = 8): RS 9– 0, with 0 most frequent at 50%. Uropod<br />
exopod lateral margin (n = 23) with 8 (8%), 9 (70%) or<br />
0 (22%) RS; mesial margin (n = 24) without RS with<br />
95
Figure 128. Rocinela satagia sp. nov. A, B, and D holotype, remainder paratype 8.5 mm. A–C, pereopods , 2, and 7, respectively;<br />
D, pereopod , propodal blade; E, pereopod 7, distal margin <strong>of</strong> carpus, mesial RS; F, pleonites 4 and 5, lateral margins.<br />
one instance <strong>of</strong> RS. Uropod endopod mesial margin<br />
(n = 23) varied from –4 RS with 3 (57%) and 4 (35%)<br />
most frequent, and 2 each occurring once; lateral<br />
margin (n = 24) with 3 (92%) RS most frequent, 4 occurring<br />
twice.<br />
<strong>The</strong> setation <strong>of</strong> the palms <strong>of</strong> pereopods –3 is highly<br />
consistent with 5 RS being the most frequent; pereopod<br />
palm (n = 23) with 5 (70%) or 6 (30%), pereopod 2<br />
palm (n = 24) with 5 (75%) or 6 (25%) and pereopod 3<br />
(n = 22) palm 5 (86%) or 6 (9%), 4 occurring once. <strong>The</strong><br />
robust setae on the inferior margin <strong>of</strong> the merus was<br />
consistently +2. <strong>The</strong>re is considerable variation in the<br />
presence <strong>of</strong> setae on the face <strong>of</strong> the propodal blade,<br />
some specimens having only one seta (as is common<br />
to nearly all species <strong>of</strong> the genus) other with a mass <strong>of</strong><br />
96<br />
setae; the differences do not seem to be connected with<br />
the sex or size <strong>of</strong> the specimens.<br />
<strong>The</strong>re is no discernable difference in number <strong>of</strong><br />
robust setae between males and females, nor in relation<br />
to size—the smallest measured here (a manca)<br />
had similar counts to adults for robust setae. <strong>The</strong><br />
characteristic ornamentation <strong>of</strong> the dorsal surface <strong>of</strong><br />
the head is most developed in larger specimens, both<br />
males and females.<br />
remarks: Rocinela satagia sp. nov. can be identified<br />
by the following combination <strong>of</strong> characters: rostrum<br />
broad, strongly produced, eyes narrowly separated,<br />
adult specimens with prominent ridge along the posterior<br />
mesial margin <strong>of</strong> each eye, pereopods –3 with
Figure 129. Rocinela satagia sp. nov. A, B, E and F holotype, remainder paratype 8.5 mm. A–D, pleopods –3, 5 respectively;<br />
E, uropod; F, uropod exopod, ventral view; G, uropod exopod distal margin.<br />
5 or 6 robust setae on propodal blade and, in adult<br />
specimens, propodal blade with numerous setae.<br />
<strong>The</strong> most similar species is Rocinela garricki, which<br />
also has numerous setae on the propodal blade <strong>of</strong> pereopods<br />
–3. That species being readily distinguished<br />
from R. satagia by the far more widely separated eyes,<br />
a greater number <strong>of</strong> robust setae on the blade <strong>of</strong> pereopods<br />
–3 (8 v. 5 in R. satagia), and having propor-<br />
tionally wider uropodal rami, the exopod <strong>of</strong> which in<br />
R. garricki has a distinct distal point.<br />
distribution: Primarily <strong>of</strong>f the eastern coast <strong>of</strong> South<br />
Island from the Cook Strait to <strong>of</strong>f the Otago coast, and<br />
eastwards on the Chatham Rise; 330 to 584 metres.<br />
etymoLogy: Adapted from the Latin satagius (anxious,<br />
worried) alluding the ‘worry lines’ between the eyes.<br />
97
Rocinela sp.<br />
materiaL examined: ♂ (~28 mm), vicinity <strong>of</strong> Bounty<br />
Islands, 47°30’S, 78°45’E, 2 March 973, 39 m, stn<br />
I705 (NIWA 23846).<br />
remarks: This specimen is similar to Rocinela leptopus<br />
sp. nov., but differs in having a short rostrum with an<br />
upturned anterior margin, pereopods –3 with three<br />
prominent robust setae on the palm, each seta set on<br />
a small lobe giving the inferior margin an irregular<br />
appearance. <strong>The</strong> single specimen lacks evident penial<br />
openings, but pleopod 2 has a short appendix masculina.<br />
<strong>The</strong> specimen is intact, and an undescribed<br />
species, but the pleon, pleotelson, posterior pereopods<br />
and uropods are all badly crushed, so the specimen is<br />
unsuitable for description.<br />
Rocinela sp.<br />
materiaL examined: ♀ (15 mm, non-ovig.), Thompson<br />
Sound, Fiordland, South Island, 45° 3.00’S, 66°57.96’E,<br />
28 May 997, 350 m, gravel, sand, coral and mud, coll.<br />
RV Munida (NMNZ Cr. 20 ).<br />
remarks: <strong>The</strong> single specimen is most similar to Rocinela<br />
satagia sp. nov., but differs notably in having eyes that<br />
meet medially and the posterior margin <strong>of</strong> the pleotelson<br />
being narrowed. <strong>The</strong> setation <strong>of</strong> the uropods is<br />
similar to that <strong>of</strong> R. satagia but the propodal blade <strong>of</strong><br />
pereopods –3 all have only four acute robust setae.<br />
Genus Syscenus Harger, 880<br />
Syscenus Harger, 880: 387.– Sars, 897: 66.– Richardson,<br />
905a: 2 2.– Stebbing, 924: 9.– Wahrberg, 930:<br />
24.– Nierstrasz & Schuurmans Steckhoven jr., 930:<br />
77.– Schultz, 969: 96.– Menzies & George, 972:<br />
2.– Kussakin, 979: 269.– Bruce, 997: 3.– Bruce,<br />
Lew Ton & Poore, 2002: 63.<br />
Harponyx Sars, 882: 60 (type species Harponyx pranizoides<br />
Sars, 882).<br />
Rocinela.– Bovallius, 885 (not Rocinela Leach, 8 8).<br />
Syscénus.– Stephenson, 948: 4 .<br />
type speCies: Syscenus infelix Harger, 880; by<br />
monotypy.<br />
diagnosis: Body dorsally vaulted. Head laterally free <strong>of</strong><br />
pereonite ; rostral point weak; eyes absent or present.<br />
Pleonite abruptly narrower than pereonite 7. Frontal<br />
lamina present, slender, elongate. Maxilliped palp<br />
2- or 3-articled. Uropodal peduncle mesial margin<br />
not produced; rami lamellar. Coxae 5–7 shorter than<br />
respective pereonite.<br />
desCription: Body elongate, 3 to 4 times as long as<br />
wide. Head anterior margin with small median (ros-<br />
98<br />
tral) point. Eyes usually absent (present in two species).<br />
Coxae <strong>of</strong> pereonites 4–7 shorter than respective<br />
segment, not posteriorly produced. Pleon abruptly<br />
narrower than pereon, approx. 30% to 60% maximum<br />
body width; pleonites all visible, all with free lateral<br />
margins; pleonites 3–5 posteriorly produced. Pleotelson<br />
large, as long as or longer than pleon, usually with<br />
blunt or narrow caudomedial point (never acute or<br />
truncate). Pleonal sternite present anterior to pleopod<br />
peduncles.<br />
Antennule short, not exceeding antenna peduncle in<br />
length. Antenna peduncle articles 4 and 5 (or, either 4<br />
or 5) and proximal flagellum provided with long setae<br />
(most species).<br />
Frontal lamina usually present; labrum present.<br />
Mandible with unicuspid incisor; molar process and<br />
spine row absent. Maxillule with terminal RS. Maxilliped<br />
3- or 4-articled, article 3 with 2–3 recurved RS,<br />
article 4 with recurved RS; endite absent.<br />
Pleopod 3 endopod usually without PMS, pleopods<br />
4 and 5 endopods without PMS; endopod 3-5 usually<br />
not distinctly smaller than exopods; coupling setae<br />
present on peduncles <strong>of</strong> pleopods –5; pleopod 5<br />
without proximomesial lobe. Pleopods not extending<br />
beyond lateral margins <strong>of</strong> pleon. Uropods flat, both<br />
rami lamellar, endopod longer than exopod.<br />
remarks: Syscenus is best recognised by the coxae <strong>of</strong><br />
pereonites 5–7 being shorter than the respective pereonite,<br />
pleon evidently narrower than the pereon, pleonite<br />
5 with free (not overlapped) lateral margins, and<br />
lack <strong>of</strong> a dorsal rostrum. Most species lack any trace <strong>of</strong><br />
eyes, but two species, Syscenus karu Bruce, 2005 from<br />
Vanuatu and Syscenus peruanus Menzies and George,<br />
972 from <strong>of</strong>f Peru, do have eyes.<br />
Syscenus is a small genus known from all oceans<br />
except the Southern Ocean. <strong>The</strong>re are six named species<br />
(following S. pacificus Nunomura, 98 being here<br />
placed in synonymy), with a further two unnamed<br />
species from <strong>New</strong> <strong>Zealand</strong> recorded here. Most species<br />
are superficially similar in appearance. It is known that<br />
at least one species, Syscenus infelix Harger, 880, is a<br />
fish predator and possibly more host-dependent (Ross<br />
et al. 200 ) than noted for Aega or Rocinela.<br />
Most species <strong>of</strong> the genus are known from only a<br />
few locations. <strong>The</strong> exception is Syscenus infelix, which<br />
has been recorded from the North Atlantic, Mediterranean,<br />
northern and southwestern Pacific, and South<br />
Africa (Kensley 2004; Kensley & Cartes 2003). Kensley<br />
& Cartes (2003) considered that many <strong>of</strong> the records<br />
for S. infelix were ‘open to doubt’, and Kensley (2004)<br />
more specifically rejected Pacific records <strong>of</strong> S. infelix<br />
as misidentifications, an opinion with which I agree.<br />
However given that some isopod mesopelagic species<br />
are known to have an enormous range (e.g. Svavarsson<br />
& Bruce 2000), and that several species <strong>of</strong> the related
genus Aega (e.g. A. falklandica, A. monophthalma, A. komai<br />
and A. urotoma and others; all this study) also have<br />
extensive ranges, the possibility that some <strong>of</strong> those<br />
records are correct cannot be excluded.<br />
As most species are known from single or a few<br />
specimens, the range and pattern <strong>of</strong> character variation<br />
is not known. Despite the large amount <strong>of</strong> material<br />
recorded by Kensley and Cartes (2003) for Syscenus<br />
infelix and Syscenus atlanticus Kononenko, 988, no assessment<br />
was made <strong>of</strong> character variation in those two<br />
species, although variation in pleotelson and uropod<br />
shape was illustrated by Kensley and Cartes (2003).<br />
Most <strong>of</strong> the remaining species <strong>of</strong> the genus are in need<br />
<strong>of</strong> further revision.<br />
<strong>The</strong> genus is likely to be found in all tropical and<br />
temperate oceans, and has been recorded from depths<br />
as shallow as 70 metres in the North Atlantic (Kensley<br />
2004) to 4609 metres <strong>of</strong>f northern Peru (Menzies &<br />
George 972); most records are between approximately<br />
500 and 2000 metres.<br />
Key to the new <strong>Zealand</strong> species <strong>of</strong> syscenus<br />
. Pereopods with blunt RS and numerous slender<br />
setae; pleotelson with caudomedial point ............<br />
............................................S. springthorpei (p. 208)<br />
— Pereopods without numerous slender setae, without<br />
prominent blunt RS; pleotelson posteriorly<br />
rounded or subtruncate ........................................2<br />
2. Body wide, ovate; pleonite 5 with dorsal median<br />
spine (female) or point (male); rostrum weak,<br />
blunt ................................................ S. latus (p. 202)<br />
–- Body elongate, margins subparallel; pleonite 5<br />
without point or spine; rostrum narrow, projecting<br />
anteriorly, then ventrally ...............................3<br />
3. Antennal flagellum extending to pereonite 3;<br />
uropodal rami without RS .....................................<br />
.............................................Syscenus kapoo (p. 99)<br />
–- Antennal flagellum extending to pereonite 6;<br />
uropodal rami with RS ...........................................<br />
...........................................Syscenus moana (p. 206)<br />
Syscenus kapoo sp. nov. (Figs 30, 3 )<br />
materiaL examined: Holotype: ♀ (non-ovig. 2 mm),<br />
Norfolk Ridge, 26°25.94’S, 67° 0.87’E, 8 May 2003,<br />
750–774 m, NORFANZ (NIWA 23780).<br />
desCription: Body 2.8 times as long as greatest width,<br />
dorsal surfaces smooth, widest at pereonite 5, lateral<br />
margins subparallel. Eyes absent. Rostrum simple or<br />
anteriorly narrow. Coxae 2–3 each with posteroventral<br />
angle with small distinct produced point; 5–7 without<br />
oblique carina. Pleon with pleonite largely concealed<br />
by pereonite 7; pleonite 4 with posterolateral margins<br />
not extending to posterior margin <strong>of</strong> pleonite 5; pleonite<br />
5 with posterolateral angles rounded. Pleotelson<br />
.0 as long as anterior width, anterior dorsal surface<br />
with 2 sub-median depressions (weak), dorsal surface<br />
smooth; lateral margins convex, posterior margin narrowly<br />
rounded, with 0 RS.<br />
Antennule peduncle as for the genus; flagellum with<br />
3 articles, extending to middle <strong>of</strong> pereonite . Antenna<br />
peduncle as for the genus; flagellum with 35 articles,<br />
extending to middle <strong>of</strong> pereonite 3.<br />
Frontal lamina longer than greatest width, anteriorly<br />
acute.<br />
Mouthparts as for the genus.<br />
Pereopod 1 basis 2.4 times as long as greatest width;<br />
ischium 0.6 times as long as basis, inferior margin with 0<br />
RS, superior distal margin with 0 setae (4 simple and 2<br />
plumose setae); merus inferior margin with 0 RS, superior<br />
distal angle with 2 setae; carpus .3 times as long as<br />
merus, inferior margin with 0 RS; propodus .7 times as<br />
long as proximal width, propodal palm simple, without<br />
blade or process, without setae, inferior margin with 0<br />
RS; dactylus .4 times as long as propodus. Pereopods 2<br />
and 3 similar to pereopod . Pereopod 6 similar to pereopod<br />
7. Pereopod 7 basis 3.5 times as long as greatest<br />
width, inferior margins with 5 palmate setae; ischium<br />
0.9 as long as basis, inferior margin with 0 RS, superior<br />
distal angle with 6 RS (and seta), inferior distal angle<br />
with 4 RS; merus 0.6 times as long as ischium, 2.3 times<br />
as long as wide, inferior margin with 5 RS (set as 2, ,<br />
and ), superior distal angle with 22 RS (in two ranks<br />
<strong>of</strong> 6 major and ~ 6 slender), inferior distal angle with<br />
6 RS; carpus .0 as long as ischium, 6.4 times as long as<br />
wide, inferior margin with 8 RS (set as 2, , 3, and ),<br />
superior distal angle with 25 RS (in two ranks <strong>of</strong> 7 major<br />
and ~ 8 slender and robust setae), inferior distal angle<br />
with 6 RS; propodus 0.9 as long as ischium, 7.3 times<br />
as long as wide, inferior margin with 0 RS (2 minute<br />
submarginal), superior distal angle with 0 slender<br />
setae, inferior distal angle with 2 RS. Pereopods distal<br />
margins <strong>of</strong> ischium to carpus without setae; without<br />
strong carina on basis.<br />
Pleopods 1 as for the genus.<br />
Uropod peduncle ventrolateral margin with 0 RS,<br />
posterior lobe about one-third as long as endopod.<br />
Uropod rami with endopod and exopod co-planar,<br />
rami extending beyond pleotelson, marginal setae in<br />
single tier. Endopod lateral margin weakly convex, lateral<br />
margin with 0 RS, mesial margin straight, with 0<br />
RS. Exopod not extending to end <strong>of</strong> endopod, 2.8 times<br />
as long as greatest width; lateral margin convex, with 0<br />
RS; mesial margin sinuate, proximally concave, distally<br />
convex, with 0 RS; distal margin rounded.<br />
remarks: <strong>The</strong> single specimen, though adult, is <strong>of</strong> uncertain<br />
maturity, and could not be identified as any <strong>of</strong><br />
99
Figure 130. Syscenus kapoo sp. nov. Holotype. A, dorsal view; B, lateral view; C, head, dorsal view; D, frons; E, pleotelson<br />
and uropods; F, pleonites, lateral view; G, uropod exopod, ventral view; H, uropod.<br />
200
Figure 131. Syscenus kapoo sp. nov. Holotype. A–C, pereopods , 2 and 7 respectively; D, mesial margin <strong>of</strong> merus; E, mesial<br />
margin <strong>of</strong> carpus.<br />
the five other species known from the southwestern<br />
Pacific. Syscenus kapoo sp. nov. can be identified by the<br />
shape <strong>of</strong> the head which has convex lateral margins<br />
and a relatively weak rostrum, the acute coxae, evenly<br />
20<br />
rounded pleotelson posterior margin, antennal flagellum<br />
extending to pereonite 3, elongate pereopods 5–7<br />
with pereopod 7 extending posteriorly beyond the<br />
posterior margin <strong>of</strong> the pleotelson, and the uropods
which extend beyond the pleotelson, the exopod <strong>of</strong><br />
which is shorter than the endopod and is proximally<br />
narrow.<br />
Comparison with the holotype <strong>of</strong> S. intermedius (see<br />
Appendix 2) shows that, in S. intermedius, the frontal<br />
lamina is narrower, the posterior legs are more robust,<br />
the uropod rami are roughly subequal in length, the<br />
coxae are posteriorly rounded and the pleotelson<br />
lateral margin has a distinct inflexion. Syscenus kapoo<br />
is also similar to the potentially sympatric S. moana,<br />
from which it differs in having a far shorter antennal<br />
flagellum (to pereonite 3 v. pereonite 6 in S. moana),<br />
the robust setae on the merus and carpus <strong>of</strong> pereopod<br />
7 are larger, and the uropods differ in lacking robust<br />
setae, the endopod being more slender, and the exopod<br />
is shorter than that <strong>of</strong> S. moana, and different in shape<br />
being wider distally as well as narrower proximally.<br />
prey: Not known.<br />
etymoLogy: <strong>The</strong> epithet is a Mäori word meaning blind<br />
or without sight.<br />
distribution: Known only from the type locality, on<br />
the Norfolk Ridge.<br />
Syscenus latus Richardson, 909 (Figs 32– 34)<br />
Syscenus latus Richardson, 1909: 85, fig. 11.– Bruce, 1997:<br />
4.– Saito, Itani & Nunomura, 2000: 6 .<br />
Syscenus pacificus Nunomura, 98 : 5, fig .– Bruce,<br />
997: 4.– Saito, Itani & Nunomura, 2000: 6 (new<br />
synonymy).<br />
materiaL examined: Lectotype (here designated): ♀ (ovig.<br />
42 mm), at Tsurikake Saki Light, <strong>of</strong>f Koshika Islands,<br />
Sea <strong>of</strong> Japan, Japan, 3 °39.5’N, 29°24.0’E, August<br />
906, 742 metres (USNM 39502). Paralectotype: ♂? ( 9<br />
mm), at Tsurikake Saki Light, <strong>of</strong>f Koshika Islands, Sea<br />
<strong>of</strong> Japan, Japan, 3 °39.0’N, 29°20.5’E, August 906,<br />
742 metres (USNM 39906 — former syntype).<br />
<strong>New</strong> <strong>Zealand</strong> specimen: ♀ (non-ovig. 38 mm), Challenger<br />
Plateau, 40° 9.65’S, 70° 3.80’E, 9 March 98 ,<br />
805–822 m, RV James Cook (NMNZ Cr. 20 2).<br />
Additional material: <strong>New</strong> caledonia, HALIPRO<br />
2, coll. B. Richer de Forges: (27 mm, non-ovig, sex<br />
uncertain), 23°59’S, 6 °55’E, 25 November 996, stn.<br />
BT96, 034– 056 m (MNHN Is.5881); (2 mm, nonovig,<br />
sex uncertain), 24°00’S, 6 °49’E, 25 November<br />
996, stn. BT97, 964– 03 m (MNHN Is.5882). Indonesia,<br />
KARUBAR: ♀ (33 mm, non-ovig.), region <strong>of</strong> Kei<br />
and Tanimbar Islands, Banda Sea, 05° 4’S, 33°00’E,<br />
25 October 99 , stn. CP2 688–694, coll. Baruna Jaya<br />
(MNHN Is.5883).<br />
desCription (based on lectotype and <strong>New</strong> <strong>Zealand</strong><br />
female): Body 2. times as long as greatest width,<br />
202<br />
dorsal surfaces smooth, widest at pereonite 5, lateral<br />
margins ovate. Eyes absent. Rostrum simple, anteriorly<br />
subtruncate. Coxae 2–3 each with posteroventral<br />
angle rounded; 5–7 without oblique carina. Pleon with<br />
pleonite visible in dorsal view; pleonite 4 with posterolateral<br />
margins not extending to posterior margin <strong>of</strong><br />
pleonite 5; pleonite 5 with posterolateral angles acute<br />
(in dorsal view). Pleotelson .2 times as long as anterior<br />
width, dorsal surface smooth; lateral margins convex,<br />
posterior margin evenly rounded, with 0 RS.<br />
Antennule peduncle article 3 0.9 times as long as<br />
combined lengths <strong>of</strong> articles and 2, 3.7 times as long<br />
as wide; flagellum with 14 articles, extending to pereonite<br />
2. Antenna peduncle article 3 .0 times as long<br />
as article 2, .3 times as long as wide; article 4 2.4 times<br />
as long as article 3, 3.0~ times as long as wide, inferior<br />
margin with 2 plumose setae (probably simple, but<br />
may have dried at some point); article 5 .2 times as<br />
long as article 4, 4.0 as long as wide, inferior margin<br />
with 0 setae, anterodistal angle with cluster <strong>of</strong> 0 short<br />
simple setae; flagellum with 32 articles (articles 2–15<br />
with conspicuous cluster <strong>of</strong> setae at distal angle), extending<br />
to pereonite 6.<br />
Frontal lamina longer than greatest width, anteriorly<br />
acute.<br />
Mandible molar process present, minute; palp article<br />
2 with ~30 marginal distolateral setae (setae multitiered);<br />
palp article 3 with 29 setae (distal 2 longest;<br />
marginal setae irregularly spaced). Maxillule with 6 RS<br />
(2 large, 4 slender). Maxilla mesial lobe with hooked<br />
RS (weakly hooked); lateral lobe with 2 hooked RS.<br />
Maxilliped palp article 3 with 2 hooked RS; article 3.<br />
Pereopod 1 basis 2.9 times as long as greatest width;<br />
ischium 0.3 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 0 setae; merus inferior<br />
margin with 0 RS, superior distal angle with 4 setae;<br />
carpus 0.3 times as long as merus, inferior margin with<br />
0 RS; propodus 2.4 times as long as proximal width, propodal<br />
palm simple, without blade or process, without<br />
setae, inferior margin with 0 RS; dactylus .6 times as<br />
long as propodus. Pereopod 6 similar to pereopod 7.<br />
Pereopod 7 basis 3.3 times as long as greatest width, inferior<br />
margins with 0 palmate setae; ischium 0.8 as long<br />
as basis, inferior margin with 0 RS, superior distal angle<br />
with RS (and 2 simple setae), inferior distal angle with<br />
0 RS; merus 0.8 times as long as ischium, 3.4 times as<br />
long as wide, inferior margin with 7 RS (submarginal;<br />
short, slender, acute), superior distal angle with 0 RS<br />
(with about 6 slender setae in several tiers), inferior<br />
distal angle with 8 RS (acute); carpus 0.95 times as long<br />
as ischium, 5.3 times as long as wide, inferior margin<br />
with RS (minute, submarginal), superior distal angle<br />
with several; setae missing, inferior distal angle with 5<br />
RS; propodus .2 as long as ischium, 8 times as long as<br />
wide, inferior margin with 2 RS (minute; submarginal),<br />
superior distal angle with 0 slender setae (possibly
Figure 132. Syscenus latus Richardson, 909. A–E, holotype, remainder NMNZ Cr. 20 2. A, dorsal view; B, lateral view;<br />
C, head, dorsal view; D, frons; E, pleonites, lateral view; F, antenna; G, antennule; H, antenna peduncle; I, dorsal view;<br />
J, frons.<br />
203
Figure 133. Syscenus latus Richardson, 909. NMNZ Cr. 20 2. A, mandible; B, mandible incisor; C, mandible palp articles 2<br />
and 3; D, maxillule; E, maxillule apex; F, maxilla; G, maxilla apex; H, maxilliped; I, maxilliped palp article 2.<br />
missing), inferior distal angle with RS. Pereopods<br />
distal margins <strong>of</strong> ischium to carpus without setae;<br />
without strong carina on basis.<br />
Pleopod 1 exopod 2.3 times as long as wide, lateral<br />
margin straight, mesial margin weakly convex, with<br />
PMS on distal margin only; endopod 2.8 times as<br />
long as wide, lateral margin straight, with PMS on<br />
distal margin only, mesial margin with PMS on distal<br />
two-thirds; peduncle mesial margin with coupling<br />
hooks. Pleopods 2–5 peduncle distolateral margin each<br />
without acute RS.<br />
Uropod peduncle ventrolateral margin with 0 RS,<br />
posterior lobe about absent. Uropod rami with endopod<br />
and exopod co-planar, rami extending beyond<br />
pleotelson. Endopod lateral margin convex, lateral<br />
margin with 0 RS, mesial margin straight, with 0 RS.<br />
Exopod not extending to end <strong>of</strong> endopod, 3.3 times as<br />
204<br />
long as greatest width; lateral margin convex, with 0<br />
RS; mesial margin sinuate, proximally concave, distally<br />
convex, with 0 RS.<br />
maLe: No male had been positively identified.<br />
size: From 9–42 mm; Nunomura ( 98 ) recorded<br />
females up to 46 mm.<br />
Variation: <strong>The</strong> <strong>New</strong> <strong>Zealand</strong> specimen has a frontal<br />
lamina, more strongly defined than in the lectotype,<br />
but similar to that <strong>of</strong> the paralectotype. <strong>The</strong> antennule<br />
flagellum extends to the middle <strong>of</strong> pereonite 2, the<br />
antennal flagellum to pereonite 6.<br />
remarks: Syscenus latus is readily identified by the<br />
prominent ocular lobes, pleonite 4 posterior margin
Figure 134. Syscenus latus Richardson, 909. NMNZ Cr. 20 2 except J, holotype. A–C, pereopods , 2 and 7 respectively;<br />
D, mesial margin <strong>of</strong> merus; E–H, pleopods , 2, 4 and 5 respectively; I, pleopod peduncle, mesial margin; J, uropod<br />
(holotype), in situ.<br />
205
with a distinct median point, pleonite 5 with a distinct<br />
median point or in the ovigerous female a short dorsally<br />
directed spine, slender dactylus on pereopods<br />
–3, slender distal articles on pereopods 5–7, all pereopods<br />
having few and small robust setae, the broadly<br />
rounded pleotelson posterior margin and the broadly<br />
rounded uropodal rami. No other species has an acute<br />
median point or dorsally directed spine on the posterior<br />
margin <strong>of</strong> pleonite 5.<br />
Nunomura ( 98 ), when describing Syscenus pacificus<br />
clearly believed that he was describing the second<br />
species <strong>of</strong> the genus, stating that ‘the genus Syscenus<br />
had hitherto been represented by the single species<br />
S. infelix’. Bruce ( 997a) questioned the validity <strong>of</strong><br />
S. pacificus. Examination <strong>of</strong> the type material <strong>of</strong><br />
Syscenus latus, also from Japan, and comparison with<br />
the description given by Nunomura ( 98 ) now confirm<br />
the synonymy.<br />
<strong>The</strong> large female is designated as lectotype. <strong>The</strong><br />
smaller <strong>of</strong> the two syntypic specimens, both from the<br />
same locality, is in poor condition, with the posterior<br />
half appearing decomposed, and remains as paralectotype.<br />
While pleonite 5 has a short spine and the frons,<br />
antennule, pleotelson and uropods are similar to those<br />
<strong>of</strong> the larger specimen, the antennal flagellum is shorter<br />
in length, extending to pereonite 4 (rather than 6).<br />
prey: Not known.<br />
distribution: In <strong>New</strong> <strong>Zealand</strong> known only from the<br />
single specimen from the Challenger Plateau to the<br />
west <strong>of</strong> northern North Island; here also recorded from<br />
<strong>New</strong> Caledonia, Lord Howe Rise and the Banda Sea,<br />
Indonesia; previous records from Japan; at depths <strong>of</strong><br />
688– 056 metres (all records).<br />
Syscenus moana Bruce, 2005 (Fig. 35)<br />
Syscenus moana Bruce, 2005: 32, figs 1–4.<br />
diagnosis (from Bruce 2005): Body 2.7 times as long as<br />
greatest width. Rostrum anteriorly narrow, anteriorly<br />
truncate in dorsal view (apically bent ventrally). Eyes<br />
absent. Coxae 2 and 3 each with posteroventral angle<br />
with small distinct produced point; 5–7 without oblique<br />
carina. Pleonite 4 with posterolateral margins not<br />
extending to posterior margin <strong>of</strong> pleonite 5; pleonite 5<br />
with posterolateral angles acute. Pleotelson 0.9 times as<br />
long as anterior width, anterior dorsal surface without<br />
2 sub-median depressions; lateral margins convex,<br />
posterior margin evenly rounded, without RS.<br />
Antennule flagellum with 8 articles, extending to<br />
anterior <strong>of</strong> pereonite . Antenna flagellum with 32<br />
articles, extending to middle <strong>of</strong> pereonite 6.<br />
Frontal lamina wider than long, anteriorly acute.<br />
206<br />
Pereopod 1 basis 2. times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin<br />
without RS, superior distal margin with 2 simple setae;<br />
merus inferior margin without RS, superior distal angle<br />
with simple setae; carpus .0 as long as merus, inferior<br />
margin without RS; propodus .3 times as long as<br />
proximal width, propodal palm simple, without blade<br />
or process, without setae, inferior margin without RS;<br />
dactylus .8 times as long as propodus. Pereopod 7 basis<br />
2.7 times as long as greatest width, inferior margins<br />
with 2 palmate setae; ischium 0.9 as long as basis,<br />
inferior margin without RS, superior distal angle with<br />
RS, inferior distal angle with 5 RS; merus 0.6 times<br />
as long as ischium, 2.0 as long as wide, inferior margin<br />
with 8 RS (set as 2, 2, 2, and ), superior distal angle<br />
with 9 RS, inferior distal angle with 6 RS; carpus 0.8<br />
times as long as ischium, 3.0 as long as wide, inferior<br />
margin with 8 RS (set as 2, , 2, , and ), superior<br />
distal angle with 20 RS, inferior distal angle with 5<br />
RS; propodus .0 as long as ischium, 5.7 times as long<br />
as wide, inferior margin with 3 RS (very small, submarginal),<br />
superior distal angle with 3 slender setae<br />
(plumose), inferior distal angle with RS. Pereopods<br />
distal margins <strong>of</strong> ischium to carpus without abundant<br />
simple setae; without strong carina on basis.<br />
Penes opening flush with surface <strong>of</strong> sternite 7.<br />
Pleopod 1 exopod 2.0 as long as wide, lateral margin<br />
straight, mesial margin strongly convex, with PMS on<br />
distal one-third; endopod 2.5 times as long as wide, lateral<br />
margin weakly convex, with PMS on distal margin<br />
only, mesial margin with PMS on distal three-quarters.<br />
Pleopod 2 appendix masculina with straight margins, 0.5<br />
times as long as endopod, distally bluntly rounded.<br />
Uropod peduncle ventrolateral margin with RS,<br />
posterior lobe absent. Uropod rami with endopod and<br />
exopod co-planar, rami extending beyond pleotelson.<br />
Endopod lateral margin weakly convex, distolateral<br />
margin with 3 RS, mesial margin straight, without RS.<br />
Exopod not extending to end <strong>of</strong> endopod, 3.9 times as<br />
long as greatest width; lateral margin convex, with 8<br />
RS; mesial margin evenly concave, without RS; distal<br />
margin rounded.<br />
remarks: Syscenus moana can be recognised by the acute<br />
coxae, antennal flagellum extending to pereonite 6 (not<br />
pereonite 3 as stated incorrectly in the original description),<br />
rounded margin to the pleotelson and uropods<br />
with robust setae.<br />
distribution: Single record from northern Norfolk<br />
Ridge. Species <strong>of</strong> Syscenus are mesopelagic fish predators<br />
or parasites, and it is quite likely that this species<br />
will occur more widely in northern <strong>New</strong> <strong>Zealand</strong><br />
waters.
Figure 135. Syscenus moana Bruce, 2005. A, dorsal view; B, head, dorsal view; C, frons; D, pleotelson and uropods, dorsal<br />
view; E, antennule; pereopod ; F, pereopod ; G, uropod.<br />
207
Syscenus springthorpei Bruce, 997 (Fig. 36)<br />
Syscenus springthorpei Bruce, 1997: 114, figs 1–4.– Bruce, Lew<br />
Ton & Poore, 2002: 63.<br />
materiaL examined: ♂? (39 mm), Tui Oceanographic<br />
Cruise, Auckland University Zoology, AUZ 098 4 ,<br />
locality not known, probable <strong>New</strong> <strong>Zealand</strong> EEZ;<br />
previously dissected (label in Hurley’s handwriting:<br />
“ex Kussakin: 23 , 269”); dissected appendages in fair<br />
condition but specimen seems to have subsequently<br />
deteriorated (NIWA 2378).<br />
desCription (after Bruce 997a): Body 3.0 as long as<br />
greatest width, dorsal surfaces smooth, widest at pereonite<br />
5, lateral margins subparallel. Eyes absent. Rostrum<br />
simple, anteriorly rounded. Coxae 2–3 each with<br />
posteroventral angle rounded; 5–7 without oblique<br />
carina. Pleon with pleonite visible in dorsal view;<br />
pleonite 4 with posterolateral margins not extending<br />
to posterior margin <strong>of</strong> pleonite 5; pleonite 5 with posterolateral<br />
angles acute. Pleotelson .2 times as long as<br />
anterior width, dorsal surface smooth; lateral margins<br />
weakly convex, posterior margin with distinct median<br />
point, with 0 RS.<br />
Antennule peduncle article 3 0.6 times as long as<br />
combined lengths <strong>of</strong> articles and 2, .6 times as<br />
long as wide; flagellum with 7 articles, extending to<br />
anterior <strong>of</strong> pereonite . Antenna peduncle article 3 2.5<br />
times as long as article 2, .5 times as long as wide;<br />
article 4 .3 times as long as article 3, .8 times as long<br />
as wide, inferior margin with 3 plumose setae (long),<br />
and 0 simple setae; 2.7 times as long as wide, inferior<br />
margin with 10 setae (long, plumose); flagellum with<br />
20 articles (approximately), extending to posterior <strong>of</strong><br />
pereonite 3.<br />
Frontal lamina longer than greatest width, anteriorly<br />
acute.<br />
Mandible molar process absent; palp article 2 with<br />
2 marginal distolateral setae, and 3 long distolateral<br />
setae; palp article 3 with 9 setae. Maxillule with 5 RS (4<br />
large, apically curved, slender, straight). Maxilla mesial<br />
lobe with 0 hooked RS; lateral lobe with 2 hooked<br />
RS. Maxilliped palp article distomesial angle with<br />
RS; article 2 with 3 hooked RS; article 3 with hooked<br />
RS (and two short simple setae).<br />
Pereopod 1 basis 2.3 times as long as greatest width;<br />
ischium 0.4 times as long as basis, inferior margin with<br />
0 RS, superior distal margin with 5 setae; merus inferior<br />
margin with RS (large, distal), superior distal angle<br />
with 2 setae; carpus .0 as long as merus, inferior margin<br />
with RS (with numerous simple setae); propodus<br />
.6 times as long as proximal width, propodal palm<br />
simple, without blade or process, inferior margin with<br />
RS (distal); dactylus .3 times as long as propodus.<br />
Pereopods 2 and 3 similar to pereopod . Pereopod 6<br />
208<br />
similar to pereopod 7. Pereopod 7 basis 2.4 times as<br />
long as greatest width; ischium 0.8 as long as basis,<br />
inferior margin with 0 RS, superior distal angle with 2<br />
RS (and numerous simple setae), inferior distal angle<br />
with 4 RS (and numerous simple setae); merus 0.6 times<br />
as long as ischium, .7 times as long as wide, inferior<br />
margin with 2 RS, superior distal angle with 6 RS (and<br />
~5 simple setae), inferior distal angle with 4 RS (and<br />
~4 simple setae); carpus .3 times as long as ischium,<br />
2.7 times as long as wide, inferior margin with 2 RS,<br />
superior distal angle with 8 RS, inferior distal angle<br />
with 4 RS; propodus 0.9 as long as ischium, 3.7 times<br />
as long as wide, inferior margin with RS, superior<br />
distal angle with 0 slender setae, inferior distal angle<br />
with RS. Pereopods distal margins <strong>of</strong> ischium to<br />
carpus with abundant simple setae; without strong<br />
carina on basis.<br />
Penes low tubercles; penial openings separated by<br />
% <strong>of</strong> sternal width.<br />
Pleopod 1 exopod 2.0 as long as wide, lateral margin<br />
weakly convex, mesial margin weakly convex, with<br />
PMS on distal three-quarters; endopod 2.5 times as<br />
long as wide, lateral margin weakly convex, with PMS<br />
on distal margin only, mesial margin with PMS on<br />
distal three-quarters; peduncle mesial margin with 9<br />
coupling hooks. Pleopod 2 appendix masculina distally<br />
narrow, 0.9 times as long as endopod, distally acute.<br />
Pleopods 2–5 peduncle distolateral margin each with<br />
acute RS.<br />
Uropod peduncle ventrolateral margin with 0 RS,<br />
posterior lobe about one-third as long as endopod.<br />
Uropod rami with endopod and exopod co-planar,<br />
rami not extending to pleotelson apex. Endopod lateral<br />
margin weakly convex, lateral margin with 0 RS, mesial<br />
margin weakly convex, with 0 RS. Exopod not extending<br />
to end <strong>of</strong> endopod, 3.7 times as long as greatest width;<br />
lateral margin weakly convex, with 0 RS; mesial margin<br />
convex, with 0 RS; distal margin rounded.<br />
femaLe: No female had been positively identified.<br />
size: Holotype 36 mm; <strong>New</strong> <strong>Zealand</strong> specimen 39<br />
mm.<br />
Variation: <strong>The</strong> <strong>New</strong> <strong>Zealand</strong> specimen is in poor condition<br />
and meaningful assessment is not possible.<br />
remarks: <strong>The</strong> single specimen is in very poor condition,<br />
appearing to have dried out at some point after it was<br />
dissected (the dissected appendages are in adequate<br />
condition) and also having suffered a mould infection.<br />
In addition, the locality <strong>of</strong> the specimen is not known,<br />
although, having been collected by the RV Tui, it will<br />
have been within the <strong>New</strong> <strong>Zealand</strong> region. For these<br />
reasons the descriptions and figures are taken from<br />
Bruce ( 997a).
Figure 136. Syscenus springthorpei Bruce, 997. A, dorsal view; B, head, dorsal view; C, frons; D, pereopod E, pereopod 2<br />
(distal articles); F, pereopod 7; G, antenna peduncle; H, antennule; I, uropod, in situ, dorsal view; J, pleotelson and uropods,<br />
ventral; view.<br />
Syscenus springthorpei can be identified by the relatively<br />
slender body shape, antennal flagellum extending<br />
to the posterior <strong>of</strong> pereonite 3, posterior margin<br />
<strong>of</strong> the pleotelson having a distinct median point, the<br />
uropodal rami not extending to the posterior margin<br />
209<br />
<strong>of</strong> the pleotelson, and the robust and setose pereopods<br />
that have conspicuous, blunt robust setae on the inferior<br />
margins.<br />
Characters separating this species from Syscenus<br />
infelix Harger, 880 are the pleotelson margins being
smoothly curved rather than sinuate (as figured by<br />
Harger 883), the presence <strong>of</strong> a blunt distinct caudomedial<br />
point (v. acute), and the uropods not extending<br />
beyond the posterior margin <strong>of</strong> the pleotelson (v.<br />
extending beyond the posterior <strong>of</strong> the pleotelson).<br />
<strong>The</strong> figures given by Kensley and Cartes (2003) show<br />
considerable variation in these characters, some <strong>of</strong><br />
which appear similar to those <strong>of</strong> S. springthorpei, and<br />
furthermore the shape <strong>of</strong> the uropodal rami is near<br />
identical in both species. <strong>The</strong> pereopods differ in<br />
S. springthorpei having robust setae on the inferior<br />
margins <strong>of</strong> pereopods 4–7, these being absent in<br />
S. infelix (Kensley 2004).<br />
<strong>The</strong> type locality for S. infelix is Cape Cod, Massachusetts,<br />
northwestern North Atlantic, and without<br />
detailed reassessment <strong>of</strong> S. infelix based on the type<br />
material and from specimens collected geographically<br />
nearby, it is not possible to say if the western Atlantic<br />
species is the same as the eastern Atlantic material<br />
recorded under that name by Kensley and Cartes<br />
(2003), although the figured shapes <strong>of</strong> the pleotelson<br />
<strong>of</strong> western Atlantic and Mediterranean specimens do<br />
differ considerably.<br />
prey: Not known.<br />
distribution: In <strong>New</strong> <strong>Zealand</strong> known only from the<br />
single specimen <strong>of</strong> uncertain locality; previous record<br />
from <strong>of</strong>f <strong>New</strong> South Wales, Australia.<br />
Syscenus sp.<br />
materiaL examined: ♂ (23 mm), east <strong>of</strong> Mahia Peninsula,<br />
29 September 989, 39°40.5–43.5’S, 78°09.2–07.2’E,<br />
764–843 m, coll. RV James Cook (NMNZ Cr. 20 3).<br />
remarks: This specimen has one conspicuous and<br />
unique character — the superior distal angle <strong>of</strong> the<br />
ischium <strong>of</strong> pereopods 4–6 is strongly produced, over-<br />
iding the merus. Other character states include: dactylus<br />
<strong>of</strong> pereopods –3 longer than in S. springthorpei. <strong>The</strong><br />
pleotelson and uropods are damaged, but the uropods<br />
do exceed the posterior margin <strong>of</strong> the pleotelson. <strong>The</strong><br />
anterior <strong>of</strong> the head is ‘short’, the frontal lamina is<br />
anteriorly rounded; the antennule flagellum extends<br />
to anterior <strong>of</strong> pereonite 1, with a robust flagellum, the<br />
antennal flagellum extends to the posterior <strong>of</strong> pereonite<br />
4 (or anterior <strong>of</strong> 5) and the dorsum vaulted. <strong>The</strong> specimen<br />
is clearly an undescribed species, but is in poor<br />
condition, and at least one undamaged specimen is<br />
needed before it can be adequately characterised.<br />
2 0<br />
uNcErtAIN stAtus Or rEcOrds<br />
Aega cyclops Haswell, 88<br />
Aega cyclops Haswell, 88 : 92; 882: 285.– Hale, 925: 80,<br />
fig. 26; 1926: 233, fig. 20; 1937: 18; 1940: 298.– Bruce,<br />
983: 769, fig. 7O.– Springthorpe & Lowry, 994:<br />
43.–Bruce, Lew Ton & Poore, 2002: 6 .<br />
Aega (Rhamphion) cyclops.– Brusca, 983: .<br />
materiaL examined: “Possible syntype”; labelled as type<br />
by A.R. McCulloch, 905. ♂ ( 0.3 mm), Port Jackson,<br />
NSW, Australia (AM G5326).<br />
[Roger Springthorpe (Australian Museum, Sydney) states,<br />
in correspondence, that this specimen cannot be identified<br />
as the holotype with any degree <strong>of</strong> certainty. Haswell<br />
( 88 ) did not designate types and did not mention how<br />
many specimens he used in the original description.<br />
Hale ( 925) redescribed this specimen and assumed<br />
that it was the ‘type’ probably because McCulloch, in<br />
his somewhat cavalier fashion, had labelled it as such.<br />
Haswell’s original description lacks detail, for example<br />
it not mentioning the damaged pleotelson that Hale<br />
described. This specimen is similar in length to the<br />
original and may be a syntype. <strong>The</strong> origin <strong>of</strong> much <strong>of</strong> the<br />
Old Collection labelled as type material is confounded. It<br />
cannot be shown that this material was used by Haswell<br />
in his original descriptions. Material from Port Jackson,<br />
and some Queensland localities, for example, may have<br />
been collected after the date <strong>of</strong> publication by others such<br />
as Whitelegge, McCulloch, and Hedley.]<br />
remarks: Despite being relatively widely recorded<br />
(Hale 925, 926, 937, 940) <strong>of</strong>f the coast <strong>of</strong> southeastern<br />
Australia the species remains poorly known<br />
and characterised. <strong>The</strong> holotype, a dry and dissected<br />
specimen, is held at the Australian Museum. All pereopods<br />
have lost their distal articles, and the uropods<br />
and pleotelson had been damaged and regrown prior<br />
to collection. <strong>The</strong>re are two slides with an entire pereopod<br />
, pereopod 7, maxilliped and appendix masculina<br />
(detached from the pleopod). From this material there<br />
are several characteristics which distinguish Aega<br />
cyclops, these being the medially fused penial processes<br />
(a defining character state for Epulaega), the huge<br />
eyes with a very small posterior clear field, the short<br />
and smoothly curved dactylus on pereopod and the<br />
small, ovate frontal lamina. No other species has this<br />
combination <strong>of</strong> characters.<br />
Hale ( 926) reported the species from ‘South-east<br />
<strong>of</strong> Sydney, in “<strong>New</strong> <strong>Zealand</strong> area,” 75 faths.’ However,<br />
that species seems more likely to be Aegiochus coroo<br />
(Bruce, 1983) given the large posterior clear field on the<br />
head (Hale, 1926, figure 20), and that the record was<br />
provisionally included in the synonymy for that species<br />
by Bruce ( 983). Hale ( 940) later reported several<br />
more localities in southeastern Australia for A. cyclops,<br />
but those records are here regarded as unconfirmed. A
evision <strong>of</strong> the species based on good-quality material<br />
that can be identified by comparison to the holotype<br />
and existing slide material will clearly establish the<br />
identity <strong>of</strong> the species, but until that time I regard Aega<br />
cyclops as species inquirenda.<br />
Aega cyclops is not regarded as occurring in <strong>New</strong><br />
<strong>Zealand</strong>.<br />
Aega novizealandiae Dana, 853, nomen dubium<br />
Æga novi-zealandiae Dana, 1853: 767, pl. 51, fig 2a–c.<br />
Æga novae Zelandiae.– Lütken, 859: 77.<br />
Æga neo-zelanica.– Thomson & Chilton, 886: 53.<br />
Æga novae-zealandiae.– Hutton, 904: 262.<br />
Aega novae-zealandiae.– Miers, 876b: 08; Thomson, 9 3:<br />
246.<br />
Aega novae-zeelandiae.– Nierstrasz, 93 : 82.<br />
Aega novi-zealandiae.– Tattersall, 1921: 213, pl. 4, figs 11–14;<br />
Hurley, 96 : 268.<br />
Aega neozelandia.– Brusca, 983: .<br />
type LoCaLity: Bay <strong>of</strong> Islands, <strong>New</strong> <strong>Zealand</strong>.<br />
remarks: <strong>The</strong> identity <strong>of</strong> Aega novizealandiae Dana, 853<br />
(the spelling <strong>of</strong> the species name has been impressively<br />
inconsistent over time) is impossible to establish. It is<br />
uncertain that the family and generic placement are<br />
correct, this being noted by Dana himself. <strong>The</strong>re are<br />
numerous Southern Ocean species <strong>of</strong> Aegiochus that<br />
are both characterised and distinguished by details<br />
<strong>of</strong> the frontal lamina, shape <strong>of</strong> the anterior pereopods<br />
together with details <strong>of</strong> the number and orientation<br />
<strong>of</strong> robust setae, and also details <strong>of</strong> the number and<br />
arrangement <strong>of</strong> robust setae on the pleotelson and<br />
uropodal rami. None <strong>of</strong> this information is available<br />
for Aega novizealandiae. Dana’s personal notes (unpublished;<br />
copy from the Smithsonian Institution) record<br />
the loss <strong>of</strong> the collections on the bar <strong>of</strong> the Colombia<br />
River, a notoriously dangerous crossing, as well as<br />
further accidental losses to the material in shipment<br />
and unpacking, so in the absence <strong>of</strong> types there is<br />
no chance to obtain data by which this species might<br />
be characterised. <strong>The</strong>re are at least two small-sized<br />
and small-eyed species <strong>of</strong> Aegiochus in northern <strong>New</strong><br />
<strong>Zealand</strong> waters, either <strong>of</strong> which may be Aega novizealandiae.<br />
Equally A. novizealandiae could belong to the<br />
Cirolanidae, Corallanidae or Tridentellidae.<br />
It seems that the identity <strong>of</strong> Aega novizealandiae can<br />
never be resolved and it is here placed in the category<br />
nomen dubium and henceforth excluded from the<br />
<strong>New</strong> <strong>Zealand</strong> fauna.<br />
2<br />
Rocinela orientalis Schioedte & Meinert, 879<br />
Rocinela orientalis Schioedte & Meinert, 879b: 395, pl 3,<br />
figs 1–2.– Gerstaecker, 1882: 260.– Miers, 1884: 304.–<br />
Stebbing, 905: 25, pl. VI.C; 9 0: 0 .– Richardson,<br />
1910: 17.– Chilton, 1911: 567.– Hale, 1925: 183, fig.<br />
27.– Nierstrasz, 1931:184, figs 75–77.– Monod, 1933:<br />
94.– Barnard, 936: 368.– Hurley, 96 : 268.– Pillai.<br />
1967: 279, fig. 7e–f, pl. II, 5.– Kensley, 2001: 227.<br />
type materiaL: Syntypes (MCZ 3 3 ) held at Museum<br />
<strong>of</strong> Comparative Zoology, Harvard, USA; type locality<br />
Calcutta, West Bengal, India.<br />
remarks: Recorded from a beached specimen on Raoul<br />
Island, Kermadec Islands by Chilton ( 9 ). <strong>The</strong> record<br />
would require confirmation given the recorded tropical<br />
distribution <strong>of</strong> South Africa and East Africa to India,<br />
the Philippines and eastern Australia. <strong>The</strong> identity <strong>of</strong><br />
many earlier records also need confirmation, particularly<br />
as differences have been commented on, and it is<br />
now apparent that the genus is diverse (nine species)<br />
in the <strong>New</strong> <strong>Zealand</strong> region. <strong>The</strong> original description<br />
<strong>of</strong> R. orientalis gives minimal data and figures only the<br />
dorsal view and frons, and there is not sufficient detail<br />
to identify the species. Several records in the synonymy<br />
(e.g. Gerstaecker 882; Hurley 96 ; Kensley 200 )<br />
are merely repeat citations and are not based on new<br />
material or records. At present it seems likely that the<br />
<strong>New</strong> <strong>Zealand</strong> record is a misidentification.<br />
Rocinela orientalis is regarded as not occurring in<br />
<strong>New</strong> <strong>Zealand</strong>.
Aega Leach, 8 5<br />
Aega acuminata Hansen, 1897; East Pacific <strong>of</strong>f Galapagos<br />
Is. and <strong>of</strong>f Costa Rica; 768– 353 m (Brusca<br />
983).<br />
Aega acuticauda Richardson, 9 0; Philippines; 245 m;<br />
(possibly the juvenile <strong>of</strong> A. antennata).<br />
Aega angustata Whitelegge, 90 ; <strong>New</strong> South Wales,<br />
Australia; 99–2 9 m (see p. 232).<br />
Aega antennata Richardson, 9 0; Philippines, between<br />
Gillolo and Kayoa Islands; 485 m; in need <strong>of</strong> redescription.<br />
Aega approximata Richardson, 9 0; Philippines, Palawan<br />
Passage; 689 m; in need <strong>of</strong> redescription.<br />
Aega bicarinata Leach, 8 8; type locality stated as<br />
‘Localité: inconnue’; northeastern Atlantic, 22 m<br />
(Holthuis 956; Kussakin 979); Holthuis ( 956)<br />
placed this name under synonymy with Aega<br />
rosacea (Risso, 8 6), but this synonymy was not<br />
accepted by Kussakin ( 979); A. rosacea is here<br />
considered as nomen dubium.<br />
Aega chelipous Barnard, 960; Madagascar, from Carcharinus;<br />
60 m; in need <strong>of</strong> redescription.<br />
Aega concinna Hale, 940; Australia, Tasmania; depths<br />
not recorded; in need <strong>of</strong> redescription. Both pereopods<br />
2 and 3 have large club shaped robust seta<br />
opposing the dactylus, and pereopod inferior<br />
margin is convex and swollen.<br />
Aega crenulata Lütken, 859; North Atlantic, Greenland,<br />
Iceland and Norway; 85–950 m; from the<br />
Greenland shark Somniosus microcephalus (Bloch<br />
and Schneider, 80 ) (Richardson 905a, Kussakin<br />
979).<br />
Aega d<strong>of</strong>leini Thielemann, 9 0; Japan, Sagami Bay;<br />
depth not recorded; in need <strong>of</strong> redescription.<br />
Whereabouts <strong>of</strong> the type material not known; apparently<br />
lost or destroyed in World War Two.<br />
Aega ecarinata Richardson, 898. Off Little Bahama<br />
Banks and <strong>of</strong>f entrance to San Juan, 65–6 7 m;<br />
Puerto Rico (Kensley & Schotte 989); in need <strong>of</strong><br />
redescription.<br />
Aega falcata Kensley & Chan, 200 ; Taiwan; 500 m.<br />
Aega hirsuta Schioedte & Meinert, 879b; Nice, Mediterranean<br />
France (no other data). Identity uncertain;<br />
possibly a junior synonym <strong>of</strong> A. tridens.<br />
Aega lecontii (Dana, 854); California, Monterey; a<br />
poorly known species <strong>of</strong> uncertain identity;<br />
whereabouts <strong>of</strong> the type material is not known<br />
to me; in need <strong>of</strong> redescription.<br />
spEcIEs INcludEd IN thE AEgIdAE<br />
This list is additive to those species treated in principal taxonomic account; entries are alphabetical by genus and<br />
species.<br />
2 2<br />
Aega magnifica (Dana, 853); both coasts <strong>of</strong> southern<br />
South America to Straits <strong>of</strong> Magellan; 0– 8 m<br />
(Bruce 2004a).<br />
Aega maxima Hansen, 897; near Galapagos Is.; 2350 m<br />
(Brusca 983).<br />
Aega megalops Norman & Stebbing, 904 (in Norman<br />
904); Portugal; 82 m; also South Africa (Barnard<br />
9 4; Stebbing 922); in need <strong>of</strong> redescription .<br />
Aega microphthalma Dana, 854; California, Monterey<br />
(Richardson 905a); the whereabouts <strong>of</strong> the type<br />
material is unknown; species inquirenda according<br />
to Brusca ( 983); in need <strong>of</strong> redescription.<br />
Aega nanhaiensis yu, 2007; South China Sea; 85–<br />
5 m.<br />
Aega platyantennata Nunomura, 993; Japan, Sea <strong>of</strong><br />
Japan, <strong>of</strong>f Himi city, Toyama Prefecture; from<br />
Lophius setigerus (currently Lophiomus setigerus<br />
(Vahl, 797), Lophiidae); depth not recorded; in<br />
need <strong>of</strong> redescription.<br />
Aega psora (Linnaeus, 758); type species; widely recorded<br />
in the North Atlantic, including Gulf <strong>of</strong><br />
Mexico, and US coasts, Greenland, Iceland, south<br />
to Irish and British waters; 48– 280 m (Kussakin<br />
979, Richardson 905a); hosts include both bony<br />
and cartilaginous fishes. <strong>The</strong> record <strong>of</strong> this species<br />
from the Red Sea (Bakhrebah 2006) is a misidentification,<br />
the figures unambiguously showing a<br />
species <strong>of</strong> Aegiochus <strong>of</strong> unknown identity.<br />
Aega punctulata Miers, 88 ; Straits <strong>of</strong> Magellan, Falkland<br />
Islands, South Atlantic; depth range not<br />
recorded but presumed shallow (see p. 234).<br />
Aega rosacea (Risso, 8 6); Mediterranean, France; the<br />
brief description was accompanied by a single<br />
simple figure <strong>of</strong> the dorsal view. It is possible<br />
that the species is a the same as A. bicarinata, but<br />
on the basis <strong>of</strong> the existing description and the<br />
lack <strong>of</strong> types, the species can be considered as<br />
nomen dubium.<br />
Aega serripes H. Milne Edwards, 840; Australia, <strong>New</strong><br />
South Wales, Victoria and South Australia; 8 m<br />
(Hale 925, Bruce 983).<br />
Aega sheni yu & Bruce, 2006; China and eastern Australia<br />
(Coral Sea); 300–435 m.<br />
Aega stroemii Lütken, 859; Lütken gave the distribution<br />
as Norway, Færoe Islands and England, and<br />
included A. monophthalma and A. bicarinata as<br />
junior synonyms. <strong>The</strong> name was used earlier as
a nomen nudum in a footnote, by Krøyer ( 843—<br />
an attributed date as actual date <strong>of</strong> publication<br />
is not clear; previously cited as 837). In need <strong>of</strong><br />
revision.<br />
Aega tridens Leach, 8 5; poorly known; northeastern<br />
Atlantic, Britain, Færoes, Norway; to 200 m (Sars<br />
897, Kussakin 979).<br />
Aega truncata Richardson, 9 0; <strong>of</strong>f North Mindanao,<br />
Philippines; 308–4 4 m, in siliceous sponges.<br />
This species is similar to Aega urotoma Barnard,<br />
9 4, but Richardson’s description provides little<br />
information on appendages. <strong>The</strong> propodus <strong>of</strong><br />
A. truncata appears to lack the large distal robust<br />
setae opposite the base <strong>of</strong> dactylus <strong>of</strong> pereopods<br />
2 and 3 (it is not mentioned), and has more robust<br />
setae on the inferior margin <strong>of</strong> the merus (her<br />
description <strong>of</strong> the anterior pereopod reverses the<br />
carpus and merus). In need <strong>of</strong> redescription.<br />
Aega webbii (Guérin-Méneville, 836); Portugal; 00–<br />
300 m; this species remains poorly known, and<br />
subsequent records are all <strong>of</strong> uncertain identity or<br />
incorrect; the species is not recognizable from the<br />
original description; the type specimen is held at<br />
the Academy <strong>of</strong> Natural Sciences, Philadelphia.<br />
Trilles and Justine’s (2004) record from <strong>New</strong> Caledonia<br />
is a misidentification <strong>of</strong> Aega urotoma (see<br />
p. 55); the species is in need <strong>of</strong> redescription.<br />
Aegapheles gen. nov.<br />
Aegapheles antillensis (Schioedte & Meinert, 879b),<br />
comb. nov.; Caribbean and Gulf <strong>of</strong> Mexico;<br />
70–240 m (Bruce 2004a).<br />
Aegapheles banda (Bruce, 2004), comb. nov.; Banda Sea,<br />
Indonesia and <strong>of</strong>f northern Western Australia;<br />
290–4 6 m.<br />
Aegapheles deshaysiana (H. Milne Edwards, 840), comb.<br />
nov.; eastern North Atlantic and Mediterranean,<br />
from the Azores at approximately 38°N south<br />
to about 5°N; 00– 46 m, one record at 05 m<br />
(Bruce 2004a).<br />
Aegapheles excisa (Richardson, 9 0), comb. nov.; Philippines<br />
and Japan; 6 and 340 m (Bruce 2004a).<br />
Aegapheles japonica (Bruce, 2004), comb. nov.; Japan;<br />
20 m.<br />
Aegapheles kixalles (Bruce, 2004), comb. nov.; <strong>New</strong> Caledonia;<br />
540–545 m.<br />
Aegapheles kwazulu (Bruce, 2004), comb. nov.; <strong>of</strong>f<br />
Natal, South Africa, western Indian Ocean;<br />
237 m.<br />
Aegapheles musorstom (Bruce, 2004), comb. nov.; <strong>New</strong><br />
Caledonia; 475–6 5 m.<br />
Aegapheles trulla (Bruce, 2004), comb. nov.; Coral Sea <strong>of</strong>f<br />
Australia and <strong>of</strong>f Ontong Java, Solomon Islands;<br />
650–752 m.<br />
2 3<br />
Aegapheles warna (Bruce, 2004), comb. nov.; southeastern<br />
Australia from Tasmania to Victoria;<br />
33–5 8 m.<br />
Aegiochus Bovallius, 885<br />
Aegiochus arctica (Lütken, 859), comb. nov.; North<br />
Atlantic, Greenland and Iceland; 720– 500 m;<br />
from the Greenland shark Somniosus microcephalus<br />
(Bloch and Schneider, 80 ) (Richardson 905a,<br />
Kussakin 979); in need <strong>of</strong> redescription.<br />
Aegiochus australis (Whitelegge, 90 ), comb. nov.; Australia,<br />
<strong>New</strong> South Wales; 89– 02 m (Hale 925);<br />
the shape <strong>of</strong> the anterior margin <strong>of</strong> the frontal<br />
lamina and the small eyes are characteristic; the<br />
antennule peduncle is moderately compressed<br />
but not expanded; the species is in need <strong>of</strong> redescription.<br />
Aegiochus crozetensis (Kussakin & Vasina, 982), comb.<br />
nov.; southern Indian Ocean, vicinity <strong>of</strong> Crozet<br />
Is.; 280 m (this account, see p. 237).<br />
Aegiochus cyclops (Haswell, 882), comb. nov.; Australia,<br />
<strong>of</strong>f <strong>New</strong> South Wales; species inquirenda (see<br />
Aega cyclops p. 2 0).<br />
Aegiochus dentata (Schioedte & Meinert, 879b), comb.<br />
nov.; Cuba (no other data); later figures (e.g. Kensley<br />
& Schotte 989) are taken from the original<br />
description; in need <strong>of</strong> redescription.<br />
Aegiochus dollfusi (Monod, 933), comb. nov.; Red Sea,<br />
Egyptian Gulf <strong>of</strong> Suez; depth not recorded.<br />
Aegiochus francoisae (Wetzer, 990), comb. nov.; Galapagos,<br />
<strong>of</strong>f Fernandina Is.; taken from cloaca <strong>of</strong> an<br />
ascidian; 3 6 m.<br />
Aegiochus gracilipes (Hansen, 895), comb. nov.; North<br />
Atlantic, <strong>of</strong>f northwestern Scotland and Gulf <strong>of</strong><br />
Mexico; 335–2787 m (Richardson 905a, Kussakin<br />
1979); confirmation that the eastern and<br />
western Atlantic population are the same species<br />
is needed.<br />
Aegiochus incisa (Schioedte & Meinert, 879b), comb.<br />
nov.; Mediterranean (no other data); in need <strong>of</strong><br />
redescription.<br />
Aegiochus leptonica (Bruce, 988), comb. nov.; western<br />
Atlantic, <strong>of</strong>f Dry Tortugas, Florida; 048 m.<br />
Aegiochus longicornis (Hansen, 897), comb. nov.; East<br />
Pacific, <strong>of</strong>f Galapagos Is.; 842 m; (Brusca 983); in<br />
need <strong>of</strong> redescription.<br />
Aegiochus perulis (Menzies & George, 972), comb.<br />
nov.; <strong>of</strong>f Peru (8° 3’S); 927– 997 m; in need <strong>of</strong><br />
redescription.<br />
Aegiochus plebeia (Hansen, 897), comb. nov.; East Pacific,<br />
near the Galapagos Islands (see p.238).<br />
Aegiochus quadratisinus (Richardson, 903), comb. nov.;<br />
Hawaii; 207– 459 m (Bruce 983); Richardson<br />
( 904b) published the description twice.
Aegiochus sarsae (Brandt & Andres, 2008), comb. nov.;<br />
North Atlantic, from the northern Mid-Atlantic<br />
Ridge; 346 m.<br />
Aegiochus spongiophila (Semper, 867), comb. nov.;<br />
Philippines, several localities; at least to 52 m<br />
(Miers 878; Richardson 9 0); in need <strong>of</strong> redescription.<br />
Aegiochus symmetrica (Richardson, 905b), comb.<br />
nov.; Alaska; 75– 96 m; Kussakin ( 979) given<br />
the maximum depth as 050 m. In need <strong>of</strong> redescription.<br />
Aegiochus synopthalma (Richardson, 909), comb. nov.;<br />
Japan; 354.6 m (Bruce, 983).<br />
Aegiochus tenuipes (Schioedte & Meinert, 879b); Cuba<br />
(no other data); see comments for A. dentata; in<br />
need <strong>of</strong> redescription.<br />
Aegiochus tumida (Nunomura, 988), comb. nov.; <strong>of</strong>f<br />
Philippines; 400 m; from ‘Venus flower basket’<br />
sponge, Euplectella sp. Probable junior synonym<br />
<strong>of</strong> Aegiochus spongiophila, a species also known<br />
from Euplectella.<br />
Aegiochus uschakovi (Kussakin, 967), comb. nov.; Chile,<br />
Drake Passage; 95– 05 m (this account, p. 24 ).<br />
Aegiochus ventrosa (M. Sars, 859), comb. nov.; type<br />
species. North Atlantic, including Greenland,<br />
Norway and Britain; (Norman 904; Richardson<br />
905a; Kussakin 997); 220–570 m.<br />
Aegiochus weberi (Nierstrasz, 93 ), comb. nov.; Indonesia,<br />
Celebes Sea; 450 m; described from three<br />
male specimens, one <strong>of</strong> which has a large pair <strong>of</strong><br />
distally spatulate cephalic processes on the head,<br />
a character unique within the family. <strong>The</strong> figures<br />
suggest the possibility that the material consists<br />
<strong>of</strong> two species.<br />
Alitropus Milne Edwards, 840<br />
Alitropus typus H. Milne Edwards, 840; Indo-<br />
Malaysian region to eastern Australia; freshwater,<br />
attacks fishes (Bruce 1983). <strong>The</strong>re are several other<br />
proposed names (see Ingle & Fernando 964),<br />
and also A. foveolatus Schioedte & Meinert, 879b,<br />
which are here all regarded as junior synonyms.<br />
Epulaega gen. nov.<br />
Epulaega lethrina (Bruce, 983), comb. nov.; Queensland,<br />
Great Barrier Reef, Coral Sea and Papua<br />
<strong>New</strong> Guinea; shallow to 0 m, likely deeper;<br />
occurs in nasal passage <strong>of</strong> some Serranidae and<br />
Lethrinidae, occasionally other fishes; also from<br />
sponges.<br />
Epulaega monilis (Barnard, 9 4), comb. nov.; South Africa,<br />
Table Bay to East London, 90–33 m (Kensley<br />
2 4<br />
978) (material examined under E. fracta, present<br />
study); in need <strong>of</strong> redescription.<br />
Epulaega nodosa (Schioedte & Meinert, 879b), comb.<br />
nov.; southeastern Australia; depths mostly<br />
not previously reported, to at least 40 m; (Bruce<br />
983).<br />
Rocinela Leach, 8 8<br />
Rocinela affinis Richardson, 904a; Japan to Australia<br />
(Bruce et al. 2002); 306 m; in need <strong>of</strong> redescription.<br />
Rocinela americana Schioedte & Meinert, 879b; western<br />
North Atlantic, Maine, USA; 55–287 m (Kussakin<br />
979).<br />
Rocinela angustata Richardson, 1904a; northern Pacific<br />
from Japan and Alaska, along the North American<br />
coast southwards to Baja California; 50–466 m<br />
(Brusca & France 992). Nunomura (2006) was<br />
apparently unaware <strong>of</strong> the redescription and<br />
designation <strong>of</strong> a lectotype by Brusca and France<br />
( 992), and made comments on numerous differences<br />
between his material, the original description<br />
and the redescription by Kussakin ( 979). <strong>The</strong><br />
identity <strong>of</strong> Nunomura’s material cannot be readily<br />
elucidated from the description and drawings.<br />
Feeding and prey preferences in captivity have<br />
been reported by Wing and Moles ( 995).<br />
Rocinela australis Schioedte & Meinert, 879b; Straits <strong>of</strong><br />
Magellan; shallow; in need <strong>of</strong> redescription.<br />
Rocinela belliceps (Stimpson, 1864); East Pacific from<br />
Alaska to Mexico; 59–284 m (Brusca & France<br />
992). Implicated in attacks on aquacultured<br />
salmon (Novotny & Mahnken 97 ).<br />
Rocinela cornuta Richardson, 898; Alaska; 43 m<br />
(Richardson 905a); 00– 200 m according to<br />
Kussakin ( 979).<br />
Rocinela cubensis Richardson, 898; Caribbean,<br />
Cuba; 262 m (Richardson 905a); in need <strong>of</strong> re-<br />
description.<br />
Rocinela danmoniensis Leach, 8 8; England 25– 250 m<br />
(Kussakin 979); in need <strong>of</strong> redescription.<br />
Rocinela dumerilii (Lucas, 849); Atlantic: South Africa<br />
(Kensley 978), Cuba and Mediterranean<br />
(Richardson 905a); 60–500 m; in need <strong>of</strong> re-<br />
description.<br />
Rocinela granulosa Barnard, 9 4; Western Indian<br />
Ocean, <strong>of</strong>f Natal, South Africa; 80–200 m (Kensley<br />
978); in need <strong>of</strong> redescription.<br />
Rocinela hawaiiensis Richardson, 903; eastern Indo-<br />
Pacific (Hawai’i) to East Pacific (Baja California,<br />
Mexico) (Brusca & France 992); 766– 200 m;<br />
Richardson ( 904b) republished the description<br />
as new.
Rocinela insularis Schioedte & Meinert, 879b; Caribbean,<br />
Mississippi to Florida; 425–499 m (Richardson<br />
905a).<br />
Rocinela japonica Richardson, 898; Japan; 20–64 m (Kussakin<br />
979); in need <strong>of</strong> redescription.<br />
Rocinela juvenalis Menzies and George, 972; East Pacific,<br />
<strong>of</strong>f Peru; 4506 m; in need <strong>of</strong> redescription.<br />
Rocinela kapala Bruce, 988; <strong>of</strong>f Cape Moreton, Queensland<br />
to Sydney, <strong>New</strong> South Wales, Australia;<br />
450–765 m.<br />
Rocinela laticauda Hansen, 897; there are only two<br />
positive records for the East Pacific <strong>of</strong>f Acapulco,<br />
Mexico and <strong>of</strong>f California; 20–960 m (Brusca &<br />
France 992).<br />
Rocinela lukini Vasina, 993; Sea <strong>of</strong> Okhotsk; 753–<br />
480 m; recorded prey Raja binoculata and<br />
Hippoglossus sp.<br />
Rocinela maculata Schioedte & Meinert, 879b; Greenland<br />
and Vladivostok, Russia; 0–22 m (Kussakin<br />
979; Richardson 905a); in need <strong>of</strong> redescription.<br />
Rocinela media Nierstrasz, 93 ; Buton Strait, southern<br />
Sulawesi, Indonesia; 75–94 m; known from a single<br />
male specimen, in need <strong>of</strong> redescription.<br />
Rocinela modesta Hansen, 1897; East Pacific, <strong>of</strong>f Bay<br />
<strong>of</strong> Panama, Panama; 848 m (Brusca & France<br />
992).<br />
Rocinela murilloi Brusca & Iverson, 1992; East Pacific<br />
from California to Chile; 786– 866 m (Brusca &<br />
France 992).<br />
Rocinela niponia Richardson, 909; Japan; 08 m; in need<br />
<strong>of</strong> redescription.<br />
Rocinela oculata Harger, 883; <strong>of</strong>f Georgia, USA; 46 m<br />
(Richardson 905a).<br />
Rocinela orientalis Schioedte & Meinert, 879b; widely<br />
recorded from the tropical Indo-Pacific (see Hale,<br />
925, present account, p. 2 ); 22–500 m; in need<br />
<strong>of</strong> redescription.<br />
Rocinela patriciae Brasil-Lima, 986; <strong>of</strong>f Rio Grande<br />
do Sul, Brazil; depth and precise position not<br />
stated.<br />
Rocinela propodialis Richardson, 905b; Admiralty Inlet,<br />
Port Townsend, Alaska; 27–48 m; in need <strong>of</strong><br />
redescription.<br />
2 5<br />
Rocinela richardsonae Nierstrasz, 93 ; Banda Sea,<br />
Indonesia; 560 m; known from a single female<br />
specimen, in need <strong>of</strong> redescription.<br />
Rocinela signata Schioedte & Meinert, 1879b; East Pacific<br />
from California to Ecuador; western Atlantic and<br />
Caribbean from Florida to Brazil; intertidal to 68<br />
m (Brusca & France 992).<br />
Rocinela sila Hale, 925; Port Adelaide, South Australia<br />
(depths not stated).<br />
Rocinela tridens Hatch, 947; Washington State, USA;<br />
in need <strong>of</strong> redescription.<br />
Rocinela tropica Brasil-Lima, 986; Vitória, Espírito Santo,<br />
Brazil, 8°38’S, 39°34’W; depth not stated.<br />
Rocinela tuberculosa Richardson, 898; Gulf <strong>of</strong> California;<br />
5–33 m (Brusca & France 992).<br />
Rocinela wetzeri Brusca & France, 1992; East Pacific<br />
at Galapagos Islands and <strong>of</strong>f Costa Rica; 57–<br />
2000 m.<br />
Syscenus Harger, 880<br />
Syscenus atlanticus Kononenko, 988; western North<br />
Atlantic; 730–200 m (Kensley 2004; Brandt &<br />
Andres 2008).<br />
Syscenus infelix Harger, 880; North Atlantic and Mediterranean;<br />
70–207 m (Kensley 2004).<br />
Syscenus intermedius Richardson, 9 0; to the south <strong>of</strong><br />
Hong Kong, South China Sea, 20°37’N, 5°43’E;<br />
380–380 m, and Banda Sea, Indonesia (see p.<br />
24 ).<br />
Syscenus karu Bruce, 2005; southwestern Pacific, <strong>of</strong>f<br />
Vanuatu, with prominently bulbous and faceted<br />
eyes; 480–455 m.<br />
Syscenus peruanus Menzies & George, 972; tropical<br />
East Pacific, <strong>of</strong>f Peru, at approximately 7°S; the<br />
species is known from a single juvenile specimen,<br />
with faceted eyes present; atypically deep for the<br />
genus at 4526–4609 m.<br />
Xenuraega Tattersall, 909<br />
Xenuraega ptilocera Tattersall, 909; north-<br />
eastern Atlantic; 3 0– 250 m (Tattersall 9 ,<br />
Bruce 993a).
Oss sEA ANd ANtArctIc IslANds (bAllENy IslANds) spEcIEs<br />
Aegiochus antarctica (Hodgson, 9 0), comb. nov.<br />
(Fig. 37)<br />
Æga australis Richardson, 906a: 87 (name pre-occupied,<br />
Aega australis Whitelegge, 90 ).<br />
Æga australis Richardson, 906b: 850.<br />
Æga australis Richardson, 1908: 4, figs 8–11.<br />
Æga antarctica Hodgson, 9 0: 7, pl. 2.– Richardson, 9 3:<br />
4.<br />
Aega antarctica.– Tattersall, 92 : 2 .– Monod, 926: 5.– Hale,<br />
937: 9; 952: 28.– Kussakin, 967: 224; 982: 74.– Amar<br />
& Roman, 974: 582.– Arnaud, 974: 647.– Jaramillo,<br />
1977: 60, fig. 1.– Schultz, 1978: 31, fig. 7.– Wägele,<br />
1990: 521.– Brandt, 1991: 221, figs 2–4.– Nunomura,<br />
2005: 68, fig. 4.<br />
Aega koltuni Kussakin, 1967: 228, figs 5, 6; 1982: 74.– Kensley,<br />
200 : 227 (new synonymy).<br />
Aega (Rhamphion) koltuni.– Brusca, 983: .<br />
materiaL examined: Paratype <strong>of</strong> Aega koltuni (c. 24 mm,<br />
head and pereonite missing), Elephant Island, South<br />
Shetlands, 6 ° 5’S, 57°48’E, 0 June 958, Ob’ stn 460,<br />
370–400 m, coll. Koltunin (LiN RAN 2 N46 4).<br />
Antarctic ross sea and balleny Islands: 2♀ (nonovig.<br />
8.5, 24 mm), 77°05’S, 64° 2’E, 24 January<br />
9 2, 256 m, Terra Nova stn 339 (NMNZ Cr. 093). ♀<br />
(non-ovig. 23 mm), RV Tangaroa stn K0803/29 (NIWA<br />
24004). cape Adare: ♀ (non-ovig. or imm. 2.5 mm),<br />
7 °43.88’S, 7 °45.00’E, 5 February 2004, 45 m, gravel,<br />
small stones, shell, coral (NIWA 23660); ♀ (non-ovig. 23<br />
mm), 7 °43.67’S, 7 °44. 2’E, 5 February 2004, 397–389<br />
m, coral, rubble, shell (NIWA 2366 ); ♀ (non-ovig. 22<br />
mm), 72°08.04’S, 7 °26.92’E, 26 February 2004, 466–438<br />
m, coral and rubble (NIWA 23662). 2♀ (non-ovig. 8.0,<br />
2.5 mm), manca (9.5 mm), southern rookery, Cape<br />
Bird, February 97 , 83 m, sponge and hydroid bottom,<br />
coll. GSK & JKL (AM P4397 ). balleny Islands: ♀<br />
(non-ovig. or imm. 5.0 mm), 67°25.07’S, 63°54.93’E,<br />
4 March 2004, 230–228 m, rubble (NIWA 23663); 2♀<br />
(non-ovig. or imm. 2.0, ovig., damaged [head missing]<br />
~20 mm), 65°24.76’S, 60°53.22’E, 7 March 2004,<br />
4– 5 m (NIWA 23664).<br />
NIwA ‘old’ ross sea collections (specimens not<br />
measured). , Cape Armitage, McMurdo Sound, 20<br />
April 1957, fish trap on seabed under bay ice, 122 m,<br />
stn Z 509 (NIWA 23665); 4, stn 298, Cape Evans,<br />
77°38.0’S, 66°20.0’E, 23 February 958, beam trawl, 24<br />
m, stn Z 5049 (NIWA 23666); , SU 24*, 27 December<br />
958, Loc. E, eel trap, stn Z 5092 (NIWA 23667); (very<br />
poor condition), 3 January 959, Trap A, on surface<br />
(NIWA 23668); 2, A449, 77°05’S, 77° 2’E, January<br />
959, s<strong>of</strong>t gritty mud, 362 m (NIWA 23669; two lots);<br />
2, A457, 77°35’S, 73° 8’E, 6 January 959, (NIWA<br />
23670); ~ 5 (2 lots, poor condition), A456, Pennell<br />
2 6<br />
Bank, 74°30’S, 79°40’E, 5 January 959, 238–30 m<br />
(NIWA 2367 ); , A464, 73°20’S, 73°00’E, 22 January<br />
959, 369–384 m, sand and pebbles (NIWA 23672); 2,<br />
A468, east <strong>of</strong> Beaufort Is, 76°59’S, 67°36’E, 26 January<br />
959, 0 m (NIWA 23673); 2, A47 , <strong>of</strong>f Cape Evans<br />
and Barne Glacier, 77°37’S, 66°20’E, 6 February 959,<br />
65– 69 m (NIWA 23674); 2, A533, Cape Barne, 77°35’S,<br />
66° 0’E, 6 February 960, #27, 97– 83 m (NIWA<br />
23675); , south <strong>of</strong> Cape Armitage, McMurdo Sound,<br />
77°5 .90’S, 66°43.23’E, 2 November 96 , Dearborn<br />
loc. 6 D, trap, NIWA stn Z 5098 (NIWA 23696). , Cape<br />
Armitage, 5 February 957, from seal stomach, ‘stn’<br />
379, Transantarctic (N.Z.) Expedition (NIWA 23676);<br />
, south <strong>of</strong> Hut Point, McMurdo Sound, 77°5 .23’S,<br />
66°39.02’E, 2 May 959, SU 26, Dearborn loc. M, 38<br />
m (D.S.T.), NIWA stn Z 5093 (NIWA 23677); , SU 7,<br />
4 January 960 (NIWA 23678); , SU 20, Cape Evans,<br />
22 January 960, Weddell seal stomach (NIWA 23679);<br />
, south <strong>of</strong> Hut Point, McMurdo Sound, 77°5 .23’S,<br />
66°39.02’E, 3 September 959, SU 8, Dearborn loc. M,<br />
38 m, NIWA stn Z 5094 (NIWA 23680); , <strong>of</strong>f Arrival<br />
Heights, McMurdo Sound, 77°50.0 ’S, 66°35.92’E,<br />
26 November 959, SU 27, Dearborn loc. S, 64.5 m,<br />
NIWA stn Z 5095 (NIWA 2368 ); , <strong>of</strong>f Hut Point, Mc-<br />
Murdo Sound, 77°5 .05’S, 66°37.50’E, 29 November<br />
959, SU 23, Dearborn loc. P, 57 m, NIWA stn Z 5096<br />
(NIWA 23682); , <strong>of</strong>f Arrival Heights, McMurdo Sound,<br />
77°49.92’S, 66°34.692’E, 6 December 959, SU 2 ,<br />
Dearborn loc. T, surface (NIWA 23683); , 3 June 96<br />
(NIWA 23684); , SN23, 29 June 96 , trap, ½ m net,<br />
Dearborn loc. 6 D, NIWA stn Z 5098 (NIWA 23685);<br />
, 3 August 96 , trap, Dearborn loc. 6 D, NIWA stn<br />
Z 5098 (NIWA 23686); , 28 September 96 , Dearborn<br />
loc. 6 D, trap, NIWA stn Z 5098 (NIWA 23687); 4,<br />
3 October 96 , Dearborn loc. 6 D, trap, NIWA stn<br />
Z 5098 (NIWA 23688); , southeast <strong>of</strong> Cape Armitage,<br />
McMurdo Sound, 77°5 .99’S, 66°43.23’E, 6 October<br />
96 , 278–290 m, Dearborn loc. 6 B, trap, NIWA stn<br />
Z 5099 (NIWA 23689); (2 lots), 26 October 96 ,<br />
Dearborn loc. 6 D, trap, NIWA stn Z 5098 (NIWA<br />
*<strong>The</strong> SU prefix indicates the Stanford University Antarctic<br />
invertebrate studies carried out in the Ross Sea in the<br />
period 1958–61. However, the ‘SU’ prefix and number as<br />
recorded on the specimen labels do not relate to anything<br />
in the published station list (Dearborn 967). Similarly the<br />
dated numbers <strong>of</strong> the format ‘6 D’ could not be related<br />
with any confidence to stations in the former <strong>New</strong> <strong>Zealand</strong><br />
Oceanographic Institute station list (Bullivant 967).<br />
<strong>The</strong> precise location <strong>of</strong> the material without coordinates<br />
remains unconfirmed.
Figure 137. Aegiochus antarctica (Hodgson, 9 0). All NIWA 2366 . A, head, dorsal view; B, frons; C, pleonites, lateral view;<br />
D, maxillule apex; E, bifid seta, distal margin <strong>of</strong> maxilliped palp article 5; F, maxilliped palp articles 4–5; G, pereopod 1;<br />
H, pereopod 2; I, pleopod ; J, uropod exopod, ventral view; K, uropod endopod, apex; L, uropod endopod, apex; M, uropod.<br />
2 7
23690); , 3 October 96 , Dearborn loc. 6 D, trap,<br />
NIWA stn Z 5098 (NIWA 2369 ); 2, Pennell Bank,<br />
74°20’S, 79°30’E, 3 February 960, A520#24, 20 –205<br />
m (NIWA 23692); , Cape Barne, 77°36.77–36.00’S,<br />
66°08.0– 2.0’E, A534#24, 380–366 m (NIWA 23693); ,<br />
probably <strong>of</strong>f Hut Point, McMurdo Sound, 77°50.95’S,<br />
66°37.72’E, 22 December, SU 22, Dearborn loc. Hole<br />
C, NIWA stn Z 5 00 (NIWA 23694); , <strong>of</strong>f northwestern<br />
shore <strong>of</strong> Cape Armitage, 77°5 .42’S, 66°38.73’E, 3<br />
December 959, SU 25, Dearborn loc. W, 53 m, NIWA<br />
stn Z 5 0 (NIWA 23695).<br />
suppLementary desCription: Body 2.3 times as long as<br />
greatest width, with lateral margins subparallel or<br />
ovate. Rostral point folded ventrally and posteriorly.<br />
Eyes small, combined widths less than 50% width <strong>of</strong><br />
head, separated by about 45% width <strong>of</strong> head; each eye<br />
made up <strong>of</strong> ~ 2 transverse rows <strong>of</strong> ommatidia, each<br />
row with ~8 ommatidia; eye colour black. Pereonite<br />
1 and coxae 2–3 each with posteroventral angle right-<br />
angled. Coxae 5–7 with entire oblique carina; posterior<br />
margins straight, posterolateral angle acute (less than<br />
45°). Pleon with pleonite visible in dorsal view; pleonite<br />
4 with posterolateral margins extending to but<br />
not beyond posterior margin <strong>of</strong> pleonite 5; pleonite<br />
5 with posterolateral angles free, not overlapped by<br />
lateral margins <strong>of</strong> pleonite 4. Dorsal surface with weak<br />
longitudinal ridge; pleotelson lateral margins straight,<br />
serrate, pleotelson posterior margin with distinct short<br />
median point, with 0 robust setae.<br />
Antennule peduncle articles and 2 slender, article<br />
2 without distal lobe; flagellum extending to posterior<br />
<strong>of</strong> pereonite . Antenna flagellum extending to middle<br />
<strong>of</strong> pereonite 3.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, rectangular,<br />
anterior margin with median point, forming median<br />
angle, posterior margin not abutting clypeus.<br />
Mandible molar process present, small distinct flat<br />
lobe; palp article 2 with 2 distolateral setae, palp article<br />
3 with 26 setae. Maxillule with 4 terminal RS (one large,<br />
3 slender). Maxilla mesial lobe with 3 RS; lateral lobe<br />
with 3 RS. Maxilliped endite with 0 apical setae; palp<br />
article 2 with 2 RS (slender); article 3 with 4 recurved<br />
RS (and 3 short slender); article 4 with 4 hooked RS;<br />
article 5 articulating with article 4, longer than wide,<br />
sub-rectangular, with 4 RS (and 1 bifid seta).<br />
Pereopod 1 ischium inferior margin with 0 RS, superior<br />
distal margin with RS; merus inferior margin<br />
with 4 RS (and long simple seta), set as two groups<br />
(set as 2 and 2), superior distal angle with RS (and 2<br />
short simple setae); RS; propodus .9 times as long as<br />
proximal width, 2 RS, propodal palm simple, without<br />
blade or process, dactylus smoothly curved, .2 as<br />
long as propodus. Pereopod 2 ischium inferior margin<br />
2 8<br />
with 0 RS, superior distal margin with RS; merus<br />
inferior margin with 4 RS, set as two groups (set as 2<br />
and 2), superior distal margin with 2 acute RS; carpus<br />
similar in size to that <strong>of</strong> pereopod , inferodistal angle<br />
with 2 RS. Pereopod 3 similar to pereopod 2; propodus<br />
without large club-shaped distal robust seta. Inferior<br />
margins with palmate setae; margin with 2 RS (in 2<br />
groups), superior distal angle with 4 RS, distal angle<br />
with 4 RS.<br />
Pleopod 1 exopod .7 times as long as wide, distally<br />
broadly rounded, lateral margin straight, mesial margin<br />
strongly convex, with PMS from distal half; endopod<br />
2.0 as long as wide, distally narrowly rounded, lateral<br />
margin straight, with PMS from distal half, mesial<br />
margin with PMS on distal one-third; peduncle 2.0 as<br />
wide as long, mesial margin with 8 coupling hooks.<br />
Pleopod 2 appendix masculina basally swollen, .3 times<br />
as long as endopod (Brandt 99 ), distally acute (with<br />
acute scales mid-length).<br />
Uropod peduncle ventrolateral margin with 3 RS,<br />
posterior lobe about three-quarters as long as endopod.<br />
Uropod rami extending beyond pleotelson, marginal<br />
setae in single tier, apices acute. Endopod apically subbifid,<br />
mesial process prominent, lateral margin proximally<br />
straight, without prominent excision, proximal<br />
lateral margin with 0 RS, distal lateral margin with 2<br />
RS (and in notch), mesial margin straight, with 4 RS.<br />
Exopod extending to end <strong>of</strong> endopod, 3.3 times as long<br />
as greatest width, apically deeply bifid or sub-bifid,<br />
medial process prominent; lateral margin weakly<br />
convex, with 8 RS; mesial margin straight, distally<br />
convex, with 4 RS.<br />
size: Recently collected material from the Ross Sea, all<br />
female, 2–24 mm; Brandt ( 99 ) recorded 0–20 mm<br />
for males, 7–30 mm for ovigerous females.<br />
Variation: Ross Sea specimens, collected 2004. Robust<br />
setae: pleotelson (n = 9) RS 8– 3 ranging from 4+4 to<br />
7+6 with 5+5 (33%), 5+6 (22%) and 6+6 (22%) most<br />
frequent. Uropod (n = 6) exopod mesial margin 3–5<br />
with 4 (50%) or 5 (44%) most frequent, lateral margin<br />
8– 0 with 8 (44%) and 9 (50%) most frequent; uropod<br />
endopod mesial margin varied from 5– 0 RS with 5<br />
(3 %) and 6 (25%) the most frequent, lateral margin<br />
variable, with 0+2, 0+3, +2 and +3; the most frequent<br />
counts were +2 (50%) and +3 (44%).<br />
remarks: Aegiochus antarctica is readily identified by<br />
the small eyes that are little larger than ‘cirolanid<br />
size’ in conjunction with a short pleotelson that has<br />
an ill-defined but distinct median longitudinal ridge.<br />
At the northern limits <strong>of</strong> its distribution the species<br />
may be sympatric with other congeners, but multiple<br />
characters <strong>of</strong> eye size, frontal lamina shape, presence <strong>of</strong>
flattened antennule peduncle articles, and pereopodal<br />
and uropodal details will separate those species from<br />
A. antarctica.<br />
Comparison <strong>of</strong> the type material <strong>of</strong> Aegiochus koltuni<br />
(Kussakin, 967) reveals no differences from Aegiochus<br />
antarctica. Kussakin ( 967) in describing A. koltuni made<br />
no reference to A. antarctica, comparing only with the<br />
rather more different Aega magnifica; A. koltuni is here<br />
placed in synonymy with Aegiochus antarctica.<br />
Nunomura (2005) recorded this species from 20°E<br />
<strong>of</strong>f Antarctica (western sector <strong>of</strong> the southern Indian<br />
Ocean), and commented on a number <strong>of</strong> differences<br />
from other, unspecified, descriptions. While the<br />
identification is probably correct, unfortunately the<br />
illustrations in Nunomura’s account are <strong>of</strong> insufficient<br />
accuracy and detail to allow for comparison with material<br />
at hand from the Ross Sea or with other figured<br />
descriptions (e.g. Hodgson 9 0 or Brandt 99 ).<br />
<strong>The</strong> original date <strong>of</strong> publication has been somewhat<br />
confused, in part by Richardson’s ( 9 3) reluctance to<br />
accept that she was not attributed authorship <strong>of</strong> the<br />
new name published by Hodgson ( 9 0), and by the<br />
repeated publication <strong>of</strong> the description as a new species.<br />
Irrespectively, the correct name, and authority, is<br />
Aegiochus antarctica (Hodgson, 9 0).<br />
Wägele ( 990) documented the growth and reproductive<br />
biology <strong>of</strong> this species in captivity.<br />
distribution: A widespread and apparently common<br />
species, with numerous records from the Weddell Sea,<br />
the Ross Sea and subantarctic islands <strong>of</strong> the Southern<br />
Ocean; the northerly records are at about 60° South; at<br />
depths (present material) <strong>of</strong> 38– 300 m, with all but one<br />
record less than 450 metres; Brandt ( 99 ) recorded a<br />
maximum depth <strong>of</strong> 7 0 m.<br />
Aegiochus glacialis (Tattersall, 92 ), comb. nov.<br />
(Fig. 38)<br />
Aega glacialis Tattersall, 1921: 211, pl. 4, figs 1–10.– Monod,<br />
926: 5– Hale, 937: 9; 952: 28.– Kussakin, 967: 225;<br />
1982: 74.– Brandt, 1991: 216, fig. 1, 4 (tel 2).<br />
Aega (Ramphion) glacialis.– Brusca, 983: .<br />
materiaL examined: All ross sea: ♀ (non-ovig. 36 mm),<br />
7 °44. –88’S, 7 °44.00–43. 5’E, 5 February 2004,<br />
429–454 m (NIWA 23697); ♀ (non-ovig. 37 mm), Hut<br />
Point, McMurdo Sound, 30 January 960, 300 m (NIWA<br />
23698). 2 ♂ (2 mm, ~ 8 mm head missing), McMurdo<br />
Sound, RS 4 (NIWA 23699).<br />
NIwA ‘old’ collections (specimens not measured):<br />
SU 9*, 0 January 960, Dearborn loc. x, 35 m,<br />
trap (NIWA 23700); , southeast <strong>of</strong> Cape Armitage,<br />
*See comments concerning station data under ‘material<br />
examined’ for A. antarctica, p. 23 .<br />
2 9<br />
McMurdo Sound, 77°5 .99’S, 66°43.23’E, 8 July 96 ,<br />
Dearborn loc. 6 B, 27 –290 m, trap (NIWA 2370 );<br />
2, southeast <strong>of</strong> Cape Armitage, McMurdo Sound,<br />
77°5 .99’S, 66°43.23’E, 24 July 96 , Dearborn loc. 6 B,<br />
27 –290 m, trap (NIWA 23702); , southeast <strong>of</strong> Cape<br />
Armitage, McMurdo Sound, 77°5 .99’S, 66°43.23’E,<br />
8 August 96 , 27 –290 m, Dearborn loc. 6 B, trap<br />
(NIWA 23703); , 25 August 96 , Dearborn loc. 6 B,<br />
trap (NIWA 23704); , 4 September 96 , Dearborn<br />
loc. 6 D, trap (NIWA 23705); , 6 October 96 , Dearborn<br />
loc. 6 B, trap (NIWA 23706); 2, 6 October 96 ,<br />
Dearborn loc. 6 B, trap (NIWA 23707).<br />
Also examined: ♂ (20 mm), Antarctica (Atlantic sector),<br />
75° 5’S, 26° 4’W, 29 January 909, 500 m, EPOS3,<br />
stn 229, GSN6, Petersen, Tendal & Schiøtte on RV<br />
Polarstern (ZMUC unreg).<br />
suppLementary desCription: Rostral point folded ventrally<br />
and posteriorly. Eyes large, not medially united,<br />
separated by about 9% width <strong>of</strong> head; each eye made<br />
up <strong>of</strong> ~23 transverse rows <strong>of</strong> ommatidia, each row with<br />
~ 2– 6 ommatidia; eye colour black. Pleon with pleonite<br />
4 with posterolateral margins extending clearly<br />
beyond posterior margin <strong>of</strong> pleonite 5; pleonite 5 with<br />
posterolateral angles free, not overlapped by lateral<br />
margins <strong>of</strong> pleonite 4. Pleotelson 0.9 times as long as<br />
anterior width, dorsal surface with weak longitudinal<br />
ridge; lateral margins weakly concave, smooth, posterior<br />
margin with distinct short median point, with<br />
0– 4 RS.<br />
Antennule peduncle articles and 2 slender,<br />
article 2 without distal lobe; flagellum extending<br />
to pereonite 2. Antenna flagellum extending to middle<br />
<strong>of</strong> pereonite 3.<br />
Frontal lamina posterior margin free, downwardly<br />
projecting, blade-like, wider than long, posterior<br />
margin concave (weakly), anterior margin rounded,<br />
posterior margin not abutting clypeus.<br />
Maxillule with 4 or 5 terminal RS (one large, 3 or 4<br />
slender). Maxilla lateral lobe with 3 RS. Maxilliped palp<br />
article 2 with 3 RS (slender); article 3 with 3 recurved<br />
RS (and 3 straight); article 4 with 4 hooked RS (and<br />
mesial short slender seta); article 5 articulating with<br />
article 4, longer than wide, sub-rectangular, with 6 RS<br />
(3 long, serrate, curved and 2 short).<br />
Pereopod 1 ischium inferior margin with 0 RS, superior<br />
distal margin with RS; merus inferior margin<br />
with 3 RS (and one simple seta), set as two groups (<strong>of</strong> 2<br />
and ), superior distal angle with 0 RS; carpus inferior<br />
margin with RS (and simple seta); propodus .9<br />
times as long as proximal width, RS, propodal palm<br />
with small distal lobe, dactylus smoothly curved, .2 as<br />
long as propodus. Pereopod 2 ischium inferior margin<br />
with 0 RS, superior distal margin with RS; merus<br />
inferior margin with 4 RS (distal RS large, proximal<br />
small), set as two groups, superior distal margin with
Figure 138. Aegiochus glacialis (Tattersall, 92 ). All NIWA 23697. A, head, dorsal view; B, frons; C, pleonite, lateral view;<br />
D, pereopod ; E, pereopod 2; F, maxillule apex; G, maxilliped palp articles 4–5; H, pleopod ; I, pleopod 2; J, uropod exopod,<br />
apex; K, uropod exopod, ventral view; L, uropod.<br />
220
0 acute RS (2 simple setae); carpus similar in size to<br />
that <strong>of</strong> pereopod , inferodistal angle with 2 RS (set<br />
on inferodistal lobe). Pereopod 3 similar to pereopod 2;<br />
propodus without large club-shaped distal RS.<br />
Penes low, mutually adjacent tubercles.<br />
Pleopod 1 exopod 2.6 times as long as wide, distally<br />
narrowly rounded, mesial margin weakly oblique,<br />
lateral margin straight, mesial margin strongly convex,<br />
with PMS from distal half; endopod 2.6 times as long<br />
as wide, distally narrowly rounded, lateral margin<br />
sinuate, with PMS from distal one-third, mesial margin<br />
with PMS on distal one-third; peduncle .8 times as<br />
wide as long, mesial margin with 9 coupling hooks.<br />
Pleopod 2 appendix masculina basally swollen, .04<br />
times as long as endopod, distally acute (with acute<br />
scales mid-length).<br />
Uropod peduncle ventrolateral margin with 3 RS,<br />
posterior lobe about one-third as long as endopod.<br />
Uropod rami not extending beyond pleotelson, marginal<br />
setae in single tier, apices acute. Endopod apically<br />
sub-bifid, mesial process prominent, lateral margin<br />
sinuate, without prominent excision, proximal lateral<br />
margin with RS, distal lateral margin with 3 RS,<br />
mesial margin straight, with 7 RS. Exopod extending<br />
to end <strong>of</strong> endopod, 3.2 times as long as greatest width,<br />
apically sub-bifid, mesial process prominent; lateral<br />
margin weakly convex, with 2 RS; mesial margin<br />
convex, with 6 RS.<br />
22<br />
remarks: A full description <strong>of</strong> Aegiochus glacialis has<br />
been given by Brandt ( 99 ). <strong>The</strong> species is readily<br />
separated from A. antarctica, the only other Antarctic<br />
species, by the far larger eyes, the inferior margin <strong>of</strong><br />
the carpus <strong>of</strong> pereopods 2 and 3 being lobate and the<br />
distal margin <strong>of</strong> the palm <strong>of</strong> pereopod being weakly<br />
lobed (compared to not lobed in A. antarctica); adult<br />
A. glacialis are also somewhat larger than A. antarctica<br />
with an adult size exceeding 30 mm (maximum here<br />
<strong>of</strong> 37 mm), while A. antarctica has not been recorded<br />
at a size greater than 24 mm.<br />
Aega australis Richardson, 906 is not a synonym<br />
<strong>of</strong> Aegiochus glacialis, contrary to the synonymy given<br />
by Brandt ( 99 ) for Aegiochus glacialis, but a junior<br />
homonym <strong>of</strong> Aega australis Whitelegge, 90 , a valid<br />
species, and a junior subjective synonym <strong>of</strong> Aegiochus<br />
antarctica [Brandt ( 99 ) did not give a synonymy for<br />
A. antarctica].<br />
distribution: Multiple records from the Ross Sea and<br />
the Weddell Sea; the most northerly record is at approximately<br />
72° South (Brandt 99 —though not all<br />
station data are provided for Brandt’s material); depth<br />
range 5–700 m (Brandt 99 ).
This monograph was supported by Foundation for<br />
Research, Science and Technology (FRST) programme<br />
Biodiversity <strong>of</strong> <strong>New</strong> <strong>Zealand</strong> Aquatic Environments, contract<br />
FRST COx02 9 (200 –2005), and subsequently<br />
FRST C0 x0502 (July 2005–2007).<br />
Many <strong>of</strong> the specimens referred to in this monograph<br />
were obtained via the R.V Tangaroa, operating<br />
for diverse FRST and Ministry <strong>of</strong> Fisheries contracts<br />
under program and/or cruise management <strong>of</strong> Dr<br />
Steve O’Shea, Don McKnight, Dr Malcolm Clark, Dr<br />
Ashley Rowden, Dr Scott Nodder, Dr Helen Neil, Dr<br />
Anne-Nina Lörz, including the following programs in<br />
their various iterations: Ocean Ecosystems (C0 x0223,<br />
C0 x0027), Earth–Ocean Change (C0 x0203), Seamounts:<br />
their importance for fisheries and marine ecosystems<br />
(C0 x0224) and Biodiversity survey <strong>of</strong> the western Ross<br />
Sea (ZBD200303). Funding for the joint <strong>New</strong> <strong>Zealand</strong>–Netherlands<br />
research initiative (TAN0 6) was<br />
also provided ALW-NWO (subsidy 835.20.008 to Dr<br />
G.C.A. Duineveld, “Frontal Organic matter Governing<br />
ecosystem Structure FROGS”).<br />
I gratefully acknowledge being allowed long-term<br />
loans <strong>of</strong> material from: National Museum <strong>of</strong> <strong>New</strong> <strong>Zealand</strong><br />
Te Papa Tongarewa (Rick Webber), the Australian<br />
Museum (Penny Berents), Museum Victoria (Gary<br />
Poore), Queensland Museum (Peter Davie), South<br />
Australian Museum (Wolfgang Zeidler), and Western<br />
Australian Museum (Diana Jones).<br />
I thank Drs Alain Crosnier and Danielle Defaye,<br />
and the Muséum national d’Histoire naturelle, Paris<br />
for arranging and supporting my study visits in 2005,<br />
and 2007 that contributed new species and numerous<br />
distributional records; for assistance with collections<br />
and equipment, I thank, too, Dr Danielle Defaye, Dr<br />
Nguen Ngoc-Ho, Régis Cleva, Gabrielle Gadaleta (all<br />
AcKNOwlEdgEmENts<br />
222<br />
Muséum national d’Histoire naturelle); and, finally,<br />
thanks to Dr Bertrand Richer de Forges (Institut de<br />
Recherche pour la Développement, Nouvelle Calédonie)<br />
for bringing additional material to my notice and<br />
for managing the field acquisition <strong>of</strong> these excellent<br />
collections and providing recently collected samples<br />
at short notice.<br />
I thank all others who kindly lent material referred<br />
to or otherwise provided assistance in the preparation<br />
and completion <strong>of</strong> this monograph: Paul Callomom<br />
(Academy <strong>of</strong> Natural Sciences, Philadelphia); Dr Alain<br />
Crosnier, Dr Danielle Defaye, Régis Cleva and Gabriel<br />
Gadaleta (all MNHN, Paris); Todd Landers (Auckland<br />
Institute and Museum); Ms Miranda Lowe (NHM<br />
London); Dr Noburu Nunomura (Toyama Science<br />
Museum); Marilyn Schotte, Cheryl Bright and the late<br />
Brian Kensley (USNM, Smithsonian Institution, Washington,<br />
DC); Dr Tin-yam Chan (Institute <strong>of</strong> <strong>Marine</strong><br />
Biology, National Taiwan Ocean University); Dr Regina<br />
Wetzer (LACM, Los Angeles); Dr Jørgen Olesen,<br />
who sourced rare and obscure literature (Zoological<br />
Museum, Natural History Museum <strong>of</strong> Denmark, University<br />
<strong>of</strong> Copenhagen); thanks to Dr A.L. Vereshchaka<br />
(Institute <strong>of</strong> Oceanology, Moscow) who hand-carried<br />
types <strong>of</strong> Kussakin’s specimens from Moscow to <strong>New</strong><br />
<strong>Zealand</strong> (and back). Dr Richard Taylor (Leigh <strong>Marine</strong><br />
Laboratory, University <strong>of</strong> Auckland) and Mr Steve<br />
Lowe (fisherman, Leigh, Auckland) are thanked for<br />
supplying specimens.<br />
Sylvie Bruce (Brisbane) inked and lettered all the<br />
original illustrations, over several years, with patience<br />
and care, for which I thank her. Ms Denise Seabright<br />
(MTQ, Townsville) pro<strong>of</strong> read the text and assisted<br />
with final figure scanning and preparation.
AMAR, R.; ROMAN, M.-L. 974: Invertébrés marins des<br />
xIIème et xVème expéditions Antarctiques Françaises<br />
en Terre Adélie. 4. Tanaidacés et isopodes. Tethys 5:<br />
56 –600.<br />
ARNAUD, P.M. 974: Contributions à la bionomie marine<br />
benthique des régions Antarctique et subantarctique.<br />
Tethys 6: 467–653.<br />
BAKHREBAH, A.O. 2006: Description <strong>of</strong> the <strong>Isopoda</strong> Aega<br />
psora (Linnaeus, 1758) infesting the Red Sea parrotfish<br />
“Scarus ferrugineus” in Jeddah, Saudi Arabia. Egyptian<br />
Journal <strong>of</strong> Aquatic Research 32(1): 450–456.<br />
BARNARD, K.H. 9 4: Contributions to the crustacean fauna<br />
<strong>of</strong> South Africa. 3. Additions to the marine <strong>Isopoda</strong> with<br />
notes on some previously incompletely known species.<br />
Annals <strong>of</strong> the South African Museum 10(11): 325–358a,<br />
359–442.<br />
BARNARD, K.H. 9 6: Contributions to the crustacean fauna<br />
<strong>of</strong> South Africa. 5.— <strong>The</strong> Amphipoda. Annals <strong>of</strong> the South<br />
African Museum 15(3): 05–302, pls. 26–28.<br />
BARNARD, K.H. 936: Isopods collected by R.I.M.S. “Investigator”.<br />
Records <strong>of</strong> the Indian Museum (Calcutta) 38(2):<br />
47– 9 .<br />
BARNARD, K.H. 940: Contribution to the crustacean fauna<br />
<strong>of</strong> South Africa. xII. Further additions to the Tanaidacea,<br />
<strong>Isopoda</strong>, and Amphipoda, together with keys for the<br />
identification <strong>of</strong> hitherto recorded marine and freshwater<br />
species. Annals <strong>of</strong> the South African Museum 32: 38 –543.<br />
BARNARD, K.H. 960: <strong>Isopoda</strong> parasitic on Madagascar<br />
fish. Institute Scientifique de Madagascar, Office de la<br />
Recherche Scientifique et Technique Outre-mers. Mémoires,<br />
série F, 3: 93–95.<br />
BATE, C.S. 878: Journal <strong>of</strong> the Royal Institute <strong>of</strong> Cornwall 5(19):<br />
65. [Cited in Neave, S.A. (Ed.) 939: Nomenclature Zoologicus:<br />
a list <strong>of</strong> the names <strong>of</strong> the genera and subgenera in zoology<br />
from the tenth edition <strong>of</strong> Linnaeus 1758 to the end <strong>of</strong> 2004. Vol.<br />
4, p. 79. Zoological Society <strong>of</strong> London, London.]<br />
BATE, C.S.; WESTWOOD, J.O. 86 – 868: A history <strong>of</strong> the<br />
British sessile-eyed <strong>Crustacea</strong>. John Van Voorst, London.<br />
536 p.<br />
BIGGS, B. 990: English–Maori Maori–English Dictionary.<br />
Auckland University Press, Auckland. 53 p.<br />
BIRSTEIN, ya.A. 973: Deep water isopods (<strong>Crustacea</strong>, <strong>Isopoda</strong>)<br />
<strong>of</strong> the North-Western part <strong>of</strong> the Pacific Ocean.<br />
Academy <strong>of</strong> Sciences <strong>of</strong> the U.S.S.R. 92: 69–224.<br />
BLEEKER, P. 857: Recherches sur les Crustacés de l’Inde<br />
Archipélagique. II. Sur les isopodes Cymothoadiens de<br />
l’Archipel Indien. Actes de la Société Indo-Neerlandaise,<br />
Batavia 2: 20–40.<br />
BOUVIER, E.L. 9 : Arthropodes marins. Annales de l’Institut<br />
Océanographique 3(3): 38–39.<br />
BOVALLIUS, C. 885: A new isopod from the Swedish Arctic<br />
Expedition <strong>of</strong> 883. Bihang til Kongeliga Svenska Vetenskapsakademiens<br />
Handlingar 10(9): – 2, pls , 2.<br />
rEFErENcEs<br />
223<br />
BOVALLIUS, C. 886: <strong>New</strong> or imperfectly known <strong>Isopoda</strong><br />
Part II. Bihang til Kongeliga Svenska Vetenskapsakademiens<br />
Handlingar 11(17): – 8, pls , 2.<br />
Branch, M.L.; Griffiths, C.L.; KensLey, B.; sieG, J.<br />
1991: <strong>The</strong> benthic <strong>Crustacea</strong> <strong>of</strong> subantarctic Marion and<br />
Prince edward Islands: Illustrated keys to the species<br />
and results <strong>of</strong> the 1982–1989 University <strong>of</strong> Cape Town<br />
surveys. South African Journal <strong>of</strong> Antarctic Research, 21(1),<br />
3–44.<br />
BRANDT, A. 99 : Redescription <strong>of</strong> the Antarctic fish<br />
parasites Aega glacialis Tattersall, 92 and Aega antarctica<br />
Hodgson, 9 0 (<strong>Crustacea</strong>: <strong>Isopoda</strong>: <strong>Aegidae</strong>). Senckenbergiana<br />
Maritima 21: 2 5–232.<br />
BRANDT, A.; ANDRES, H.-G. 2008: Description <strong>of</strong> Aega<br />
sarsae sp. nov. and redescription <strong>of</strong> Syscenus atlanticus<br />
Kononenko, 988 (<strong>Crustacea</strong>, <strong>Isopoda</strong>, <strong>Aegidae</strong>) from the<br />
Mid-Atlantic Ridge. <strong>Marine</strong> Biology Research 4: 6 –75.<br />
BRANDT, A.; CRAME, J.A.; POLZ, H.; THOMSON, M.R.A.<br />
999: Late Jurassic Tethyan ancestry <strong>of</strong> recent southern<br />
high-latitude marine isopods (<strong>Crustacea</strong>, Malacostraca).<br />
Paleontology 42: 663–675.<br />
BRANDT, A.; POORE, G.C.B. 2003: Higher classification <strong>of</strong><br />
the flabelliferan and related <strong>Isopoda</strong> based on a reappraisal<br />
<strong>of</strong> relationships. Invertebrate Systematics 17: 893–923.<br />
BRASIL-LIMA, I.M. 986: O gênero Rocinela Leach no<br />
litoral brasileiro, com descrição de duas espécies novas<br />
(<strong>Crustacea</strong>, <strong>Isopoda</strong>). Atas da Sociedade de Biologia do Rio<br />
de Janeiro 26: – 4.<br />
BROCCHI, G. 877: Liste des crustacés isopodes provenant<br />
de l’île Saint-Paul et déposés dan les galeries du Muséum<br />
d’histoire naturelle par MM. Vélain et de Lisle. Bulletin de<br />
la Société philomathématique de Paris (6), 11: 97– 0 .<br />
BROWN, R.W. 956: Composition <strong>of</strong> Scientific Words. Smithsonian<br />
Institution Press, Washington, DC. 863 p.<br />
BRÖKELAND, W.; WÄGELE, J.W.; BRUCE, N.L. 200 : Paravireia<br />
holdichi n. sp., an enigmatic isopod crustacean from<br />
the Canary Islands with affinities to species from <strong>New</strong><br />
<strong>Zealand</strong>. Organisms, Diversity and Evolution 1(2): 83–98.<br />
BRUCE, N.L. 98 : Cirolanidae (<strong>Crustacea</strong>: <strong>Isopoda</strong>) <strong>of</strong><br />
Australia: Diagnoses <strong>of</strong> Cirolana Leach, Metacirolana<br />
Nierstrasz, Neocirolana Hale, Anopsilana Paulian & Debouteville,<br />
and three new genera – Natatolana, Politolana<br />
and Cartetolana. Australian Journal <strong>of</strong> <strong>Marine</strong> and Freshwater<br />
Research 32: 945–966.<br />
BRUCE, N.L. 983: <strong>Aegidae</strong> (<strong>Isopoda</strong>: <strong>Crustacea</strong>) from Australia<br />
with descriptions <strong>of</strong> three new species. Journal <strong>of</strong><br />
Natural History 17: 757–788.<br />
BRUCE, N.L. 984: A new family for the isopod crustacean<br />
genus Tridentella Richardson, 905, with description <strong>of</strong><br />
a new species from Fiji. Zoological Journal <strong>of</strong> the Linnean<br />
Society 80: 447–455.<br />
BRUCE, N.L. 988: Aega leptonica, a new species <strong>of</strong><br />
aegid isopod crustacean from the tropical western At
lantic, with notes on Rocinela oculata Harger and Rocinela<br />
kapala, new species. Proceedings <strong>of</strong> the Biological Society <strong>of</strong><br />
Washington 101: 95– 0 .<br />
BRUCE, N.L. 993a: Redescription <strong>of</strong> the overlooked crustacean<br />
isopod genus Xenuraega (<strong>Aegidae</strong>, Flabellifera).<br />
Journal <strong>of</strong> the <strong>Marine</strong> Biological Association <strong>of</strong> the United<br />
Kingdom 73: 6 7–625.<br />
BRUCe, n.L. 1993b: Two new genera <strong>of</strong> marine isopod<br />
crustaceans (Flabellifera: sphaeromatidae) from southern<br />
Australia, with a reappraisal <strong>of</strong> the sphaeromatidae.<br />
Invertebrate Taxonomy 7: 151–171.<br />
BRUCE, N.L. 996: Aega komai, a new species <strong>of</strong> marine isopod<br />
crustacean (<strong>Aegidae</strong>: Flabellifera) from Japan. <strong>Crustacea</strong>n<br />
Research 25: 29– 36.<br />
BRUCE, N.L. 997a: A new species <strong>of</strong> Syscenus Harger, 880<br />
(<strong>Crustacea</strong>: <strong>Isopoda</strong>: <strong>Aegidae</strong>) from eastern Australia<br />
with a revised diagnosis to the genus. Records <strong>of</strong> the Australian<br />
Museum 49: 3– 20.<br />
BRUCE, N.L. 997b: A new genus <strong>of</strong> marine isopod (<strong>Crustacea</strong>:<br />
Flabellifera: Sphaeromatidae) from Australia and<br />
the Indo-Pacific region. Memoirs <strong>of</strong> the Museum <strong>of</strong> Victoria<br />
56(1): 45–234.<br />
BRUCE, N.L. 200 : <strong>Marine</strong> isopod crustaceans in <strong>New</strong> <strong>Zealand</strong>.<br />
Water and Atmosphere 9(3): 2– 3.<br />
BRUCE, N.L. 2002: Parasites or predators? <strong>New</strong> <strong>Zealand</strong>’s<br />
aegid isopod crustaceans. Biodiversity Update 5: 8.<br />
BRUCE, N.L. 2003: Giant marine bloodsuckers. Water and<br />
Atmosphere 11(3): 4.<br />
BRUCE, N.L. 2004a: Reassessment <strong>of</strong> the isopod crustacean<br />
Aega deshaysiana (Milne Edwards, 840) (Cymothoida:<br />
<strong>Aegidae</strong>) — a worldwide complex <strong>of</strong> 2 species. Zoological<br />
Journal <strong>of</strong> the Linnean Society 142(2): 35–232.<br />
BRUCE, N.L. 2004b: <strong>New</strong> species <strong>of</strong> the Cirolana ‘parva-group’<br />
(<strong>Crustacea</strong>: <strong>Isopoda</strong>: Cirolanidae) from coastal habitats<br />
around <strong>New</strong> <strong>Zealand</strong>. Species Diversity 9(1): 47–66.<br />
BRUCE, N.L. 2005: Two new species <strong>of</strong> the mesopelagic<br />
isopod genus Syscenus Harger, 880 (<strong>Crustacea</strong>: <strong>Isopoda</strong>:<br />
<strong>Aegidae</strong>) from the southwestern Pacific. Zootaxa 1070:<br />
3 –42.<br />
BRUCE, N.L.; LEW TON, H.M.; POORE, G.C.B. 2002: <strong>Aegidae</strong><br />
White, 850. Pp. 59– 63 in: Poore, G.C.B. (Ed.)<br />
<strong>Crustacea</strong>: Malacostraca: Syncarida and Peracarida:<br />
<strong>Isopoda</strong>, Tanaidacea, Mictacea, <strong>The</strong>rmosbaenacea,<br />
Spelaeogriphacea. (Zoological Catalogue <strong>of</strong> Australia Vol.<br />
19.2A). CSIRO Publishing, Melbourne.<br />
BRUSCA, R.C. 980: Handbook to the common intertidal invertebrates<br />
<strong>of</strong> the Gulf <strong>of</strong> California. 2nd edn. University <strong>of</strong><br />
Arizona Press, Tucson, Arizona. –5 3 p.<br />
BRUSCA, R.C. 983: A monograph on the isopod family<br />
<strong>Aegidae</strong> in the tropical eastern Pacific. <strong>The</strong> genus Aega.<br />
Allan Hancock Monographs in <strong>Marine</strong> Biology 12: –39.<br />
BRUSCA, R.C.; FRANCE, S.C. 992: <strong>The</strong> genus Rocinela (<strong>Crustacea</strong>:<br />
<strong>Isopoda</strong>: <strong>Aegidae</strong>) in the tropical eastern Pacific.<br />
Zoological Journal <strong>of</strong> the Linnean Society 106: 23 –275.<br />
BRUSCA, R.C.; IVERSON, E.W. 985: A guide to the marine<br />
isopod <strong>Crustacea</strong> <strong>of</strong> Pacific Costa Rica. Revista de Biologia<br />
Tropical (Universidad de Costa Rica) 33(1): –77.<br />
224<br />
BRUSCA, R.C.; WILSON, G.D.F. 99 : A phylogenetic analysis<br />
<strong>of</strong> the <strong>Isopoda</strong> with some classificatory recommendations.<br />
Memoirs <strong>of</strong> the Queensland Museum 31: 43–204.<br />
BULLIVANT, J.S. 967: <strong>New</strong> <strong>Zealand</strong> Oceanographic Institute<br />
Ross Sea investigations, 958–60: general account<br />
and station list. <strong>New</strong> <strong>Zealand</strong> Oceanographic Institute<br />
Memoir 32: 9–29.<br />
CANZ. 997: <strong>New</strong> <strong>Zealand</strong> Region Bathymetry. NIWA<br />
Chart, Miscellaneous Series, No 73. Charting Around <strong>New</strong><br />
<strong>Zealand</strong> Group (Carter, L.; Cook, J. D.; Foster, G. A.;<br />
Garlick, R. D.; Litchfield, N. J.; Mitchell, J. S.; Wright, J.<br />
C.), Wellington.<br />
CHILTON, C. 9 : <strong>The</strong> <strong>Crustacea</strong> <strong>of</strong> the Kermadec Islands.<br />
Transactions and Proceedings <strong>of</strong> the <strong>New</strong> <strong>Zealand</strong> Institute<br />
43: 544–573.<br />
CHILTON, C. 925: A new genus <strong>of</strong> <strong>Isopoda</strong> (Family Sphaeromidae).<br />
Records <strong>of</strong> the Canterbury Museum 2: 32 –326.<br />
CHILTON, C. 926: Zoological results <strong>of</strong> a tour in the Far<br />
East. <strong>The</strong> Tanaidacea and <strong>Isopoda</strong> <strong>of</strong> Tale Sap. Records <strong>of</strong><br />
the Indian Museum 28: 73– 85.<br />
CLARK, P.F.; PRESSWELL, B. 200 : Adam White: <strong>The</strong> crustacean<br />
years. Raffles Bulletin <strong>of</strong> Zoology 49(1): 49– 66.<br />
DALLWITZ, M.J.; PAINE, T.A.; ZURCHER, E.J. 997: User’s<br />
guide to the DELTA system. A general system for processing<br />
taxonomic descriptions. 4.08. CSIRO Division <strong>of</strong> Entomology,<br />
Canberra. – 60 p.<br />
DANA, J.D. 1852: On the classification <strong>of</strong> the <strong>Crustacea</strong> Choristopoda<br />
or Tetradecapoda. American Journal <strong>of</strong> Sciences<br />
and Arts 14(41): 297–3 6.<br />
DANA, J.D. 853: <strong>Crustacea</strong>. Pp. 696–805 in: United States<br />
Exploring Expedition during the years 1838, 1839, 1840,<br />
1841, 1842, under the command <strong>of</strong> Charles Wilkes, U.S.N. C.<br />
Sherman, Philadelphia.<br />
DANA, J.D. 854: Catalogue and descriptions <strong>of</strong> <strong>Crustacea</strong><br />
collected in California by Dr. John L. Le Conte. Proceedings<br />
<strong>of</strong> <strong>The</strong> Academy <strong>of</strong> Natural Sciences <strong>of</strong> Philadelphia 7:<br />
75– 77.<br />
DEARBORN, J.H. 967: Stanford University invertebrate<br />
studies in the Ross Sea 958–6 : general account and<br />
station list. <strong>New</strong> <strong>Zealand</strong> Oceanographic Institute Memoir<br />
32: 3 –47.<br />
DELANEy, P.M. 989: Phylogeny and biogeography <strong>of</strong> the<br />
marine isopod family Corallanidae (<strong>Crustacea</strong>, <strong>Isopoda</strong>,<br />
Flabellifera). Contributions in Science, Natural Museum <strong>of</strong><br />
Los Angeles County 409: –75.<br />
DE LIMA, J.; CHELLAPPA, S.; THATCHER, V.E. 2005:<br />
Livoneca redmanni Leach (<strong>Isopoda</strong>, Cymothoidae) and<br />
Rocinela signata Schioedte & Meinert (<strong>Isopoda</strong>, <strong>Aegidae</strong>),<br />
ectoparasites <strong>of</strong> Scomberomorus brasilensis Collette, Russo<br />
& Zavala-Camin (Ostheichthyes, Scombridae) no Rio<br />
Grande do Norte, Brazil. Revista Brasileira de Zoologia<br />
22(4): 04– 08.<br />
DESMAREST, A.-G. 825: Considérations Générales sur la<br />
Classe des Crustacés, et description des espèces de ces<br />
animaux, qui vivent dans la mer, sur les côtes, ou dans<br />
les eaux douces de la France. F.G. Levrault, Libraire,<br />
Strasbourg, Paris. i–xix, –446, tables –5, pls –55.
DOLLFUS, A. 89 : Crustacés Isopodes. In: Mission Scientifique<br />
du Cap Horn. 1882–1883. pp F55–F76, pls 8a, 8b.<br />
Gauthier-Villars et fils, Paris.<br />
DREyER, H.; WÄGELE, J.-W. 200 : Parasites <strong>of</strong> crustaceans<br />
(<strong>Isopoda</strong>: Bopyridae) evolved from fish parasites: molecular<br />
and morphological evidence. Zoology 103: 57– 78.<br />
DREyER, H.; WÄGELE, J.W. 2002: <strong>The</strong> Scutocoxifera ta.<br />
nov. and the information content <strong>of</strong> nuclear ssu rDNA<br />
sequences for reconstruction <strong>of</strong> isopod phylogeny (Peracarida:<br />
<strong>Isopoda</strong>). Journal <strong>of</strong> <strong>Crustacea</strong>n Biology 22(2):<br />
2 7–234.<br />
ELLIS, J. 98 : Some type specimens <strong>of</strong> <strong>Isopoda</strong> (Flabellifera)<br />
in the British Museum (Natural History), and the isopods<br />
in the Linnaean Collection. Bulletin <strong>of</strong> the British Museum<br />
(Natural History) 40(4): 2 – 28.<br />
FELDMANN, R.M.; GOOLAERTS, S. 2005: Palaega rugosa,<br />
a new species <strong>of</strong> fossil isopod (<strong>Crustacea</strong>) from Maastrichtian<br />
rocks in Tunisia. Journal <strong>of</strong> Paleontology 79:<br />
03 – 035.<br />
FILHOL, H. 885: Mission de l’île Campbell. Recherches<br />
zoologiques, botaniques et géologiques faites a l’île<br />
Campbell et en Nouvelle-Zélande. Recueil de Mémoires,<br />
Rapports et Documents relatifs a l’observation du passage<br />
de Vénus sur le soleil du 9 Décembre, 874. Libraire<br />
des Comptes Rendus des Séances de l’Academie des Sciences,<br />
Paris 3(2): 82, 35 pls, Atlas 55.<br />
FROESE, R.; PAULy, D. 2007: FishBase: World Wide Web<br />
electronic publication. 2006. www.fishbase.org<br />
GARZON-FERREIRA, J. 990: An isopod, Rocinela signata<br />
(<strong>Crustacea</strong>: <strong>Isopoda</strong>: <strong>Aegidae</strong>), that attacks humans. Bulletin<br />
<strong>of</strong> <strong>Marine</strong> Science 46(3): 8 3–8 5.<br />
GERSTAECKER, A. 882: Sechste Ordnung: <strong>Isopoda</strong>-Asseln.<br />
Pp. 8–278 in: Bronn, H.G. (Ed.) Die Klassen und Ordnungen<br />
des Thier-Reiches: wissenschaftlich dargestellt in Wort und<br />
Bild. C.F. Winter, Leipzig.<br />
GOSSE, P.H. 855: A Manual <strong>of</strong> <strong>Marine</strong> Zoology for<br />
the British Isles. Part I. John van Voorst, London.<br />
203 p.<br />
GRUBE, A.E. 864: Die Insel Lussin und ihre Meeresfauna: nach<br />
einem Sechswöchentlichen Aufenthalte. F. Hirt, Breslau.<br />
–75 p.<br />
GUÉRIN-MÉNEVILLE, F.-É. 836: Classe 7, Crustacés.<br />
Magasin de Zoologie, Sixième Année, chapitre un [pages<br />
without folios].<br />
GURJANOVA, E. 933: Die marinen Isopoden der Arktis. Pp.<br />
392–472 in: Römer, F.; Schaudinn, F. (Eds). <strong>Fauna</strong> Arctica.<br />
G. Fischer Verlag, Jena.<br />
GURJANOVA, E. 936: Crustacées. Isopodes des mers orientales.<br />
Institut de Zoologie, l’Académie des Sciences de<br />
l’URSS, Faune de l’URSS, n. ser. no. 6: i–xii, –278.<br />
HALE, H.M. 925: Review <strong>of</strong> Australian isopods <strong>of</strong> the cymothoid<br />
group. Part I. Transactions <strong>of</strong> the Royal Society <strong>of</strong><br />
South Australia 49: 28– 85.<br />
HALE, H.M. 926: Review <strong>of</strong> Australian isopods <strong>of</strong> the cymothoid<br />
group. Part II. Transactions <strong>of</strong> the Royal Society <strong>of</strong><br />
South Australia 50: 20 –34, pls 26, 27.<br />
HALE, H.M. 937: <strong>Isopoda</strong> and Tanaidacea. Australasian<br />
Antarctic Expedition 9 – 4. Under the leadership <strong>of</strong><br />
225<br />
Sir Douglas Mawson, O.B.E., B.E,. D.Sc., F.R.S. Scientific<br />
Reports, Series C.— Zoology and Botany 2 (Part 2): –45.<br />
HALE, H.M. 940: Report on the cymothoid <strong>Isopoda</strong> obtained<br />
by the F.I.S. “Endeavour” on the coasts <strong>of</strong> Queensland,<br />
<strong>New</strong> South Wales, Victoria, Tasmania, and South Australia.<br />
Transactions <strong>of</strong> the Royal Society <strong>of</strong> South Australia<br />
64(2): 288–304.<br />
HALE, H.M. 952: <strong>Isopoda</strong>. Families Cymothoidae and Serolidae.<br />
Pp. 2 –36 in: B.A.N.Z. Antarctic Research Expedition<br />
1929–1931, under the command <strong>of</strong> Sir Douglas Mawson,<br />
Reports—Series B (Zoology and Botany). University <strong>of</strong><br />
Adelaide, Adelaide, South Australia.<br />
HANSEN, H.J. 890: Cirolanidae et familiae nonnulae propinquae<br />
Musei Hauniensis. Det Kongelige Danske Videnskabernes<br />
Selskab Skrifter, Naturvidenskabelige og Mathematisk<br />
6(3): 237–426.<br />
HANSEN, H.J. 895: Isopoden, Cumaceen und Stomatopoden<br />
der Plankton-Expedition. Ergebnisse der Plankton-<br />
Expedition der Humboldt-Stiftung 2: – 05.<br />
HANSEN, H.J. 897: Reports on the dredging operations <strong>of</strong>f<br />
the west coast <strong>of</strong> central America to the Galapagos, to<br />
the west coast <strong>of</strong> Mexico, and in the Gulf <strong>of</strong> California,<br />
in charge <strong>of</strong> Alexander Agassiz, carried on by the U.S.<br />
Fish Commission Steamer ‘Albatross’, during 89 , Lieut.<br />
Commander Z.L. Tanner, U.S.N. commanding. xxII.<br />
Bulletin <strong>of</strong> the Museum <strong>of</strong> Comparative Zoology at Harvard<br />
College 31(5): 93– 29, pls –7.<br />
HANSEN, H.J. 9 6: <strong>The</strong> order <strong>Isopoda</strong>. Pp. –262 in: <strong>Crustacea</strong><br />
Malacostraca. Danish Ingolf Expedition. Zoologisk<br />
Museum, Copenhagen.<br />
HARGER, O. 880: Report on the marine <strong>Isopoda</strong> <strong>of</strong> <strong>New</strong><br />
England and adjacent waters. Report <strong>of</strong> the U.S. Commission<br />
for Fisheries 6: 297–462.<br />
HARGER, O. 883: Reports on the results <strong>of</strong> dredging, under<br />
the supervision <strong>of</strong> Alexander Agassiz, on the east coast<br />
<strong>of</strong> the United States, during the summer <strong>of</strong> 880, by the<br />
U.S. Coast Survey Steamer “Blake,” Commander J.R. Bartlett,<br />
U.S.N., commanding. xxIII. Report on the <strong>Isopoda</strong>.<br />
Bulletin <strong>of</strong> the Museum <strong>of</strong> Comparative Zoology at Harvard<br />
College 11(4): 9 - 04, pls -4.<br />
HASWELL, W.A. 88 : On some new Australian marine<br />
<strong>Isopoda</strong>. Part I. Proceedings <strong>of</strong> the Linnean Society <strong>of</strong> <strong>New</strong><br />
South Wales 5(4): 470–48 , pls 6– 9.<br />
HASWELL, W.A. 882: Catalogue <strong>of</strong> the Australian stalk-<br />
and sessile-eyed <strong>Crustacea</strong>. Australian Museum, Sydney.<br />
–324 p.<br />
HATCH, M.H. 947: <strong>The</strong> Chelifera and <strong>Isopoda</strong> <strong>of</strong> Washington<br />
and adjacent waters. University <strong>of</strong> Washington<br />
Publications in Biology 10(5): 55–274.<br />
HELLER, C. 868: Crustaceen. Reise des Österreischen<br />
Fregatte Novara um die Erde 857, 858, 859 unter den<br />
befehlen der Commodore B. von Wüllerstorf-Urbair.<br />
Zoologischer <strong>The</strong>il 2 (3): –280.<br />
HEMMINGSEN, W.; maCKENZIE, K. 996: A checklist <strong>of</strong><br />
the protozoan and metazoan parasites reported from<br />
the Atlantic cod, Gadus morhua L. Bulletin <strong>of</strong> the European<br />
Association <strong>of</strong> Fish Pathologists 13(4): 34– 37.<br />
HICKS, G.R.F.; HUAKI, M.J.; WEBBER, W.R.; yALDWyN,<br />
J.C. 99 : Inventory <strong>of</strong> the cnidarian, pycnogonid and
crustacean type specimens in the National Museum <strong>of</strong><br />
<strong>New</strong> <strong>Zealand</strong>. National Museum <strong>of</strong> <strong>New</strong> <strong>Zealand</strong> Miscellaneous<br />
Series 22: –23.<br />
HO, J.S.; TONGUTHAI, K. 992: Flabelliferan isopods<br />
(<strong>Crustacea</strong>) parasitic on freshwater fishes <strong>of</strong> Thailand.<br />
Systematic Parasitology 21(3): 203–2 0.<br />
HODGSON, T.V. 9 0: <strong>Crustacea</strong> Ix. <strong>Isopoda</strong>. Pp. –77 in:<br />
Harmer, S.F. (Ed.) National Antarctic Expedition 1901–1904.<br />
Natural History. 0 pls. British Museum (Natural History),<br />
London.<br />
HOLTHUIS, L.B. 956: <strong>Isopoda</strong> en Tanaidacea. <strong>Fauna</strong> van<br />
Nederland, Leiden 161: –280.<br />
HOLTHUIS, L.B. 977: <strong>The</strong> dates <strong>of</strong> publication <strong>of</strong> C. Spence<br />
Bate and J.O. Westwood’s `A history <strong>of</strong> British sessileeyed<br />
<strong>Crustacea</strong>.’. <strong>Crustacea</strong>na 33(3): 3 3–3 6.<br />
HURLEy, D.E. 957: Some Amphipoda, <strong>Isopoda</strong> and Tanaidacea<br />
from Cook Strait. Zoology Publications from Victoria<br />
University <strong>of</strong> Wellington 21: –20.<br />
HURLEy, D.E. 96 : A checklist and key to the <strong>Crustacea</strong><br />
<strong>Isopoda</strong> <strong>of</strong> <strong>New</strong> <strong>Zealand</strong> and Subantarctic Islands.<br />
Transactions <strong>of</strong> the Royal Society <strong>of</strong> <strong>New</strong> <strong>Zealand</strong> (Zoology)<br />
1: 259–292.<br />
HURLEy, D.E. 990: Charles Chilton: the Phreatoicoidea and<br />
other interests <strong>of</strong> a phreatic pioneer from down under.<br />
Bijdragen tot de Dierkunde 60(3–4): 233–238.<br />
HURLEy, D.E.; JANSEN, K.P. 977: <strong>The</strong> marine fauna <strong>of</strong><br />
<strong>New</strong> <strong>Zealand</strong>: Family Sphaeromatidae (<strong>Crustacea</strong> <strong>Isopoda</strong>:<br />
Flabellifera). <strong>New</strong> <strong>Zealand</strong> Oceanographic Institute<br />
Memoir 63: –95.<br />
HUTTON, F.W. 904: Index Faunæ Novæ <strong>Zealand</strong>iæ. Publ. for<br />
Philosophical Institute <strong>of</strong> Canterbury, <strong>New</strong> <strong>Zealand</strong>, by<br />
Dulan & Co., London. vii, 372 pp.<br />
INGLE, R.W.; FERNANDO, C.H. 964: On some fresh and<br />
brackish water crustaceans from Ceylon. <strong>Crustacea</strong>na 6:<br />
02– 09.<br />
JARAMILLO, E. 977: Aega antarctica Hodgson y Plak-<br />
arthrium typicum Chilton, en bahía South Antarctica<br />
Chilena (<strong>Crustacea</strong>, <strong>Isopoda</strong>). Serie Cientifica Instituto<br />
Antártico Chileno 5: 59–64.<br />
JOHNSTON, G. 834: Illustrations in British Zoology. Annals<br />
and Magazine <strong>of</strong> Natural History (ser. 1). 7: 230–235.<br />
KEABLE, S.J. 2006: Taxonomic revision <strong>of</strong> Natatolana (<strong>Crustacea</strong>:<br />
<strong>Isopoda</strong>: Cirolanidae). Records <strong>of</strong> the Australian<br />
Museum 58(2): 33–244.<br />
KEABLE, S.J.; POORE, G.C.B.; WILSON, G.D.F. 2004: [keys<br />
to the] Australian <strong>Isopoda</strong>: Families. Version 2 October<br />
2002. Australian Museum. http://crustacea.net (accessed<br />
June 2006).<br />
KENSLEy, B. 976: <strong>Isopoda</strong>n and tanaidacean <strong>Crustacea</strong><br />
from the St Paul and Amsterdam Islands, Southern Indian<br />
Ocean. Annals <strong>of</strong> the South African Museum 69(11):<br />
26 –323.<br />
KENSLEy, B. 978: Guide to the <strong>Marine</strong> Isopods <strong>of</strong> Southern<br />
Africa. South African Museum and <strong>The</strong> Rustica Press,<br />
Wynberg, Cape Town. 73 p.<br />
226<br />
KENSLEy, B. 980: <strong>Marine</strong> isopods from Marion, Prince<br />
Edward, and Crozet Islands (<strong>Crustacea</strong>, <strong>Isopoda</strong>). Annals<br />
<strong>of</strong> the South African Museum 82(5): 55– 85.<br />
KENSLEy, B. 200 : Biogeography <strong>of</strong> the marine <strong>Isopoda</strong><br />
<strong>of</strong> the Indian Ocean, with a check-list <strong>of</strong> species and<br />
records. Pp. 205–264 in: Kensley, B.; Brusca, R.C. (Eds).<br />
Isopod Systematics and Evolution. <strong>Crustacea</strong>n Issues 13. A.A.<br />
Balkema, Rotterdam.<br />
KENSLEy, B. 2004: Redescription and distribution <strong>of</strong> two<br />
species <strong>of</strong> Syscenus (<strong>Crustacea</strong>, <strong>Isopoda</strong>, <strong>Aegidae</strong>) in the<br />
North Atlantic. Sarsia 89(3): 60– 74.<br />
KENSLEy, B.; CARTES, J.E. 2003: Records and distribution<br />
<strong>of</strong> Syscenus infelix in the deep Mediterranean (<strong>Crustacea</strong>:<br />
<strong>Isopoda</strong>: <strong>Aegidae</strong>). Journal <strong>of</strong> the <strong>Marine</strong> Biological Association<br />
<strong>of</strong> the United Kingdom 83(4): 775–777.<br />
KENSLEy, B.; CHAN, T.-y. 2001: Two species <strong>of</strong> deep-sea<br />
flabelliferan isopods from Taiwan (<strong>Crustacea</strong>: Peracarida:<br />
<strong>Aegidae</strong>, Anuropidae). Journal <strong>of</strong> Natural History 35:<br />
48 –496.<br />
KENSLEy, B.; SCHOTTE, M. 989: Guide to the <strong>Marine</strong> Isopod<br />
<strong>Crustacea</strong>ns <strong>of</strong> the Caribbean. Smithsonian Institution Press,<br />
Washington, DC and London. 308 p.<br />
KENSLEy, B.; SCHOTTE, M.; SCHILLING, S. 2007: World<br />
list <strong>of</strong> <strong>Marine</strong>, Freshwater and Terrestrial <strong>Crustacea</strong><br />
<strong>Isopoda</strong>. National Museum <strong>of</strong> Natural History Smithsonian<br />
Institution: Washington DC. http://invertebrates.<br />
si.edu/isopod/<br />
KLITGAARD, A.B. 995: <strong>The</strong> fauna associated with outer<br />
shelf and upper slope sponges (Porifera, Demospongiae)<br />
at the Faroe Islands, northeastern Atlantic. Sarsia<br />
80: –22.<br />
KONONENKO, A.F. 988: A new parasitic isopode Syscenus<br />
atlanticus n. sp. (<strong>Isopoda</strong>, <strong>Aegidae</strong>) from the Atlantic<br />
Ocean. Parazitologiya (Leningrad) 22: 266–269.<br />
KRØyER, H.N. 843– 845: Footnote. Pp. 40–4 in: Danmarks<br />
Fiske. Kjøbenhavn, S. Triers.<br />
KUSSAKIN, O.G. 967: <strong>Fauna</strong> <strong>of</strong> <strong>Isopoda</strong> and Tanaidacea<br />
in the coastal zones <strong>of</strong> the Antarctic and subantarctic<br />
water. Pp. 220–380 in: Andriyashev, A.P.; Ushakov, P.V.<br />
(Eds). Biological Reports <strong>of</strong> the Soviet Antarctic Expedition<br />
( 955– 958). (Issled <strong>Fauna</strong> Moreii). Akademii Nauk<br />
SSSR, Leningrad.<br />
KUSSAKIN, O.G. 979: <strong>Marine</strong> and brackishwater likefooted<br />
<strong>Crustacea</strong> (<strong>Isopoda</strong>) from the cold and temperate waters<br />
<strong>of</strong> the Northern Hemisphere. Suborder Flabellifera.<br />
Izdatel’stvo Nauka, Leningrad. Opredeliteli po Faune SSSR,<br />
Izdavaemye Zoologicheskim Institutom Akademii Nauk SSSR.<br />
472 p. [In Russian].<br />
KUSSAKIN, O.G. 982: Supplement to the isopod crustacean<br />
fauna from the shelf zones <strong>of</strong> the Antarctic (from the<br />
material <strong>of</strong> the Soviet Antarctic Expedition 965– 968).<br />
Pp. 73– 05 in: Kafanov, A.I. (Ed.) <strong>Fauna</strong> and distribution <strong>of</strong><br />
<strong>Crustacea</strong>ns from the Southern and Antarctic Waters. Academy<br />
<strong>of</strong> Sciences <strong>of</strong> the USSR (Far East Science Center),<br />
Vladivostok.<br />
KUSSAKIN, O.G.; VASINA, G.S. 980: Additions to the<br />
marine <strong>Isopoda</strong> and Gnathiidae <strong>of</strong> Kerguelen Islands<br />
(Southern Indian Ocean). Tethys 9: 355–369.
KUSSAKIN, O.G.; VASINA, G.S. 982: Addition to the fauna<br />
<strong>of</strong> benthic <strong>Isopoda</strong> and Gnathiida (<strong>Crustacea</strong>) <strong>of</strong> sub-<br />
antarctic waters <strong>of</strong> the Indian Ocean. . <strong>Isopoda</strong> (Flabellifera<br />
and Anthuridea). Tethys 10(3): 26 –273.<br />
LANCHESTER, W.F. 902: On the <strong>Crustacea</strong> collected during<br />
the “Skeat Expedition” to the Malay Peninsula. Part<br />
II. Anomura, Cirripedia, and <strong>Isopoda</strong>. Proceedings <strong>of</strong> the<br />
Zoological Society <strong>of</strong> London 2: 363–38 .<br />
LATREILLE, P.A. 829: Crustacés, Arachnides et partie des<br />
Insectes. Pp. 29– 44 in: Cuvier, G. (Ed.) La Règne Animal<br />
distribuès d’après son organisation, pour servir de base à<br />
l’histoire naturelle des animaux et d’introduction à l’anatomie<br />
comparée. Deterville, Paris.<br />
LEACH, W.E. 8 5: A tabular view <strong>of</strong> the external characters<br />
<strong>of</strong> four classes <strong>of</strong> animals which Linné arranged under Insecta<br />
with the distribution <strong>of</strong> the genera composing three<br />
<strong>of</strong> these classes into Orders, and description <strong>of</strong> several<br />
new genera and species. Transactions <strong>of</strong> the Linnean Society<br />
<strong>of</strong> London 11: 306–400.<br />
LEACH, W.E. 8 8: Cymothoadées. Pp. 338–354 in: Cuvier,<br />
F. (Ed.) Dictionnaire des Sciences Naturelles. Strasbourg et<br />
Levrault, Paris.<br />
LILJEBORG, O. 85 : Norges, Crustaceer. Ofversigt af Konglige<br />
Vetenskaps-Akademiens Forhandlingar 8(2): 55–95.<br />
LINCOLN, R.J. 985: <strong>The</strong> marine fauna <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>:<br />
deep-sea <strong>Isopoda</strong> Asellota, family Haploniscidae. <strong>New</strong><br />
<strong>Zealand</strong> Oceanographic Institute Memoir 94: –56.<br />
LINNAEUS, C. 758: Systema naturae per regna tria naturae,<br />
seundum classes, ordines, genera, species, cum characteribus,<br />
diferrentiis, synonymis locis. (Edn. 0). –824 p.<br />
LOCKINGTON, W.N. 877: Description <strong>of</strong> seventeen new<br />
species <strong>of</strong> <strong>Crustacea</strong>. Proceedings <strong>of</strong> the California Academy<br />
<strong>of</strong> Sciences. Proceedings <strong>of</strong> the California Academy <strong>of</strong><br />
Sciences 7: 44–46.<br />
LUCAS, H. 849: Histoire naturelle des animaux articulés.<br />
Premier partie. Crustacés, Arachnides, Myriapodes et<br />
Hexapodes. Exploration scientifique de l’Algerie pendant<br />
les années 1840, 1841, 1842, Sciences physiques. Zoologie 1:<br />
xxxv, 88 , 8 pls.<br />
LÜTKEN, C.F. 859: Nogle bemærkninger om de nordiske<br />
Æga-Arter samt om Æga-Slægtens rette Begrændsning.<br />
Videnskabelige Meddelelser fra den naturhistorisk Forening i<br />
Kjøbenhavn for Aaret 1858: 65.<br />
MAnIgheTTI, B. 2001: Ocean circulation: the planet’s great<br />
heat engine. Water & Atmosphere 9(4): 12–14<br />
VON MARTENS, E. 868: Ueber einige ostasiatische Süsswasserthiere.<br />
Archiv für Naturgeschichte 34: 09–279.<br />
MENZIES, R.J. 962: <strong>The</strong> zoogeography, ecology, and systematics<br />
<strong>of</strong> the Chilean marine isopods. Reports <strong>of</strong> the<br />
Lund University Chile Expedition 1948–49. C.E.K. Gleerup,<br />
Lund. 62 p.<br />
MENZIES, R.J.; GEORGE, R.y. 972: Isopod <strong>Crustacea</strong> <strong>of</strong> the<br />
Peru–Chile Trench. Anton Bruun Report. Scientific Results <strong>of</strong><br />
the Southeast Pacific Expedition, Texas A & M Press: College<br />
Station Texas 9: – 24.<br />
MENZIES, R.; GLyNN, P.W. 968: <strong>The</strong> common marine<br />
isopod <strong>Crustacea</strong> <strong>of</strong> Puerto Rico. Studies on the <strong>Fauna</strong> <strong>of</strong><br />
Curaçao and other Caribbean Islands 104: – 33.<br />
227<br />
MENZIES, R.J.; KRUCZyNSKI, W.L. 983: Isopod <strong>Crustacea</strong><br />
(exclusive <strong>of</strong> Epicaridea). Memoirs <strong>of</strong> the Hourglass Cruises<br />
6(1): – 26.<br />
MIERS, E.J. 875: Descriptions <strong>of</strong> three additional species <strong>of</strong><br />
<strong>Crustacea</strong> from Kerguelen’s Land and Crozet Island, with<br />
remarks upon the genus Paramoera. Annals and Magazine<br />
<strong>of</strong> Natural History (ser. 4) 4. 16: 5– 8.<br />
MIERS, E.J. 876a: Descriptions <strong>of</strong> some new species <strong>of</strong> <strong>Crustacea</strong>,<br />
chiefly from <strong>New</strong> <strong>Zealand</strong>. Annals and Magazine <strong>of</strong><br />
Natural History 17: 2 8–229.<br />
MIERS, E.J. 876b: Catalogue <strong>of</strong> the stalk and sessile-eyed<br />
<strong>Crustacea</strong> <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>. Colonial Museum and Geological<br />
Department <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>, National History Publication<br />
10: i–xii, – 33.<br />
MIERS, E.J. 878: On species <strong>of</strong> <strong>Crustacea</strong> living within<br />
the Venus’s flower basket Euplectella and in<br />
Meyerina claviformis. Journal <strong>of</strong> the Linnean Society<br />
(Zoology) 8: 506–5 2.<br />
MIERS, E.J. 879 : <strong>Crustacea</strong>. In: An account <strong>of</strong> the petrological,<br />
botanical, and zoological collections made in<br />
Kerguelen’s Land and Rodriguez during the Transit <strong>of</strong><br />
Venus Expeditions ... in the years 874-75. Philosophical<br />
Transactions <strong>of</strong> the Royal Society <strong>of</strong> London 168: 200-2 4,<br />
pl. .<br />
MIERS, E.J. 88 : <strong>Crustacea</strong>. Pp. 6 –80 in: Günther, Albert.<br />
Account <strong>of</strong> the zoological collections made during the<br />
survey <strong>of</strong> H.M.S. Alert in the Straits <strong>of</strong> Magellan and the<br />
coast <strong>of</strong> Patagonia. Proceedings <strong>of</strong> the Zoological Society <strong>of</strong><br />
London 1881. pl. VII.<br />
MIERS, E.J. 884: <strong>Crustacea</strong>. Pp. 78–33 in: Report <strong>of</strong> the<br />
Zoological Collections made in the Indo-Pacific Ocean<br />
during the voyage <strong>of</strong> HMS ‘Alert’, 88 – 882 : 78-33 .<br />
MILNE EDWARDS, H. 840: Histoire naturelle des Crustacés,<br />
comprenant l’anatomie, la physiologie et la classification<br />
de ces animaux. Librairie Encyclopédique de Roret. Vol. 3:<br />
i–ii, –638. Roret, Paris.<br />
MONOD, T. 926: Tanaidacés, Isopodes et Amphi-<br />
podes. Pp. –67 in: Résultats du Voyage du S.y. ‘Belgica’<br />
en 897– 899. Volume 4. J.-E. Buschmann, Anvers.<br />
MONOD, T. 933: Mission Robert Ph. Dollfus en Égypte.<br />
Tanaidacea et <strong>Isopoda</strong>. Mémoirs Institute Égypte 21:<br />
6 –264.<br />
MONOD, T. 934: Isopodes marins des campagnes du ‘Le Lanessan’.<br />
Notes de l’Institut Océanographique de l’Indochine,<br />
Saigon 23: –22, pls –45.<br />
MOREIRA, P.S.; SADOWSKy, V. 979: An annotated bibliography<br />
<strong>of</strong> parasitic <strong>Isopoda</strong> <strong>Crustacea</strong>: <strong>of</strong> Chondrichthyes.<br />
Boletim do Instituto Oceanográfico, Sao Paulo 27: 95– 52.<br />
NAIR, G.A.; NAIR, N.B. 983: Effect <strong>of</strong> infestation with the<br />
isopod, Alitropus typus M. Edwards (<strong>Crustacea</strong>: Flabellifera:<br />
<strong>Aegidae</strong>) on the haematological parameters <strong>of</strong> the<br />
host fish, Channa striatus (Bloch). Aquaculture 30(1–4):<br />
– 9.<br />
NIERSTRASZ, H.F.; SCHUURMANS STEKHOVEN JR, J.H.<br />
930: <strong>Isopoda</strong> genuina. Die Tierwelt der Nord- und Ostsee<br />
X.e2: 57– 33.<br />
NIERSTRASZ, H.F. 93 : Die isopoden der Siboga-<br />
expedition. Pp. 6–227 in: Siboga-Expeditie. E.J. Brill,<br />
Leiden.
NORDENSTAM, A. 930: Tanaidacea and marine <strong>Isopoda</strong><br />
from Juan Fernandez. Pp. 525–552 in: Skottsberg, C. (Ed.)<br />
<strong>The</strong> natural history <strong>of</strong> Juan Fernandez and Easter Island. Vol.<br />
3. Uppsala, Almqvist & Wiksells, 20 pl.<br />
NORMAN, A.M. 904: British <strong>Isopoda</strong> <strong>of</strong> the families <strong>Aegidae</strong>,<br />
Cirolanidae, Idoteidae, and Arcturidae. Annals<br />
and Magazine <strong>of</strong> Natural History, Series 7. 12: 430–448, pls<br />
2, 3.<br />
NORMAN, A.M. 905a: Revised nomenclature <strong>of</strong> the species<br />
described in Bate and Westwood’s `British sessile-eyed<br />
<strong>Crustacea</strong>’. Annals and Magazine <strong>of</strong> Natural History ser. 7:<br />
7. 16: 77–95.<br />
NORMAN, A.M. 905b: III. <strong>Crustacea</strong>. Pp. –47 in: Musem<br />
Normanium or A catalogue <strong>of</strong> the Invertebrata <strong>of</strong> the Arctic<br />
and North Atlantic Temperate Ocean and Paloearctic Region<br />
which are contained in the collection <strong>of</strong> the Rev. Canon A.M.<br />
Norman. Thos. Caldeugh & Son, Durham.<br />
NOVOTNy, A.J.; MAHNKEN, C.V.W. 97 : Predation on<br />
juvenile Pacific salmon by Rocinela belliceps pugettensis<br />
(<strong>Crustacea</strong>: <strong>Isopoda</strong>). U.S. National <strong>Marine</strong> Fisheries Service<br />
Fishery Bulletin 69: 699–70 .<br />
NUNOMURA, N. 1981: Three species <strong>of</strong> flabelliferan isopods<br />
(<strong>Crustacea</strong>) from the East China Sea, including the<br />
description <strong>of</strong> a new species <strong>of</strong> Syscenus. Bulletin <strong>of</strong> the<br />
Toyama Science Museum 3: 3– 8.<br />
NUNOMURA, N. 988a: A new species <strong>of</strong> the genus<br />
Aega (<strong>Crustacea</strong> <strong>Isopoda</strong>) from the sea <strong>of</strong>f Okinawa. Bulletin<br />
<strong>of</strong> the Toyama Science Museum 12: 9–22.<br />
NUNOMURA, N. 988b: A new aegid isopod (<strong>Crustacea</strong>)<br />
collected from a glass sponge. Bulletin <strong>of</strong> the Toyama Science<br />
Museum 12: 23–26.<br />
NUNOMURA, N. 993: A new species <strong>of</strong> the genus Aega<br />
from Toyama Bay. Bulletin <strong>of</strong> the Toyama Science Museum<br />
16: – 4.<br />
NUNOMURA, N. 2005: <strong>Marine</strong> isopod crustaceans collected<br />
from Breid Bay and Lützow-holm Bay, Antarctica, during<br />
JARE-26 cruise. Bulletin <strong>of</strong> the Toyama Science Museum<br />
28: 63–80.<br />
NUNOMURA, N. 2006: <strong>Marine</strong> isopod crustaceans in the<br />
Sagami Sea, Central Japan. Memoirs <strong>of</strong> the National Science<br />
Museum, Tokyo, 41: –42.<br />
PILGRIM, R.L.C. 2005: Chilton, Charles 860– 929. http://<br />
www.dnzb.govt.nz/; date accessed, November 2005.<br />
PILLAI, N.K. 954: A preliminary note on the Tanaidacea<br />
and <strong>Isopoda</strong> <strong>of</strong> Travancore. Bulletin <strong>of</strong> the Central Research<br />
Institute, University <strong>of</strong> Kerala, India 3(1): –2 .<br />
PILLAI, N.K. 967: Littoral and parasitic isopods from Kerala:<br />
Families Eurydicidae, Corallanidae and <strong>Aegidae</strong> - 2. Journal<br />
<strong>of</strong> the Bombay Natural History Society 64(2): 267–283.<br />
POLZ, H. 2005: Zwei neue Asselarten (<strong>Crustacea</strong>: <strong>Isopoda</strong>: Scutocoxifera)<br />
aus den Plattenkalken von Brunn (Oberkimmeridgium,<br />
Mittlere Frankenalb). Archaeopteryx 23: 67–8 .<br />
POORE, G.C.B. 2005: Supplement to the 2002 catalogue <strong>of</strong><br />
Australian <strong>Crustacea</strong>: Malacostraca — Syncarida and<br />
Peracarida (Volume 9.2A): 2002–2004. Museum Victoria<br />
Science Reports 7: – 5.<br />
POORE, G.C.B.; BRUCE, N.L. in press: Order <strong>Isopoda</strong>—slaters,<br />
fish lice and kin. In: Gordon, D.P. (Ed.) <strong>The</strong> <strong>New</strong> Zea-<br />
228<br />
land Inventory <strong>of</strong> Biodiversity. Volume 2: Kingdom Animalia<br />
— Chaetognatha, Ecdysozoa, and Ichn<strong>of</strong>ossils. Canterbury<br />
University Press, Christchurch.<br />
RAFI, F. 985: Synopsis Speciorum. <strong>Crustacea</strong>: <strong>Isopoda</strong> et<br />
Tanaidacea. Bibliographia Invertebratorum Aquaticorum<br />
Canadensium 4: –50.<br />
RATHKE, H. 837: Beitrage zur <strong>Fauna</strong> der Krym. Mémoires<br />
présentes à l’Académie Impériale des Sciences, St Petersbourg<br />
par des Savants Étrangers du Académie des Sciences, St<br />
Pétersbourg 3: 29 –454, 0 pl.<br />
RICHARDSON, H. 898: Description <strong>of</strong> four new species <strong>of</strong><br />
Rocinela, with a synopsis <strong>of</strong> the genus. Proceedings <strong>of</strong> the<br />
American Philosophical Society 38: 8– 7.<br />
RICHARDSON, H. 903: Isopods collected at the Hawaiian<br />
Islands by the U.S. Fish Commission Steamer Albatross.<br />
Bulletin <strong>of</strong> the United States Fish Commission 23(3):<br />
8 7–826.<br />
RICHARDSON, H. 904a. Contributions to the natural history<br />
<strong>of</strong> the <strong>Isopoda</strong>. Proceedings <strong>of</strong> the United States National<br />
Museum 27: –89.<br />
RICHARDSON, H. 904b. Contribution to the natural History<br />
<strong>of</strong> the <strong>Isopoda</strong>. VI. Isopods collected at the Hawaiian<br />
Islands by the U.S. Fish Commission Steamer Albatross.<br />
Proceedings <strong>of</strong> the United States National Museum 27:<br />
67 –68 .<br />
RICHARDSON, H. 905a: A monograph on the isopods<br />
<strong>of</strong> North America. Bulletin <strong>of</strong> the United States National<br />
Museum 54: vii–liii, –727.<br />
RICHARDSON, H. 905b: Isopods <strong>of</strong> the Alaska salmon investigation.<br />
Bulletin <strong>of</strong> the Bureau <strong>of</strong> Fisheries 24: 209–22 .<br />
RICHARDSON, H. 906a: Sur les isopodes de l’Expédition<br />
Française Antarctique. Bulletin du Muséum d’Histoire<br />
Naturelle, Paris 12(4): 87– 89.<br />
RICHARDSON, H. 906b: Sur les isopodes de l’Expédition<br />
Française Antarctique. Comptes Rendus des Séances de<br />
l’Académie des Sciences 142(14): 849–85 .<br />
RICHARDSON, H. 908: Isopodes. Pp. –22 in: Expédition<br />
Antarctique Français 1903–1905 commandée par Dr Jean<br />
Charcot. Pl I. Masson et Cie, Paris.<br />
RICHARDSON, H. 909: Isopods collected in the Northwest<br />
Pacific by the U.S. Bureau <strong>of</strong> Fisheries Steamer ‘Albatross’<br />
in 906. Proceedings <strong>of</strong> the U.S. National Museum 37(1701):<br />
75– 29.<br />
RICHARDSON, H. 9 0: <strong>Marine</strong> isopods collected in the<br />
Philippines by the U.S. Fisheries steamer Albatross in<br />
907–08. Bureau <strong>of</strong> Fisheries Document 736: –44.<br />
RICHARDSON, H. 9 . Description <strong>of</strong> a new species <strong>of</strong> Aega<br />
from the Atlantic coast <strong>of</strong> the United States. Proceedings <strong>of</strong><br />
the United States National Museum 40: 623–624.<br />
RICHARDSON, H. 9 3: Crustacés Isopodes. Pp. –24 in:<br />
Deuxième Expédition Antarctique Française 1908–1910 commandée<br />
par le Dr J. Charcot. Masson et Cie, Paris.<br />
RISSO, A. 8 6: Histoire naturelle des crustacés des environs de<br />
Nice. D’Hautel, Paris. 75 p.<br />
ROHDE, K. 982: Ecology <strong>of</strong> <strong>Marine</strong> Parasites. University <strong>of</strong><br />
Queensland Press, Brisbane, Queensland. i–xvi, –245 p.
ROHDE, K. 2005: <strong>Marine</strong> Parasitology. CSIRO, Melbourne.<br />
–592 p.<br />
ROMAN, M.L.; DALENS, H. 999: <strong>Isopoda</strong> order (excluding<br />
Epicaridea), (<strong>Isopoda</strong> Latreille, 8 7). Pp. 77–278<br />
in: Traité de Zoologie: Anatomie, Systematique, Biologie.<br />
Tome 7, fascicule 3A: Crustacés Peracaridés,<br />
Musée Oéanographique, Monaco. (Mémoires de l’Institut<br />
Océanographique de Monaco). l’Institut Océanographique,<br />
Monaco.<br />
ROSS, S.W.; SULAK, K.J.; MUNROE, T.A. 200 : Association<br />
<strong>of</strong> Syscenus infelix (<strong>Crustacea</strong>: <strong>Isopoda</strong>:<br />
<strong>Aegidae</strong>) with benthopelagic rattail fishes,<br />
Nezumia spp. (Macrouridae), along the western North Atlantic<br />
continental slope. <strong>Marine</strong> Biology 138(3): 595–602.<br />
SAITO, N.; ITANI, N.; NUNOMURA, N. 2000: A preliminary<br />
checklist <strong>of</strong> isopod crustacean from Japan. Bulletin <strong>of</strong> the<br />
Toyama Science Museum 23: – 07.<br />
SARS, G.O. 882: Oversigt af Norges Crustaceer med<br />
foreløbige Bemærkninger over de nye eller mindre<br />
bekjendte Arter. I. (Podophthalmata–Cumacea–<strong>Isopoda</strong>–<br />
Amphipoda). Forhandlinger I Videnskabs-Selskabet I Kristiania,<br />
1882(18): – 24.<br />
SARS, G.O. 897: <strong>Isopoda</strong>. Part III, IV. Anthuridae, Gnathiidae,<br />
<strong>Aegidae</strong>, Cirolanidae, Limnoriidae. An Account <strong>of</strong><br />
the <strong>Crustacea</strong> <strong>of</strong> Norway with Short Descriptions and Figures<br />
<strong>of</strong> All the Species. Bergen Museum, Bergen, Norway, 2:<br />
4 –80.<br />
SARS, G.O. 899: Volume II. <strong>Isopoda</strong>, parts 3, 4. An Account<br />
<strong>of</strong> the <strong>Crustacea</strong> <strong>of</strong> Norway with Short Descriptions<br />
and Figures <strong>of</strong> All the Species. Bergen Museum, Christiana.<br />
264 p.<br />
SARS, M. 859: Oversigt over de i den norsk-arctiske region<br />
forekommende Krebsdyr. Forhandlinger I Videnskabs-<br />
Selskabet I Kristiania 1858: 22– 63.<br />
SCHIOEDTE, J.C.; MEINERT, F. 879a: De cirolanis Ægas<br />
simulantibus. Commentatio brevis. Naturhistorisk<br />
Tidsskrift, Kjøbenhavn 3: 279–302, pls 3–5.<br />
SCHIOEDTE, J.C.; MEINERT, F. 879b: Symbolæ ad monographium<br />
Cymothoarum crustaceorum isopodum familiæ.<br />
I. Aegidæ. Naturhistorisk Tidsskrift, Kjøbenhavn 12:<br />
32 –4 4, pls 7– 3.<br />
SCHULTZ, G.A. 969: How to know the marine isopod crustaceans.<br />
Wm. C. Brown, Iowa. 359 p.<br />
SCHULTZ, G.A. 978: Nonasellote isopod crustaceans from<br />
Anvers Island and other Antarctic locations. Biology <strong>of</strong><br />
the Antarctic Seas VIII. Antarctic Research Series 28(2):<br />
2 –4 .<br />
SEMPER, C. 867: Wiegmann’s Archiv für Naturgeschichte 33:<br />
84–89 [not sighted].<br />
SMITH, W. 867: Dictionary <strong>of</strong> Greek and Roman Biography<br />
and Mythology. http://www.ancientlibrary.com/index.<br />
php; date accessed, October 2006.<br />
SOUTHWELL, T. 9 5: Notes from the Bengal Fisheries Laboratory,<br />
Indian Museum, No. 2. On some Indian parasites<br />
<strong>of</strong> fish, with a note on carcinoma in trout. Records <strong>of</strong> the<br />
Indian Museum 11: 3 –330, pls26–28.<br />
SPRINGTHORPE, R.; LOWRy, J.K. 994: Catalogue <strong>of</strong><br />
crustacean type specimens in the Australian Museum:<br />
229<br />
Malacostraca. Technical Reports <strong>of</strong> the Australian Museum<br />
11. Australian Museum, Sydney, – 34 p.<br />
STEBBING, T.R.R. 893: A History <strong>of</strong> <strong>Crustacea</strong>. Recent Malacostraca.<br />
<strong>The</strong> International Scientific Series. Kegan Paul,<br />
Trench, Trübner & Co. Ltd., London, xvii, 466 p.<br />
STEBBING, T.R.R. 905: Report on the <strong>Isopoda</strong> collected by<br />
Pr<strong>of</strong>essor Herdman, at Ceylon, in 902. Report to the Government<br />
<strong>of</strong> Ceylon on the Pearl Oyster Fisheries <strong>of</strong> the Gulf <strong>of</strong><br />
Manaar, 1905 Supplementary Report 4(23): 47–64.<br />
STEBBING, T.R.R. 9 0: <strong>Isopoda</strong> from the Indian Ocean and<br />
British East Africa. <strong>The</strong> Percy Sladen Trust Expedition to<br />
the Indian Ocean under the leadership <strong>of</strong> Mr J. Stanley<br />
Gardiner. Volume III. Transactions <strong>of</strong> the Linnean Society<br />
<strong>of</strong> London (Zoology) 14: 83– 22, pls 5– .<br />
STEBBING, T.R.R. 920: <strong>Crustacea</strong> from the Falkland Islands<br />
collected by Mr Rupert Vallentin, F.L.S—Part III. Proceedings<br />
<strong>of</strong> the Zoological Society <strong>of</strong> London: 327–340, pls. –5.<br />
STEBBING, T.R.R. 922: <strong>Isopoda</strong> and Amphipoda from<br />
Angola and South Africa. Goteborgs Kungl vetenskaps- och<br />
vitterhets-samhalles Handlingar 25: – 6, pls i–iv.<br />
STEBBING, T.R.R. 924, for 922: <strong>Crustacea</strong> <strong>of</strong> Natal. Union <strong>of</strong><br />
South Africa, Fisheries and <strong>Marine</strong> Biological Survey, Report<br />
3 (Special Reports III): – 5.<br />
STEPHENSEN, K. 937: <strong>Marine</strong> <strong>Isopoda</strong> and Tanaidacea.<br />
Zoology <strong>of</strong> Iceland 27: –26.<br />
STEPHENSEN, K. 947: Tanaidacea, <strong>Isopoda</strong>, Amphipoda,<br />
and Pycnogonida. Scientific Results <strong>of</strong> the Norwegian Antarctic<br />
Expedition, 1927–1928 27: –90.<br />
STEPHENSEN, K. 948: Storkrebs IV. Ringkrebs. 3. Tanglus<br />
(<strong>Marine</strong> Isopoder) og Tanaider. Pp. – 87 in: Danmarks<br />
<strong>Fauna</strong>. G.E.G. Gads Forlag, København.<br />
STEPHENSON, A.B. 980: Aega angustata Whitelegge, 90<br />
(<strong>Isopoda</strong>: <strong>Aegidae</strong>), a new record for <strong>New</strong> <strong>Zealand</strong><br />
waters. Records <strong>of</strong> the Auckland Institute and Museum 17:<br />
53– 55.<br />
STIMPSON, W. 864: Descriptions <strong>of</strong> new species <strong>of</strong> marine<br />
invertebrates from Puget Sound. Proceedings <strong>of</strong> the Philadelphia<br />
Academy <strong>of</strong> Sciences 16: 53– 6 .<br />
STUDER, T. 884: Isopoden, gesammelt während der Reise<br />
S.M.S. Gazelle um die Erde 874–76. Abhandlungen Koniglich<br />
Preussischen Akademis der Wissenschaften, Berlin<br />
1883(1): –28, 2 pls.<br />
SVAVARSSON, J.; BRUCE, N.L. 2000: Redescription <strong>of</strong> the<br />
cosmopolitan meso- and bathypelagic cirolanid Metacirolana<br />
caeca (Hansen, 9 6) comb. nov. (<strong>Crustacea</strong>, <strong>Isopoda</strong>).<br />
Steenstrupia 25(2): 47– 58.<br />
SWOFFORD, D.L. 2004: PAUP*. Phylogenetic Analysis Using<br />
Parsimony (*and other methods). Version 4.0b 0. Sinauer<br />
Associates, 42 p.<br />
TATTERSALL, W.M. 909: II. Amphipoda and <strong>Isopoda</strong>, with<br />
descriptions <strong>of</strong> two new species. Memoirs <strong>of</strong> the Challenger<br />
Society 1: 2 0–2 9.<br />
TATTERSALL, W.M. 9 : Die Nordischen Isopoden. Pp.<br />
8 –3 3 in: Apstein (Ed.) (Nordischen Plankton) Lipsiusand<br />
Tischer, Kiel and Leipzig.<br />
TATTERSALL, W.M. 92 : <strong>Crustacea</strong>. Part VI. Tanaidacea<br />
and <strong>Isopoda</strong>. In: British Antarctic “Terra Nova” Expe-
dition 9 0. Natural History Report, Zoology, 3(8). pp<br />
9 –258, pls. – . British Museum, Natural History,<br />
London.<br />
THIELEMANN, M. 9 0: Beiträge zur Kenntnis der Naturgechichte<br />
Ostasiens. Herausgegeben von F. D<strong>of</strong>lein. Band<br />
II. No. 9. Beiträge zur Kenntnis der Isopodenfauna<br />
Ostasiens. Abhandlungen der Mathematisch-Naturwissenschaftlichen<br />
Klasse der K. Bayer. Akademia der Wissenschaften<br />
(suppl), 2(3): – 09, 2 pls.<br />
THOMSON, G.M. 884: Descriptions <strong>of</strong> new crustaceans.<br />
Transactions <strong>of</strong> the <strong>New</strong> <strong>Zealand</strong> Institute, Zoology 16:<br />
234–240.<br />
THOMSON, G.M. 9 3: <strong>The</strong> natural history <strong>of</strong> Otago Harbour<br />
and the adjacent sea with a record <strong>of</strong> the researches<br />
carried on at the Portobello <strong>Marine</strong> Fish-hatchery: Part<br />
. Transactions and Proceedings <strong>of</strong> <strong>New</strong> <strong>Zealand</strong> Institute<br />
45: 225–25 .<br />
THOMSON, G.M.; CHILTON, C. 886: Critical list <strong>of</strong> the<br />
<strong>Crustacea</strong> Malacostraca <strong>of</strong> <strong>New</strong> <strong>Zealand</strong>. Transactions and<br />
Proceedings <strong>of</strong> the <strong>New</strong> <strong>Zealand</strong> Institute 18: 4 – 59.<br />
TRACEy, D.; ANDERSON, O.F.; CLARK, M.R.; OLIVER,<br />
M.D. 2005: A guide to common deepsea invertebrates<br />
in <strong>New</strong> <strong>Zealand</strong> waters. <strong>New</strong> <strong>Zealand</strong> Aquatic Environment<br />
and Biodiversity Report 1. Ministry <strong>of</strong> Fisheries,<br />
Wellington, – 60 p.<br />
TREAT, S.A.F. 980: <strong>New</strong> record <strong>of</strong> Aega monophthalma Johnston<br />
(<strong>Isopoda</strong>: Flabellifera: <strong>Aegidae</strong>) in the tropical western<br />
Atlantic. Bulletin <strong>of</strong> <strong>Marine</strong> Science 30(4): 9 2–9 4.<br />
TRILLES, J.P.; JUSTINE, J.L. 2004: Une nouvelle espèce de Cymothoidae<br />
et trois <strong>Aegidae</strong> (<strong>Crustacea</strong>, <strong>Isopoda</strong>) récoltés<br />
sur des poissons de pr<strong>of</strong>ondeur au large de la Nouvelle<br />
Calédonie. Zoosystema 26(2): 2 –233.<br />
VAN NAME, W.G. 924: Isopods from the Williams Galapagos<br />
Expedition. Zoologica, Scientific Contributions <strong>of</strong> the<br />
<strong>New</strong> York Zoological Society 5(18): 8 –2 0.<br />
VASINA, G.S. 993: A new species <strong>of</strong> the genus Rocinela<br />
(<strong>Crustacea</strong>, <strong>Isopoda</strong>, Flabellifera, <strong>Aegidae</strong>) from the Sea<br />
<strong>of</strong> Okhotsk. Biologiya Morya (Vladivostok) 1993(1): 40–43<br />
[In Russian].<br />
WÄGELE, J-W. 989: Evolution und phylogenetisches System<br />
der <strong>Isopoda</strong>. Stand der Forschung und neue Erkenntnisse.<br />
Zoologica 140: –262.<br />
WÄGELE, J.-W. 990: Growth in captivity and aspects<br />
<strong>of</strong> reproductive biology <strong>of</strong> the Antarctic fish parasite<br />
230<br />
Aega antarctica (<strong>Crustacea</strong>, <strong>Isopoda</strong>). Polar Biology 10(7):<br />
52 –527.<br />
WAHRBERG, R. 930: Sveriges marina och lacustra Isopoder.<br />
Göteborgs Kunglinga Vetenskaps-och Vitterhets-samhälles<br />
Handlinger, Femte Följder, Ser B 1 (9): –76. pls – 8.<br />
WATLING, L. 1989: A classification concept for crustacean<br />
setae based on the homology concept. Pp. 5–26 in: Felgenhauer,<br />
B.E.; Watling, L.; Thistle, A.B. (Eds). Functional<br />
morphology <strong>of</strong> feeding and grooming in <strong>Crustacea</strong>. <strong>Crustacea</strong>n<br />
Issues. A.A. Balkema, Rotterdam.<br />
WEIDER, R.; FELDMANN, R.M. 992: Mesozoic and cenozoic<br />
fossil isopods <strong>of</strong> North America. Journal <strong>of</strong> Paleontology<br />
66(6): 958–972.<br />
WETZER, R. 990: A new species <strong>of</strong> isopod, Aega (Rhamphion)<br />
francoisae (Flabellifera: <strong>Aegidae</strong>), from the cloaca <strong>of</strong> an<br />
ascidian from the Galapagos Islands. Proceedings <strong>of</strong> the<br />
Biological Society <strong>of</strong> Washington 103(3): 655–662.<br />
WETZER, R.; BRUSCA, R.C. 997: Descriptions <strong>of</strong> the species<br />
<strong>of</strong> the suborders Anthuridea, Epicaridea, Flabellifera,<br />
Gnathiidea, and Valvifera. Pp. 9–58, 0– 20 in: Blake,<br />
J.A.; Scott, P.H. (Eds). Taxonomic Atlas <strong>of</strong> the Benthic <strong>Fauna</strong><br />
<strong>of</strong> the Santa Maria Basin and Western Santa Barbara Channel.<br />
Vol. 11. <strong>The</strong> <strong>Crustacea</strong>, Part 2: <strong>Isopoda</strong>, Cumacea and<br />
Tanaidacea. Santa Barbara Museum <strong>of</strong> Natural History,<br />
Santa Barbara.<br />
WHITE, A. 847: List <strong>of</strong> species in the collection <strong>of</strong> the British<br />
Museum. British Museum, London, i–viii, – 43 pp.<br />
WHITE, A. 850: List <strong>of</strong> the specimens <strong>of</strong> British animals in the<br />
collection <strong>of</strong> the British Museum. Part IV.– <strong>Crustacea</strong>. British<br />
Museum, London.<br />
WHITELEGGE, T. 90 : <strong>Crustacea</strong>. Part II. <strong>Isopoda</strong>. Part I.<br />
Scientific results <strong>of</strong> the Trawling Expedition <strong>of</strong> HMCS<br />
“<strong>The</strong>tis”. Memoirs <strong>of</strong> the Australian Museum 1: 20 –24 .<br />
WING, B.A.; MOLES, D.L. 995: Behavior <strong>of</strong> Rocinela angustata<br />
(<strong>Isopoda</strong>, <strong>Aegidae</strong>), an ectoparasite <strong>of</strong> Alaskan marine<br />
fishes. Journal <strong>of</strong> Aquatic Animal Health 7(1): 34–37.<br />
WOODWARD, H. 870: Contribution to British fossil <strong>Crustacea</strong>.<br />
Geological Magazine 7: 493–497.<br />
yU, H. 2007: A new species <strong>of</strong> <strong>Aegidae</strong> (Isopod, Cymothoida)<br />
from the South China Sea. <strong>Crustacea</strong>na 80(8): 909–9 5.<br />
yU, H.; BRUCE, N.L. 2006: Aega sheni, a new species <strong>of</strong> <strong>Aegidae</strong><br />
(<strong>Crustacea</strong>: <strong>Isopoda</strong>: Cymothoida) from southern<br />
China and Australia. Zootaxa 1224: 23–3 .
AppENdIx 1. INvAlId AEgId NAmEs<br />
Included here are species no longer in the family as a consequence <strong>of</strong> being junior synonyms, relegated to species<br />
inquirenda or nomen dubium, or having been transferred to other combinations or placed in other families.<br />
Aega affinis H. Milne Edwards, 840; no locality was<br />
given by Milne Edwards ( 840) for this species<br />
which, with only four lines <strong>of</strong> description, remains<br />
incertae sedis [Milne Edwards also used<br />
the ‘popular’ name <strong>of</strong> Aega voisine]; regarded as a<br />
junior synonym <strong>of</strong> Aega psora by Kussakin ( 979).<br />
This is not Rocinela affinis Richardson, 904a.<br />
Aega alaskensis Lockington, 877; now in Rocinela.<br />
Aega australis Richardson, 906b (and Richardson,<br />
908); junior homonym <strong>of</strong> A. australis Whitelegge,<br />
90 ; = Aega antarctica Hodgson, 9 0.<br />
Aega basalis Heller, 868; Corallana (Corallanidae).<br />
Aega belliceps Stimpson, 864; now in Rocinela.<br />
Aega bicavata Nordenstam, 930; = Aega semicarinata<br />
Miers, 875.<br />
Aega dubia Richardson, 9 0; = Aega vigilans Haswell,<br />
88 .<br />
Aega edwardsii Dollfus, 89 ; = Aega punctulata Miers,<br />
88 (present study, see Appendix 2, p. 235).<br />
Aega efferata Dana, 853; nomen dubium.<br />
Aega emarginata Leach, 8 5; = Aega psora Linnaeus,<br />
758.<br />
Aega entailee Latreille, 829; = Aega psora Linnaeus,<br />
758.<br />
Aega giganteocula Nunomura, 988a; = Aegiochus vigilans<br />
Haswell, 88 (present study, p. 50).<br />
Aega harfordi Lockington, 877; long placed in Cirolana<br />
Leach, 8 8 (Cirolanidae).<br />
Aega hirta White, 847; nomen nudum; subsequently<br />
cited by Hansen ( 890) and listed by Nierstrasz<br />
( 93 ); see Clarke and Preswell (200 ).<br />
Aega interrupta von Martens, 868; = Alitropus typus<br />
Milne Edwards, 840.<br />
Aega koltuni Kussakin, 967; = Aega antarctica Hodgson,<br />
9 0 (present study, p. 226).<br />
Aega loveni Bovallius, 886; = Aega ventrosa Sars,<br />
859.<br />
Aega macronema Bleeker, 857; now in Argathona (Corallanidae).<br />
Aega magnoculis Richardson, 909; = Aega plebeia<br />
Hansen, 897.<br />
Aega maorum Filhol, 885; = Pseudaega punctata (Thomson,<br />
884) (Cirolanidae).<br />
Aega meinerti Miers, 884; = Aega serripes Milne Edwards,<br />
840.<br />
Aega multidigita Dana, 853; now in Alcirona (Corallanidae).<br />
Aega novizaelandiae Dana, 853; nomen dubium (present<br />
study, p. 2 ).<br />
Aega ommatophylax Stebbing, 905; = Aega vigilans<br />
Haswell, 88 .<br />
Aega ornata Richardson, 9 ; now in Tridentella Richardson,<br />
905 (Tridentellidae Bruce, 984).<br />
23<br />
Aega schioedteana Bovallius, 885; = Aega deshaysiana<br />
(Milne Edwards, 840).<br />
Aega stroemii Lütken, 859; = Aega bicarinata Rathke,<br />
837.<br />
Aega tumida Nunomura, 988b; there is little to differentiate<br />
this species from the poorly known<br />
Aegiochus spongiophila (Semper, 867), the likelihood<br />
<strong>of</strong> A. tumida being a junior synonym being<br />
further strengthened by the fact that both species<br />
are known only from ‘glass’ sponges (Hexactinellidae).<br />
Acherusia rotundicauda Lilljeborg, 85 ; = Rocinela danmoniensis<br />
Leach, 8 8.<br />
Acherusia complanata Grube, 864; = Rocinela dumerilii<br />
(Lucas, 849)<br />
Aegiochus nordenskjoldii Bovallius, 885; = Aega ventrosa<br />
M. Sars, 859.<br />
Alitropus dimorphus Pillai, 954; = Alitropus typus Milne<br />
Edwards, 840.<br />
Alitropus foveolatus Schioedte and Meinert, 879b; =<br />
Alitropus typus Milne Edwards, 840.<br />
Harponyx pranizoides Sars, 882; = Syscenus infelix,<br />
Harger, 880.<br />
Rocinela aries Schioedte and Meinert, 879b; = Rocinela<br />
signata Schioedte and Meinert, 879 (see Brusca<br />
& France 992).<br />
Rocinela alascensis (Stimpson, 864); = Rocinela belliceps<br />
(Stimpson, 864).<br />
Rocinela deshaysiana Milne Edwards, 840; long placed<br />
in Aega; now in Aegapheles.<br />
Rocinela latis Southwell, 1915: 321, figs 12–15; a species<br />
<strong>of</strong> Nerocila, aegathoid stage, from Lates calcarifer.<br />
Rocinela lilljeborgii Bovallius, 885; = Syscenus infelix,<br />
Harger, 880.<br />
Rocinela major Brocchi, 877; southern Indian Ocean,<br />
St. Paul Island; the identity <strong>of</strong> this species is entirely<br />
unknown and there is no information on<br />
the whereabouts <strong>of</strong> any potential type material;<br />
here regarded as nomen dubium.<br />
Rocinela mundana Lanchester, 902; = Alitropus typus<br />
Milne Edwards, 840.<br />
Rocinela ophthalmica Milne Edwards, 840. Type locality<br />
Sicily. No further data than that given by Milne<br />
Edwards ( 840) are available. Milne Edwards<br />
stated that Aega deshaysiana (then as Rocinela<br />
deshaysiana) was very similar to this species, and<br />
it is possible that it is a species <strong>of</strong> Aega. Species<br />
inquirenda.<br />
Rocinela simplex Chilton, 926; = Alitropus typus Milne<br />
Edwards, 840.
Aega angustata Whitelegge, 90 (Figs 39, 40)<br />
Aega angustata Whitelegge, 1901: 232, fig. 21a–21f.– Hale,<br />
1925: 170, fig. 20.– Nierstrasz, 1931: 182.– Bruce, Lew<br />
Ton & Poore, 2002: 60.<br />
Aega (Aega) angustata.– Brusca, 983: 0.<br />
Not Aega angustata.– Stephenson, 980: 53, figs –5<br />
(misidentification, = Aega komai Bruce, 996).<br />
Not Aega angustata.– Trilles & Justine, 2004: 220, figs 6, 7<br />
(misidentification, = undescribed species).<br />
materiaL examined: Holotype, ♂ ( 4.3 mm), 5.5–6.5 km<br />
<strong>of</strong>f Wattamolla, NSW, 34° 0’S, 5 ° ’E, 22 March<br />
898, stn 57, 99– 08 m, coll. E.R. Waite on HMCS <strong>The</strong>tis<br />
(AM G2 60). ♂ ( 7.0 mm), Bass Strait, 8 -HK- , stn<br />
48/29, from Raja (NMV J8878). ♂ ( 7.5 mm), BSS stn<br />
57 (NMV J8882).<br />
desCription: Body 3.7 times as long as greatest width,<br />
dorsal surfaces smooth or polished in appearance, widest<br />
at pereonite 5 or pereonite 6, lateral margins subparallel.<br />
Rostral point projecting anteriorly, not ventrally<br />
folded. Eyes large, not medially united, separated by<br />
about 30% width <strong>of</strong> head. Pereonite 1 and coxae 2–3 each<br />
with posteroventral angle rounded. Coxae 5–7 with<br />
entire oblique carina; posterior margins convex, posterolateral<br />
angle rounded. Pleon with pleonite largely<br />
concealed by pereonite 7; pleonite 4 with posterolateral<br />
margins extending clearly beyond posterior margin <strong>of</strong><br />
pleonite 5; pleonite 5 with posterolateral angles free,<br />
not overlapped by lateral margins <strong>of</strong> pleonite 4. Pleotelson<br />
0.9 times as long as anterior width, dorsal surface<br />
with longitudinal carina on distal third; lateral margins<br />
weakly convex, deeply serrate, posterior margin with<br />
distinct short median point, with 0 RS.<br />
Antennule peduncle articles 1 and 2 flattened, article<br />
2 anterodistal lobe extending to end <strong>of</strong> article 3; articles<br />
3 and 4 0.3 times as long as combined lengths <strong>of</strong> articles<br />
1 and 2, article 3 3.1 times as long as wide; flagellum<br />
with 5 articles, extending to mid-point <strong>of</strong> eye. Antenna<br />
peduncle article 2 inferior surface without distinct<br />
longitudinal suture; article 5 flattened and expanded,<br />
2.1 times as long as article 4 (in situ); flagellum with 8<br />
articles, extending to posterior <strong>of</strong> pereonite .<br />
Frontal lamina flat, longer than greatest width, lateral<br />
margins converging posteriorly, anterior margin<br />
rounded, without small median point, posterior margin<br />
not abutting clypeus.<br />
Pereopod 1 basis .9 times as long as greatest width;<br />
ischium 0.5 times as long as basis, inferior margin with<br />
AppENdIx 2. ExtrA-lImItAl spEcIEs<br />
<strong>The</strong> species included here are some <strong>of</strong> those in need <strong>of</strong> at least partial redescription because <strong>of</strong> their similarity to<br />
<strong>New</strong> <strong>Zealand</strong> species in the body <strong>of</strong> the monograph. Some <strong>of</strong> these have been placed in synonymy with older<br />
names at some point. For most species the descriptive notes are based solely on the type material.<br />
232<br />
0 RS, superior distal margin with 2 RS; merus inferior<br />
margin convex and thickened, merus inferior margin<br />
with 2 RS (small), set as distal group, superior distal<br />
angle with 2 RS; carpus 0.7 as long as merus, inferior<br />
margin with 0 RS; propodus .3 times as long as proximal<br />
width, inferior margin with RS (distal), propodal<br />
palm simple, without blade or process, dactylus<br />
smoothly curved, .3 as long as propodus. Pereopod<br />
3 ischium inferior margin with 3 RS, superior distal<br />
margin with RS; merus inferior margin with 5 RS<br />
(set as 3 + 2), set as two groups, superior distal margin<br />
with 2 acute RS; carpus longer than that <strong>of</strong> pereopod<br />
, with inferodistal lobe, inferodistal angle with RS,<br />
propodus with large club-shaped distal RS. Pereopods<br />
5–7 inferior margins <strong>of</strong> ischium–carpus with long acute<br />
RS. Pereopod 6 basis 2.9 times as long as greatest width,<br />
inferior margins with 6 palmate setae; ischium 0.6 as<br />
long as basis, inferior margin with 5 RS (set loosely as<br />
, , 2 and ), superior distal angle with 5 RS, inferior<br />
distal angle with 8 RS; merus 0.7 as long as ischium, 2.0<br />
as long as wide, inferior margin with 6 RS (set loosely<br />
as 3 and 3, 4 being submarginal), superior distal angle<br />
with 5 RS, inferior distal angle with 5 RS; carpus 0.9<br />
as long as ischium, 3.6 times as long as wide, inferior<br />
margin with 4 RS (set as and 3), superior distal angle<br />
with 3 RS, inferior distal angle with 6 RS; propodus .0<br />
as long as ischium, 6.8 times as long as wide, inferior<br />
margin with 3 RS (set as and 2), superior distal angle<br />
with slender seta, inferior distal angle with 4 RS.<br />
Penes short rectangular lobes; penial openings<br />
separated by % <strong>of</strong> sternal width, penial process 3.5<br />
times as long as basal width.<br />
Pleopod 1 exopod .8 times as long as wide, distally<br />
broadly rounded, lateral margin straight, mesial margin<br />
weakly convex, with PMS on distal two-thirds;<br />
endopod .5 times as long as wide, distally narrowly<br />
rounded, lateral margin weakly concave, with PMS on<br />
distal half, mesial margin with PMS on distal one-third;<br />
peduncle .5 times as wide as long, mesial margin with<br />
6 coupling hooks. Exopods <strong>of</strong> pleopods –3 each with<br />
distolateral margin not digitate.<br />
Uropod peduncle ventrolateral margin with 2 RS,<br />
posterior lobe about as long as endopod. Uropod<br />
rami with endopod and exopod co-planar, rami not<br />
extending beyond pleotelson, marginal setae in single<br />
tier, apices acute. Endopod apically not bifid, lateral<br />
margin proximally convex and distally convex, without<br />
prominent excision, proximal lateral margin with 0 RS,
Figure 139. Aega angustata Whitelegge, 90 . Holotype. A, dorsal view; B, lateral view; C, head; D, frons; E, pleonites, lateral<br />
view; F, pleotelson; G, sternite 7; H, uropod; I, antennule.<br />
233
Figure 140. Aega angustata Whitelegge, 90 . Holotype. A–C, pereopods , 3 and 6, respectively; D, pleopod .<br />
distal lateral margin with RS, mesial margin straight<br />
(deeply serrate), with 4 RS. Exopod not extending to<br />
end <strong>of</strong> endopod, 3.8 times as long as greatest width,<br />
apically not bifid; lateral margin weakly sinuate, with<br />
8 RS (prominent); mesial margin sinuate, proximally<br />
concave, with 2 RS.<br />
remarks: An abbreviated description <strong>of</strong> the holotype<br />
is given here to facilitate identification <strong>of</strong> this species.<br />
<strong>The</strong> specimen was not further dissected, consequently<br />
descriptive details were taken from pereopods 3 and 6<br />
rather than the usual 2 and 7. Aega angustata is readily<br />
234<br />
identified by the elongate body, antennule peduncle<br />
articles and 2 being strongly compressed and expanded,<br />
antenna peduncle article 5 expanded, pereopod<br />
3 with a large robust seta opposing the dactylus,<br />
the serrate pleotelson posterior margin, uropods with<br />
the rami not extending posterior to the pleotelson apex,<br />
the uropodal endopod with a truncate and irregularly<br />
serrate posterior margin and the lateral margin <strong>of</strong> the<br />
uropodal exopod with prominent robust setae.<br />
<strong>The</strong>re are several similar species (see remarks for<br />
Aega komai, p. 37), and A. angustata is immediately<br />
separated from those species by the lateral margin
<strong>of</strong> the uropodal exopod lacking serrations and having<br />
prominent robust setae. Trilles and Justine (2004)<br />
figured a specimen from <strong>New</strong> Caledonia as having<br />
deeply serrate lateral margins to the uropodal endopod<br />
and exopod, those margins also lacking robust setae,<br />
indicating clearly that their material is not A. angustata<br />
(there are further differences in pleopod setation but<br />
the illustrations are not <strong>of</strong> a standard that permits<br />
confident interpretation).<br />
distribution: Known from southeastern Australia with<br />
records from <strong>of</strong>f Sydney to the Bass Strait, Victoria.<br />
Aega punctulata Miers, 88 (Fig. 4 )<br />
Aega edwardsii White, 847: 07 (nomen dubium; also nomen<br />
nudum, see Clark & Preswell 200 ).<br />
Æga punctulata Miers, 1881: 77, pl. 7, figs 10–12.<br />
Æga edwardsii Dollfus, 1891: F58, pl. VIII, fig. 3a–d (new<br />
synonymy).<br />
Aega punctulata.– Nierstrasz, 93 : 84.<br />
Aega (Aega) punctulata.– Brusca, 983: .<br />
?Aega edwardsi.– Kussakin & Vasina, 1980: 359, fig. 1 [identity<br />
uncertain, see ‘remarks’].<br />
Not Aega punctulata.– Hale, 1937: 17, fig. 5.– Bruce, Lew Ton<br />
& Poore, 2003: 161 [misidentification, see ‘remarks’].<br />
Not Aega cf. punctulata.– Barnard, 1960, 95, fig. 2 [identity<br />
uncertain, see ‘remarks’].<br />
materiaL examined: Holotype: ♂ (non-ovig. 29 mm),<br />
Wolsely Sound, Straits <strong>of</strong> Magellan, H.M.S. Alert<br />
(BMNH 79. 8).<br />
Non-type: ♀ (non-ovig. 33 mm), Port Stanley, Falkland<br />
Is., from mullet’s gills, coll. A.G. Bennett (BMNH<br />
920.7.5.2).<br />
Aega edwardsii, ♂ (20 mm), ♀ (non-ovig. 26 mm),<br />
syntypes; label data: “type (Miss. Sc.du Cap Horn, 8:vi,<br />
Zool, Crust, p. 28, ’63)” (MNHN Is.2437). [Type locality<br />
is ‘Baie Orange, Cape Horn’ (Dollfus 89 ).]<br />
desCriptiVe notes: Eyes small, separated by 4 % width<br />
<strong>of</strong> head. Body dorsal surfaces coarsely punctate, with<br />
abundant stiff setae, these being most dense posteriorly.<br />
Frontal lamina anteriorly rounded, posterior<br />
margin abutting labrum. Pleotelson posterior margins<br />
angled, forming shallow median point, provided with<br />
0– 2 (as 5+5 or 6+6) RS. Antenna peduncle article 5<br />
slightly shorter than article 4. Pereopods –3 propodal<br />
palm with small distal lobe. Pereopod merus inferior<br />
margin with 2+3 and +3 RS; pereopod 2 merus inferior<br />
margin 4+5 RS, arranged as a single proximal row<br />
<strong>of</strong> 4 RS and distal double row; pereopod 3 similar to<br />
pereopod 4. Uropod rami extending slightly beyond<br />
posterior margin <strong>of</strong> pleotelson; dorsal surfaces with<br />
stiff setae; uropod endopod mesial margin with 7 or 8<br />
RS, lateral with +2 RS; uropod exopod mesial with 3<br />
or 4 RS lateral margin with 9 or 0 RS.<br />
235<br />
remarks: Aega punctulata can be immediately identified<br />
by the prominent, stiff setae over the dorsal body<br />
surfaces, these setae being longest on pereonites 6 and<br />
7, pleon and pleotelson; other distinguishing characters<br />
include the relatively small and widely separated<br />
eyes (<strong>of</strong> almost cirolanid proportions), short antennule<br />
(extending only to posterior <strong>of</strong> head), short antenna<br />
(extends to posterior <strong>of</strong> pereonite ), pattern <strong>of</strong> robust<br />
setae <strong>of</strong> the merus <strong>of</strong> pereopods –3, and the shape<br />
<strong>of</strong> the posterior margin <strong>of</strong> the pleotelson, which is<br />
indistinctly angled.<br />
<strong>The</strong> holotype is in poor condition, having lost the<br />
distal articles to all the anterior pereopods. <strong>The</strong> Port<br />
Stanley specimen is largely intact, but the uropods<br />
and pleotelson posterior margin are heavily rubbed<br />
and the specimen is fragile. <strong>The</strong> descriptive details<br />
provided here were therefore obtained through direct<br />
examination.<br />
Menzies ( 962) placed Aega punctulata into synonymy<br />
with Aega semicarinata without explanation.<br />
Earlier, Hale ( 937) had clearly considered the species<br />
to be valid. <strong>The</strong>re are substantial differences between<br />
A. punctulata and A. semicarinata and indeed all other<br />
species <strong>of</strong> Aega, most particularly the prominently<br />
setose dorsal body surfaces, but also the shape <strong>of</strong> the<br />
pleotelson which in A. semicarinata is medially excavate,<br />
differences in the setation <strong>of</strong> pereopods –3 and<br />
in A. semicarinata much larger eyes. Aega urotoma is<br />
similar, but again lacks the setose body surfaces, has a<br />
subtruncate posterior margin to the pleotelson, which<br />
also lacks robust setae, has more strongly expanded<br />
antennule peduncle and antenna peduncle articles 4<br />
and 5, and has short robust setae on the merus <strong>of</strong> pereopods<br />
–3 in comparison to A. punctulata, these robust<br />
setae being arranged in a different pattern.<br />
Examination <strong>of</strong> the syntypes <strong>of</strong> Aega edwardsii<br />
Dollfus, 1891 allows confirmation that the species is<br />
a junior synonym <strong>of</strong> Aega punctulata. Although most<br />
dorsal setae are missing, enough setae remain and the<br />
presence <strong>of</strong> numerous setal sockets indicate that these<br />
specimens bear the unique setosity <strong>of</strong> A. punctulata.<br />
<strong>The</strong> synonymy is further confirmed by the eye size,<br />
frontal lamina shape, pleotelson, pereopod and uropod<br />
morphology and setation.<br />
Hale’s ( 937) record <strong>of</strong> this species from Maria<br />
Island, <strong>of</strong>f Tasmania, is a misidentification. Hale specifically<br />
mentions that his specimens lack the setose<br />
body surfaces described by Miers ( 88 ) as well as<br />
having more strongly dilated antennule and antenna,<br />
and illustrated his material as having larger eyes, a<br />
clearly rounded and crenulated posterior margin to the<br />
pleotelson and a more elongate frontal lamina. In A.<br />
punctulata antenna peduncle article 5 is shorter than article<br />
4 while in Hale’s figure it is longer. <strong>The</strong> identity <strong>of</strong><br />
a Hale’s record remains uncertain at present, although
Figure 141. Aega punctulata Miers, 88 . Holotype, except E and H (BMNH Port Stanley specimen). A, dorsal view;<br />
B, lateral view; C, head; D, frons; E, frontal lamina; F, pleotelson and left uropod; G, pleonites; H, pereopods (right) and<br />
2 (left), in situ.<br />
his figures show a species similar to Aega semicarinata<br />
and Aega urotoma.<br />
<strong>The</strong> record <strong>of</strong> Aega cf. punctulata from Madagascar<br />
by Barnard ( 960) is <strong>of</strong> equally uncertain identity. Bar-<br />
236<br />
nard was aware that the shape <strong>of</strong> the frontal lamina<br />
appeared to be unique, commenting that it was ‘almost<br />
sufficient to justify the institution <strong>of</strong> a separate species’,<br />
but desisted in the absence <strong>of</strong> comparative material. At
present this record can only be considered as a generic<br />
record <strong>of</strong> an undescribed species.<br />
Kussakin and Vasina ( 980) recorded Aega edwardsii<br />
from the Kerguelen Islands. <strong>The</strong> descriptive information<br />
provided is <strong>of</strong> family or generic level only and is<br />
inadequate to confirm or reject their identification. Setose<br />
body surfaces are not mentioned, and pereopod<br />
is figured as lacking robust setae which is in contrast to<br />
Aega punctulata, which has robust setae on the inferior<br />
margin <strong>of</strong> the merus.<br />
White’s ( 847) name is included in the synonymy,<br />
although the real identity <strong>of</strong> this nomen nudum can-<br />
237<br />
not be established. <strong>The</strong>re is nothing to indicate that<br />
this is the same species that was described by Dollfus<br />
( 89 ).<br />
distribution: Known from the Straits <strong>of</strong> Magellan, Falkland<br />
Islands and <strong>of</strong>f Cape Horn, South America.<br />
Aegiochus crozetensis (Kussakin & Vasina, 982),<br />
comb. nov. (Fig. 42)<br />
Aega crozetensis Kussakin & Vasina, 1982: 264, figs 5, 6.–<br />
Kensley, 200 : 226.<br />
Figure 142. Aegiochus crozetensis (Kussakin & Vasina, 982). Holotype. A, lateral view; B, head; C, frons; D, pereopods –3<br />
(from right to left); E, uropod endopod, ventral view; F, uropod; G. posterior margin <strong>of</strong> pleotelson.
materiaL examined: Holotype, ♂ ( 8.5 mm), Crozet<br />
Island, southern Indian Ocean, 46°36.2’S, 50°40. ’E,<br />
29 November 970, 280 m, coll. Skif III. (ZIASL RAN<br />
/7 626) [Specimen damaged, pereonite 3 crushed;<br />
dissected P , left uropod, pleopods and 2 not with<br />
specimen].<br />
desCriptiVe notes: Eyes separated by 6% width <strong>of</strong> head.<br />
Penial processes opening flush with ventral surface <strong>of</strong><br />
sternite 7, separated by ~ 0% width <strong>of</strong> sternite. Frontal<br />
lamina anterior margin with median point, posterior<br />
margin rounded, not ventrally directed, not blade-like.<br />
Posterior margin <strong>of</strong> pleotelson weakly serrate at the<br />
points <strong>of</strong> insertion <strong>of</strong> RS, with 2 (6+6) RS. Uropod<br />
endopod mesial margin weakly serrate, with 8 RS,<br />
lateral with 0+ (or 0+2 RS, missing RS not clearly distinguishable);<br />
uropod exopod mesial with 8 RS, lateral<br />
margin with ~ 2 RS. Pereopodal robust setae: pereopod<br />
propodus without small, distal RS; carpus with 0<br />
RS; merus with 0+2 or +2 RS; pereopod 2 propodus<br />
with small distal RS; carpus with 2 distal RS, merus<br />
with 2+2 RS; pereopod 3 propodus with 2 distal RS;<br />
carpus with 2 distal RS, merus with 2+2 RS.<br />
remarks: Permission to dissect was not granted, and<br />
the specimen, which had been previously dissected,<br />
was not accompanied by the dissected appendages.<br />
Aegiochus crozetensis can be identified by narrowly<br />
separated eyes, shape <strong>of</strong> the frontal laminar with an<br />
anterior median point and wide and rounded posterior<br />
margin, pereopods –3 with small robust setae on the<br />
merus and a weakly curved dactylus, and the shape<br />
and setation <strong>of</strong> the uropodal rami and posterior margin<br />
<strong>of</strong> the pleotelson.<br />
Aegiochus kanohi sp. nov. is similar to A. crozetensis<br />
but that species has united eyes and the frontal lamina<br />
has a blade-like posterior margin.<br />
distribution: Known from the vicinity <strong>of</strong> Crozet Islands,<br />
southern Indian Ocean.<br />
Aegiochus plebeia (Hansen, 897), comb. nov.<br />
(Figs 43, 44)<br />
Aega plebeia Hansen, 1897: 105, pl. 2, figs 4a–d.– Richardson,<br />
904: 29.– Van Name, 924: 83.– Birstein, 973: 72.<br />
Aega magnoculis Richardson, 909: 80, fig. 7; 9 0: 7.–<br />
Nierstrasz, 93 : 8 .– Gurjanova, 936: 70, 259.–<br />
Kussakin, 1979: 247, fig 118.<br />
Aega plebeja.– Nierstrasz, 93 : 83.– Gurjanova, 936: 72.<br />
[lapsus].<br />
Aega (Ramphion) plebeia.– Brusca, 1983: 19, figs 1b, 10, 11.<br />
materiaL examined: Syntypes, ♂ (23 mm), ♀ (non-ovig.<br />
30, ovig. 35 mm), <strong>of</strong>f Cocos Island, <strong>of</strong>f Panama, East<br />
238<br />
Pacific, 05°43’N, 85°50’W, 26 February 89 , Albatross<br />
stn 3363, 788 m (USNM 20726 [<strong>The</strong> non-ovigerous<br />
female has had all right-side appendages from P to the<br />
uropod dissected and the left-side antenna, antennule,<br />
mouthparts and uropod; labelling indicates dissection<br />
by Brusca.]). ♀ (non-ovig., 7.5 mm), 8°23’S, 7 ° 3’W,<br />
<strong>of</strong>f Peru, 972, 00 m, coll. E. del Solar (USNM 89292<br />
[previously examined by Brusca 983]). ♀ (non-ovig<br />
9 mm, damaged, crushed), <strong>of</strong>f Arica, Chile, 8°40.5–<br />
32.2’S, 70°36.0–29.8’W, 7 May 972, 768–968 m, 25’<br />
otter trawl (LACM C29 6, Acc#BI72-5SIO Benthic<br />
Invertebrates, MV72-II-27).<br />
Additional material: <strong>New</strong> caledonia: Manca ( 3.0<br />
mm), Norfolk Ridge, 24° 9’S, 67°49’E, 2 September<br />
985, BIOCAL stn. CP62, 395– 4 0 (MNHN Is.5863). ♀<br />
( 8.0 mm, non-ovig.), Norfolk Ridge, 23°52’S, 67°58’E,<br />
3 September 985, BIOCAL stn. CP69, 220– 225 m<br />
(MNHN Is.5864).<br />
desCriptiVe notes: Eyes separated by 7% width if head.<br />
Penes low tubercles; penial openings separated by<br />
3% <strong>of</strong> sternal width. Coxae not acute and posteriorly<br />
produced, posterior margins straight (2–4) or convex<br />
(5–7). Frontal lamina anterior margin with weakly<br />
produced median point, posterior margin rounded,<br />
blade-like, not ventrally directed. Posterior margin <strong>of</strong><br />
pleotelson weakly serrate at the points <strong>of</strong> insertion <strong>of</strong><br />
RS, with 2 (6+6) RS. Uropod endopod mesial margin<br />
weakly serrate, with 7 or 8 RS, lateral with 0+3 (or +3)<br />
RS; uropod exopod mesial with 4 RS, lateral margin<br />
with ~ 2 RS. Pereopodal RS: pereopod propodus<br />
with large, distal RS; carpus with small RS; merus<br />
with small distal RS; pereopod 2 propodus with<br />
large distal RS; carpus with large curved RS, merus<br />
with +2 small RS.<br />
Variation: Robust setae: Pleotelson – 4 (as 7+7,<br />
5+6 and 6+8). Uropodal exopod lateral margin 2– 4,<br />
mesial margin 4 (twice) or 5 (four times); uropodal<br />
endopod lateral margin 0+3 (twice) or +3 (ovigerous<br />
female), mesial margin 7 or 8 (three each).<br />
remarks: Among those species which have a posteriorly<br />
folded rostrum and frontal lamina with a free<br />
posterior margin, Aegiochus plebeia can be identified<br />
by the large and close-set eyes, pereopods –3 with a<br />
weak propodal lobe which is provided with a prominent,<br />
conspicuous robust seta, and the relatively wide<br />
uropodal rami. Aegiochus piihuka sp. nov. is immediately<br />
distinguished by the prominent propodal lobe on<br />
pereopods –3. Aegiochus tara sp. nov. is more similar,<br />
but has strongly laterally expressed and posteriorly<br />
acute coxae that are conspicuous in dorsal view and all<br />
<strong>of</strong> which have the posterior margin concave, posterior<br />
pereopods that are more slender, uropodal rami that
Figure 143. Aegiochus plebeia (Hansen, 897). Female syntype, 30 mm. A, dorsal view; B, lateral view; C, head; D, frons;<br />
E, maxilliped; F, maxilliped palp, articles 3–5; G, maxilla apex; H, maxillule apex; I, pleotelson posterior margin; J, pleonites,<br />
lateral margin.<br />
are more slender and acute, and the pleotelson lateral<br />
margins are noticeably sinuate with a more strongly<br />
produced and acute apex.<br />
Two specimens from the Norfolk Ridge (<strong>New</strong><br />
Caledonia) are here identified as Aegiochus plebeia,<br />
239<br />
agreeing in all characters but two. <strong>The</strong> robust seta at<br />
the distal end <strong>of</strong> the propodal palm <strong>of</strong> pereopods –3<br />
is somewhat smaller than in the type material, and the<br />
uropodal endopod lateral margin has a robust seta pattern<br />
<strong>of</strong> +3. Such variation occasionally occurs in what
Figure 144. Aegiochus plebeia (Hansen, 897). Female syntype, 30 mm. A, right pereopod ; B, right pereopod propodus;<br />
C, pereopod 2, ischium–dactylus; D, uropod; E, uropod exopod, ventral view.<br />
are otherwise consistent characters, and at present I<br />
consider these differences to be regional variation. It<br />
would require a far larger series <strong>of</strong> specimens to determine<br />
if there are two populations <strong>of</strong> cryptic species.<br />
Aegiochus plebeia was revised by Brusca ( 983),<br />
who included Aega magnoculis as a junior synonym.<br />
<strong>The</strong> distribution <strong>of</strong> this species, based on existing<br />
identifications and records is somewhat disjunct, with<br />
several records from the tropical East Pacific, two from<br />
the Alaskan region, and four records from the northwestern<br />
and western Pacific from Indonesia to Japan<br />
(Brusca 983). <strong>The</strong> depth range is given as 688–2534<br />
metres (Brusca 983). In the light <strong>of</strong> the new species<br />
described in this publication, the characters given by<br />
Brusca ( 983) to distinguish A. plebeia no longer do so.<br />
Given that there are two other similar species in the<br />
240<br />
Pacific (A. piihuka sp. nov. and A. tara sp. nov.), that<br />
there are two similar species in the East Pacific (see below)<br />
and one in the Atlantic, and that the stated depth<br />
range is inconsistent with distribution patterns shown<br />
by most aegids, I would regard all determinations other<br />
than those made here and those from the tropical East<br />
Pacific as requiring confirmation.<br />
Three other species are similar in appearance to<br />
Aegiochus plebeia:<br />
Aegiochus ventrosa (M. Sars, 859)—also has pereopods<br />
–3 with a weak propodal lobe that is provided<br />
with a prominent, conspicuous robust seta (ZMUC<br />
specimens), although this has been inconsistently<br />
figured for the species (M. Sars 1897; Bovallius 1885,<br />
886; Kussakin 979). A. ventrosa has a wider frontal<br />
lamina, with subtruncate lateral margins and a straight
posterior margin (rather than convex), and more robust<br />
pereopods –3 than A. plebeia. Known distribution:<br />
northern Atlantic.<br />
Aegiochus francoisae (Wetzer, 990)—has acute<br />
points on all coxae and no lobe on the carpus and propodus,<br />
the penes are basally fused and the pleotelson<br />
has 4+4 robust setae. It may be that some records <strong>of</strong><br />
A. plebeia are misidentifications <strong>of</strong> this species. Known<br />
distribution: the Galapagos region.<br />
Aegiochus symmetrica (Richardson, 905b)—is very<br />
similar to A. plebeia, but has no propodal lobe at all on<br />
pereopods –3 (and is therefore also very similar to<br />
A. francoisae). <strong>The</strong> recorded depth for this species is<br />
far more shallow than for A. plebeia. Known distribution:<br />
northeastern Pacific region (southeastern Alaska,<br />
Vancouver Island).<br />
distribution: Accepted records from the East Pacific:<br />
Panama and Chile; here provisionally recorded from<br />
the Norfolk Ridge, south <strong>of</strong> <strong>New</strong> Caledonia; at depth<br />
<strong>of</strong> 768– 788 metres. Likely to occur within the <strong>New</strong><br />
<strong>Zealand</strong> chart area, the Norfolk ridge records being<br />
just on the edge <strong>of</strong> the distributional limit for this<br />
monograph.<br />
Aegiochus uschakovi (Kussakin, 967), comb. nov.<br />
(Fig. 45)<br />
Aega uschakovi Kussakin, 1967: 225, figs 1, 2.<br />
Aega (Rhamphion) uschakovi.– Brusca, 983: 2.<br />
Not Aega sp. (aff. uschakovi).– Nunomura, 2005: 70, fig. 5 [=<br />
Aega sp.].<br />
materiaL examined: Holotype, ♀ (ovig. 8.0 mm), Drake<br />
Passage, 55°45’S, 70° ’W, 6 June 958, Ob’ stn 45 ,<br />
95– 05 m, (ZIASL RAN No /464 5).<br />
desCriptiVe notes: Eyes separated by 7% width <strong>of</strong> head.<br />
Frontal lamina triangular, posterior margin free, not<br />
downwardly directed. Posterior margin <strong>of</strong> pleotelson<br />
serrate at the points <strong>of</strong> insertion <strong>of</strong> RS, with 8 (4+4) RS.<br />
Uropod rami with margins distinctly serrate, most conspicuously<br />
on distal half <strong>of</strong> uropodal exopod. Uropod<br />
endopod mesial margin distinctly serrate, with 7 (both<br />
rami) RS, lateral with 2+2 RS (both rami); uropod exopod<br />
mesial with 6 RS (both rami), lateral margin with<br />
~ 2 RS. Pereopodal RS: pereopod propodus without<br />
RS; carpus with 0 RS; merus with 0+ RS; pereopod 2<br />
propodus with small distal RS, none on palm; carpus<br />
with small RS, merus with +2 RS.<br />
remarks: Permission to further dissect the type specimen<br />
was not given, and as it was not accompanied by<br />
the previously dissected appendages, a redescription<br />
<strong>of</strong> this poorly known species is not possible. Pereopods<br />
–3 on the left side are all damaged or missing; on<br />
24<br />
the right side pereopods and 2 were not in position<br />
suitable for drawing and pereopod 3 was obscured<br />
under pereopods 4 and 5; without dissection it is not<br />
possible to provide more precise detail <strong>of</strong> the setation<br />
<strong>of</strong> the anterior pereopods.<br />
<strong>The</strong> tentative identification <strong>of</strong> A. uschakovi by Nunomura<br />
(2005) is probably not correct. <strong>The</strong> figures<br />
given by Nunomura show that, in his material, the<br />
eyes are more widely separated, the pleotelson is more<br />
triangular in shape and the uropod endopod far narrower<br />
than in the type material.<br />
Aegiochus uschakovi can be identified by the relatively<br />
narrow eyes, which are clearly separated, the<br />
triangular frontal lamina, and the pattern and number<br />
<strong>of</strong> robust setae on the margins <strong>of</strong> the pleotelson and<br />
uropodal rami.<br />
distribution: Known from Drake Passage, Tierra del<br />
Fuego.<br />
Syscenus intermedius Richardson, 9 0 (Fig. 46)<br />
Syscenus intermedius Richardson, 1910: 17, fig. 16.– Bruce,<br />
997: 4.<br />
materiaL examined: Holotype, ♂ (26 mm), China Sea,<br />
20°37’N, 5°43’E, 8 August 908, 380–380 m, US<br />
Bureau <strong>of</strong> Fisheries Albatross Philippine Expedition,<br />
907–9, stn 530 (USNM 4 009).<br />
Additional material: ♂ (23 mm), Indonesia, <strong>of</strong>f Tanimbar<br />
Islands, 08°42’S, 3 °53’E, 2 November 99 ,<br />
stn. CP69, 356–368 m, coll. Baruna Jaya 1 (MNHN<br />
Is.5884).<br />
desCriptiVe notes: Anterior margin <strong>of</strong> head subtruncate,<br />
with obscure median point; dorsum <strong>of</strong> pereon<br />
somewhat vaulted; coxae each posteriorly rounded;<br />
pleotelson posteriorly rounded, lateral margins evenly<br />
convex, not inflected; frontal lamina wide, diamondshaped;<br />
antennule flagellum extends to middle <strong>of</strong><br />
pereonite ; antennal flagellum extends to posterior <strong>of</strong><br />
pereonite 4. Pereopod lacking RS; anterodistal angle<br />
<strong>of</strong> merus with about 6 simple setae. Pereopods 5–7 notably<br />
elongate, inferior margin <strong>of</strong> merus and carpus <strong>of</strong><br />
pereopod 7 with 6 and 9 acute RS respectively; uropod<br />
exopod and endopod subequal in length.<br />
remarks: Syscenus intermedius can be identified by having<br />
a subtruncate anterior margin to the head, rounded<br />
coxae, a broadly rounded pleotelson, antennal flagellum<br />
extending to posterior <strong>of</strong> pereonite 4, few setae<br />
on the distal margins <strong>of</strong> the ischium to merus <strong>of</strong> the<br />
pereopods, and pereopods 6 and 7 elongate, with the<br />
inferior margins <strong>of</strong> the merus and carpus provided<br />
with 6 and 9 acute robust setae respectively.
Figure 145. Aegiochus uschakovi (Kussakin, 967). Holotype. A, dorsal view; B, lateral view; C, head; D, frons; E, pleonite;<br />
F, pleotelson; G, pleotelson posterior margin; H, uropod endopod, ventral view; I, uropod exopod, ventral view.<br />
Bruce ( 997a) and Kensley (2004) both stated incorrectly<br />
that the species was known from the Philippines,<br />
whereas station data indicate that it was in reality collected<br />
closer to Hong Kong.<br />
242<br />
G<br />
distribution: Known from the type locality, south <strong>of</strong><br />
Hong Kong, South China Sea, here recorded from the<br />
Banda Sea, Indonesia.<br />
I
H<br />
Figure 146. Syscenus intermedius Richardson, 1910. All figs holotype. A, dorsal view; B, lateral view; C, pereopod 1;<br />
D, pereopod 7; E, pleonites, lateral view; F, head, dorsal view; G, frons; H, pleotelson.<br />
243
AppENdIx 3. OthEr mAtErIAl ExAmINEd<br />
This appendix lists identified specimens examined or identified in the course <strong>of</strong> preparing this monograph but<br />
not otherwise reported or cited in the text.<br />
1. species coded directly into AegA data set<br />
Aega antennata Richardson, 9 0: ♀ (non-ovig. 4 mm),<br />
eastern Indian Ocean, 29 m NW <strong>of</strong> Port Hedland,<br />
Western Australia, 8° 0’S, 8° 8’E, 0 October<br />
982, 298–300 m, coll. LMM on RV Soela (WAM<br />
2278-86). ♀ (non-ovig. 48 mm), eastern Indian<br />
Ocean, 29 m NW <strong>of</strong> Port Hedland, Western<br />
Australia, 8°26’S, 7°34’E, April 982, 4 8 m,<br />
coll. LMM on RV Soela (WAM 2275-86).<br />
Aega psora (Linnaeus, 758): ♂ (2 mm), ♀ (non-ovig.<br />
3 .5 mm), Disko, Godhavn, Greenland, December<br />
908, <strong>of</strong>f Somniosus microcephalus (ZMUC<br />
unreg).<br />
Aega serripes Milne Edwards, 840: ♀ (34 mm), Australia,<br />
64 km west <strong>of</strong> Kingston South Australia,<br />
36°50’S, 39°05’E, 6 Aug 909, FIS Endeavour<br />
(AM P43984). ♂ ( 7.5 mm), ♀ (ovig. 39, non-ovig.<br />
30 mm), d’Entrecasteaux Channel, Tasmania,<br />
October 929, abt. 5 fathoms, presented Mel<br />
Ward (AM P 0682). ♀ (non-ovig. 36 mm), Bottle<br />
and Glass, Port Jackson, Sydney, 20 January<br />
934, Iredale and Whitley (AM P37508). ?♀ ( 7.0<br />
mm), Shellharbour, NSW, 29 April 926, coll. Mel<br />
Ward (AM P45438). ♀ (non-ovig. 38 mm), east <strong>of</strong><br />
Wilsons Promontory, Victoria, 24 August 994,<br />
30 m, stn SS05/94/30 (AM P43965).<br />
Aegiochus arctica (Lütken, 859): ♂ (24 mm), ‘Greenland<br />
Island and Finmark’, no other data; specimen<br />
from H.M. Hale’s collections (AM P37520).<br />
Aegiochus maxima (Hansen, 897): holotype, ♀ (nonovig.<br />
54 mm), <strong>of</strong>f Cocos Island, <strong>of</strong>f Panama, 26<br />
February 89 , Albatross stn 3362, 2350 m [as 25<br />
fms] (USNM 20727).<br />
Aegiochus ventrosa M. Sars, 859: 3♀ (non-ovig. 8.5,<br />
20, 33 mm), Ingolf stn 95, 75 fvn, RT .70 (ZMUC<br />
unreg). ♂ (2 mm), 64°2 .5’N, 57°0 .5’W, Davis<br />
Strait, 2 December 992, 787–772 m, coll. Shinwa<br />
Maru (ZMUC unreg).<br />
Epulaega lethrina (Bruce, 983): ♂ (7.5 mm), north<br />
<strong>of</strong> Lord Howe Island, May 979, 23°58.00’S<br />
59°3.99’E, stn I738 (NIWA 34828).<br />
Epulaega nodosa Schioedte & Meinert, 879: ♂ ( 6.5<br />
mm), Bass Strait, 27 August 994, sled, 20 m, stn<br />
SS05/94/54 (AM P43966).<br />
244<br />
2. species examined or consulted in course <strong>of</strong><br />
monograph preparation<br />
Alitropus typus H. Milne Edwards, 840: ♀ (non-ovig.<br />
3.5 mm), Dawson River, tributary <strong>of</strong> Fitzroy<br />
River, Queensland, June 959, from giant perch<br />
[Lates calcarifer], coll. C. Vallis (AM P375 6). ♀<br />
(non-ovig. 3.5 mm), Northern Territory, Australia,<br />
#36 TMB0 - 7 (NTM unreg); 2♂ ( 2.0, 9.5<br />
mm), ♀ (non-ovig. 3.5 mm) 3 juveniles, Northern<br />
Territory, Australia, #36 TMB0 - 8 (NTM<br />
unreg).<br />
Aega australis Whitelegge, 90 . Syntypes; ♂ ( 2.5 mm;<br />
head detached, previously heavily dissected), ♀<br />
(ovig. . mm; previously dissected), 8.0–9.5 km<br />
<strong>of</strong>f Coogee, NSW, 33°57.0’S, 5 °2 .5’E, stn 44,<br />
91 m, fine sand, coll. E.R. Waite on HMCS <strong>The</strong>tis<br />
(AM G228 ). Non-type: ♀? (non-ovig. 9.5 mm),<br />
.0– 2.5 km <strong>of</strong>f Wollongong, NSW, 34°27’S,<br />
5 °04’E, stn 48, 02 m, sand, mud and rock, coll.<br />
E.R. Waite on HMCS <strong>The</strong>tis (AM P9604). (Note:<br />
only two [?types] specimens not four as stated<br />
on label.)<br />
Aega concinna Hale, 940. Holotype; ♂ (33 mm), entrance<br />
to Oyster Bay, Tasmania, 30 July 909,<br />
42°40’S, 48°03’E (AM E6740).<br />
Aega aff. falcata: entrance <strong>of</strong> Otsuchi Bay, Japan, 39°2’N,<br />
42°0’E, 9 April 995, <strong>of</strong>f Lophiomus setigerus<br />
(Vahl) (ZMUC unreg).<br />
Aega aff. monophthalma: ♀ (non-ovig. 50 mm), east<br />
<strong>of</strong> Heron Island, QLD, Australia, 2 ° 8.9’S,<br />
53°3 .7’E, 20 Nov 985, 502 m, coll. CSIRO Soela<br />
Cruise 0685, stn 8, AJB (NTM unregistered).<br />
Aega aff. semicarinata: ♂ (2 mm), South Africa, 6 km<br />
south <strong>of</strong> Cape Barracouta 3 December 929, 4<br />
m s Cap, 68 m, stn 50, coll. Dr. Th. Mortensen’s<br />
Java–South Africa Expedition (ZMUC unreg).<br />
Aega aff. semicarinata: ♀ (ovig. 53 mm), southeastern<br />
Atlantic, South Africa, 34°2 ’S, 7°57’E, 8 December<br />
929, 320 m, mud, coll. Dr. Th. Mortensen’s<br />
Java–South Africa Expedition (ZMUC unreg).<br />
Similar to A. semicarinata but lacking the large<br />
propodal RS, covered with short blunt glassy<br />
‘setae’ and wider body shape; the RS on anterior<br />
pereopods seem larger. Identity uncertain.<br />
Aegiochus aff. gracilipes Hansen, 895: ♀ (ovig. 24<br />
mm), Gulf <strong>of</strong> Guinea, Victoria–Banana, 02°00’N,<br />
09° 4’E, 2 December 950, 560 m, Galathea stn<br />
24 (ZMUC 2804).
Aegiochus aff. tara: ♀ (ovig. 27 mm), Western Indian<br />
Ocean, <strong>of</strong>f Kenya, 04°00’S, 4 °27’E, 5 March 95 ,<br />
55 m, Galathea stn 63 (ZMUC 2803).<br />
Aegiochus sp. ♀ (ovig. .5 mm), Japan, Okinawa, Sagami<br />
Sea, 2 July 9 4, 400 fv, coll. Th. Mortensen<br />
(ZMUC unreg). Not any species that I can recognise;<br />
eyes narrowly separate, no pleotelson RS,<br />
pleopods not digitate, but otherwise similar to<br />
A. coroo.<br />
Aegiochus sp. ♀ (ovig. .5 mm), Canada, British Columbia,<br />
‘W <strong>of</strong> Snake Island, between Snake Island<br />
and Gabriola Island, Straits <strong>of</strong> Georgia’, 4 June<br />
9 5, c. 63 m, from sponge, coll. Th. Mortensen<br />
(ZMUC unreg). Not any species I that I can recognise;<br />
eyes small, similar to A. laevis, but pleotelson<br />
RS, pleopods not digitate.<br />
Aegiochus sp. ♂ ( .5 mm), Indonesia, Java, 07°35’N,<br />
4°42’E, 0 April 929, 200 m, coll. Th. Mortensen<br />
(ZMUC unreg). Not any species that I can recognise;<br />
4 pleotelson RS, pleopods not digitate, no<br />
RS on propodal palm <strong>of</strong> P –P3, otherwise similar<br />
to A. laevis.<br />
Aegiochus sp. (nov.): ♂ (9.5 mm), 3 ♀ (non- ovig 2.0,<br />
ovig 9.3, 3.5 mm), Indonesia, Kei Islands, 2 May<br />
922, stn 59, 385 m, coral (ZMUC unreg). Not any<br />
species I can recognise; vaguely similar to A. coroo<br />
but pleopods not digitate, no pleotelson RS, males<br />
and non-ovig. with long RS P –P3, these being<br />
shorter in ovig female.<br />
Aegiochus vigilans (Haswell, 88 ): , Indonesia, Kei<br />
Islands, 2 April 922, stn 6, sand, Lithothamnion,<br />
Danske Exped. til Kei Islands, 922 (ZMUC unreg).<br />
, Indonesia, Kei Islands, 24 April 922, stn<br />
39, 60 m, sand, Lithothamnion, Danske Exped. til<br />
Kei Islands, 922 (ZMUC unreg).<br />
245<br />
Barybrotes species. At present it is accepted that the<br />
genus is monotypic, the single species being<br />
B. indus Schioedte and Meinert, 879a. No attempt<br />
is made here to reassess the identity or<br />
validity <strong>of</strong> other proposed names. ♀ (ovig. 2<br />
mm), Cauda, Nha-Trang, 5 May 929, Dana stn<br />
37 0 (ZMUC unreg); ♀ (ovig. with ova, 2 mm),<br />
5th Thai–Danish Expedition, 966, stn, 022, haul<br />
6, 4 – 966 m (ZMUC unreg); ♀ (non-ovig.,<br />
6 mm), 5th Thai–Danish Expedition, 966, stn.<br />
025, haul 4, 8 – 966 m [7th leg present, no<br />
app. M.] (ZMUC unreg); manca ( .5 mm), 5th<br />
Thai–Danish Expedition, 966, stn, 022, haul<br />
9, 4 – 966 m [no 7th leg] (ZMUC unreg); ?( 5<br />
mm, may be male), 4°4 ’N, 98° 3’E, 9 Nov 929,<br />
Dana stn 3900 [pleopods too fragile to examine]<br />
(ZMUC unreg); ♀ (non-ovig., 7.5 mm), 4°20’N,<br />
98°47’E, 0 Nov 929, Dana stn 390 [7th leg<br />
present, no app. M.] (ZMUC unreg); manca (9.0<br />
mm), 4°4 ’N, 98° 3’E, 9 Nov 929, Dana stn 3900<br />
(ZMUC unreg); manca (7.2 mm), <strong>of</strong>f Mombasa,<br />
Kenya, 22 Mar 95 Galathea stn. 259 [telson damaged]<br />
(ZMUC unreg).<br />
Rocinela ‘orientalis’: , Indonesia, Kei Islands, 9 May<br />
922, stn 53, 85 m, sand, coral, Danske Exped. til<br />
Kei Islands, 922 (ZMUC unreg).
1. matrix Aega species<br />
Polymorphic states: x = /2; y = 2/3; z = 2/4.<br />
AppENdIx 4. mAtrIcEs<br />
'Tridentella' 1111111111 1x11111111 1111111111 1??1??1111 x1111x1111 1111111111 1111111111 11x11<br />
alazon 1122131213 1211143211 2211232121 1412223211 1222111212 2413312111 2122211131 12313<br />
angustata 1122121133 2311421412 21?22222?? ????????22 1211212213 11133????1 2?31213111 11131<br />
antarctica 1x32211211 2212324111 1111142112 2321112111 211122111? 1313521221 2221313212 21212<br />
antennata 2122221223 1223131222 2311222121 1312123211 22212112?? 23322????1 2131113111 21122<br />
antillensis 1222131211 2213144211 2111232111 1312213211 1222111212 2433212311 2122211132 22313<br />
arctica 1132222123 1211353111 1111142312 2311112112 2111221112 1312111111 2121213112 21212<br />
banda 1222121x13 1211144211 12?11321?1 1??222?211 1222111212 2431312111 2122211131 22313<br />
beri x232222111 2211355111 11111331?2 2311112111 2121221113 1212112111 2121113112 21212<br />
bertrandi 1212231311 2211321111 1111133112 2321112111 2121221113 1211221212 2221113112 21211<br />
birubi 2222121113 1211143211 2121232121 1322233211 1222111212 2432112111 2122211131 22313<br />
copidis x222132311 2213143211 2?112321?? 1???211211 1224111211 2231412111 2122211131 22312<br />
coroo 1112231311 2211321111 1111133112 2321112112 2121211113 1311211212 2221113112 11211<br />
derkoma 1211131122 221112?111 1111132112 2231????11 12212111?? 13111????1 2221113112 11111<br />
deshaysiana 1222121213 1213143211 2211232121 1311?21211 1222111212 2433512111 2122211132 22311<br />
excise 1122121113 1211143211 2321222111 1312223211 12211112?? 24332????1 2122211131 12312<br />
falcate 2122221121 22211y1222 1121222121 1412113221 22212111?? 13311????1 11?11?2112 2??12<br />
falklandica x222122413 1213123312 2121222211 1522213211 1222211111 2231312211 2131111121 31211<br />
fracta 2111131123 2211122111 1111123112 2321124111 1221211131 1331122111 2231113112 21221<br />
francoisae 1232222111 1211323111 1111143?13 ?32?112111 3121?21131 1311211211 ?221111?11 11211<br />
glacialis 1232221123 2213324111 111?142312 2321112111 2113221121 2312121221 2221213212 31212<br />
gordoni 12322212x2 2211321111 1111142312 2221112111 2111221112 2313111212 1221313211 21112<br />
hamiota 2222132123 1211145211 2321232121 1312233211 1221111211 2431212111 2132212131 12313<br />
insomnis x232221z12 2211321111 1111223112 2221112111 2111211121 2312521212 1221113212 21111<br />
japonica x122131213 x211143211 2111232221 1422223x11 12221112?? 24332????1 2122211131 22312<br />
kakai 1232232313 1211323111 1111142312 2321112111 2111221122 2313121221 2221313212 31213<br />
kanohi 1232231212 2211323111 1111142312 2431112112 2111221112 2312121211 2211313112 21111<br />
kixalles x222131123 1213145211 2111232121 1322233211 12241212?? 24333????1 2122211131 22313<br />
komai 1122122131 2311435412 21222222?1 12121?3222 1221222213 111?312111 ??3131311? ?2?31<br />
kwazulu x222131113 1212143211 23112321?1 1???2???11 1221111212 2412312111 2122211132 22311<br />
laevis 1232211311 1211322111 1111142312 22?1112112 1111221112 1113511222 2221113111 21211<br />
leptonica x132132311 1213354111 1111133113 22?1112212 21212211?? 13131????1 2121113212 21212<br />
lethrina 1211131121 2211121111 2111132112 2111114212 3221?11131 1311111111 2221113211 21111<br />
246
magnifica 1122112411 2213124211 1311122223 1322223211 1224111112 2331211111 2131133121 22211<br />
mahana x122131313 1213145211 2311222111 1212213211 1222121212 2433312111 2122211131 22313<br />
maxima 2222122121 2211123211 1311122221 1512123211 12221111?? 23335????1 213x11x121 22212<br />
monophthalma 3222132231 1121135412 2321221211 141x123211 1222211111 2332511211 2231132111 22222<br />
musorstom 2222132x23 1211141211 2111232121 1322223211 1224111212 2431212111 212221x121 12313<br />
nodosa 1211131121 1212122111 1111132112 2311114211 122122113? 1311122111 1221313112 22211<br />
nohinohi 1132222122 2211321111 1211142112 2221112111 2111221112 2312121212 2221313212 21112<br />
piihuka 2232222z11 1211324111 1111142313 2321112111 2113221111 2211221111 2221113212 21211<br />
plebeian 1232221121 2211324111 1111142313 2221112112 2113221111 1311221211 2221313112 21212<br />
psora 21221212x1 2212123312 2211231221 1412111211 1221111112 2231211211 1131111121 21212<br />
pushkini x232222221 2211323111 1111142312 2321112111 2111221111 1312121211 2221313112 21211<br />
rickbruscai 1222132223 1211141211 2311232121 1412223211 1222121212 2431212111 213221x121 12313<br />
riwha x132221121 2211354111 1111133112 2321112112 2121121111 1112111111 2221313211 31213<br />
semicarinata 2122122121 1232231312 2121222221 1322223211 1222212112 1332212111 2221232112 11213<br />
serripes 3122121131 1222231212 2211222211 1512223111 1221111121 2231212111 2131212112 21212<br />
sheni 1222121??1 1211122211 2321122211 1412213211 1224111112 2231111111 213133x121 21212<br />
stevelowei 2222131211 1211225312 1321121221 152x233211 1222111111 1331212211 2131123212 22221<br />
tara x2322223x1 2213124111 1111142312 2321112112 21112211?? 22122????1 2221313112 31212<br />
trulla 1222131213 2213145211 2121232121 1422123211 12141212?? 24314????1 212221x121 22312<br />
umpara 1222131123 2211143211 2111122111 1322223211 1222211212 2432212111 2122211131 12311<br />
urotoma 2122112121 1212231412 2322222221 1412123211 1221212112 13322????1 2231232112 21211<br />
ventrosa 1232222311 2221324111 1111142312 2211112111 2113221122 1313121221 2211313112 2x212<br />
vigilans 1112131121 1211324111 1111131113 2211111211 1221221113 1331111211 1121213112 22211<br />
warna x222132213 1211144211 2311232121 1312213211 12211112?? 24312????1 2122212131 22313<br />
whanui 3222111223 1231324312 1121122221 x???????12 12221111?? 22314????1 2131111111 32212<br />
247
2. matrix <strong>Aegidae</strong> genera<br />
Polymorphic states: x = /2; y = /3; z = 2/3.<br />
Tridentella 1111111111 111111x111 1111111111<br />
Aega 1122311121 1211122111 1221211211<br />
Aegapheles 1122311121 1211122111 1221211212<br />
Aegiochus 11221111x2 1111132121 1121211111<br />
Alitropus 2112321121 1111233122 2--2222322<br />
Epulaega 1111311121 1111132121 1321211111<br />
Rocinela 2122211121 212123322x 2--2222322<br />
Syscenus 12111222y1 2122123222 z--2122322<br />
Xenuraega 1211122231 2122--4222 3--2--21—<br />
248
Aega, 7, 0, 2, 3, 4, 5, 27, 98<br />
Aega acuminata, 32, 2 2<br />
Aega acuticauda, 2 2<br />
Aega angustata, 7, 4, 37, 2 2, 232, 235<br />
Aega antennata, 7, 4, 27, 2 2, 245<br />
Aega approximata, 2 2<br />
Aega australis, 23 , 245<br />
Aega bicarinata, 2 2, 2 3<br />
Aega bicavata, 49, 50<br />
Aega chelipous, 2 2<br />
Aega concinna, 50, 2 2, 245<br />
Aega crenulata, 2 2<br />
Aega cyclops, 6, 210, 2 , 2 3<br />
Aega d<strong>of</strong>leini, 6, 27, 2 2<br />
Aega ecarinata, 2 2<br />
Aega edwardsii, 23 , 235, 236<br />
Aega falcata, 4, 27, 2 2<br />
Aega falklandica, 28, 32, 55, 98<br />
Aega fracta, 5<br />
Aega giganteocula, 5<br />
Aega hirsuta, 2 2<br />
Aega komai, 4, 28, 34, 37, 98, 233<br />
Aega lecontii, 2 2<br />
Aega magnifica, 27, 32, 2 2, 2 9<br />
Aega magnoculis, 239<br />
Aega maxima, 4, 27, 32, 33, 2 2<br />
Aega megalops, 2 2<br />
Aega microphthalma, 2 2<br />
Aega monophthalma, 7, 28, 37, 98<br />
Aega nanhaiensis, 2 2<br />
Aega novizealandiae, 211, 23<br />
Aega platyantennata, 2 2<br />
Aega psora, 4, 26, 2 2, 245<br />
Aega punctulata, 49, 50, 55, 2 2, 235, 244<br />
Aega rosacea, 2 2<br />
Aega semicarinata, 28, 44, 49, 50, 60, 235, 236<br />
Aega serripes, 4, 2 2, 245<br />
Aega sheni, 27, 2 2<br />
Aega stevelowei, 28, 50, 55<br />
Aega stroemii, 50, 2 3<br />
Aega tridens, 2 3<br />
Aega truncata, 2 3<br />
Aega urotoma, 28, 49, 50, 55, 60, 99, 2 3, 235, 236<br />
Aega ventrosa, 83<br />
Aega vigilans, 2<br />
Aega webbii, 50, 60, 2 3<br />
Aega whanui, 28, 61<br />
Aega sp., 245<br />
INdEx<br />
Principal taxonomic account is in bold font. Synonymic entries and material examined are not included;<br />
<strong>New</strong> <strong>Zealand</strong>, North Island and South Island are not indexed<br />
249<br />
Aegapheles, 7, 2, 4, 5, 27, 65<br />
Aegapheles alazon, 7, 3, 66, 68, 79<br />
Aegapheles antillensis, 2 3<br />
Aegapheles banda, 2 3<br />
Aegapheles birubi, 66, 68<br />
Aegapheles copidis, 66, 70, 73<br />
Aegapheles deshaysiana, 3, 4, 2 3<br />
Aegapheles excisa, 2 3<br />
Aegapheles hamiota, 66, 73, 8<br />
Aegapheles japonica, 2 3<br />
Aegapheles kixalles, 73, 79, 2 3<br />
Aegapheles kwazulu, 2 3<br />
Aegapheles mahana, 66, 75, 79<br />
Aegapheles musorstom, 73, 2 3<br />
Aegapheles rickbruscai, 66, 79, 8<br />
Aegapheles trulla, 73, 2 3<br />
Aegapheles umpara, 5, 66, 81<br />
Aegapheles warna, 79, 2 3<br />
<strong>Aegidae</strong>, 3, 5, 22, 26<br />
Aegiochus, 6, 7, 0, 2, 83, 2<br />
Aegiochus antarctica, 6, 23 , 216, 2 8, 220<br />
Aegiochus arctica, 4, 2 3, 245<br />
Aegiochus australis, 2 3<br />
Aegiochus beri, 3, 85, 88, 44<br />
Aegiochus bertrandi, 3, 88, 92, 97, 23<br />
Aegiochus coroo, 3, 8, 92, 93, 03, 9, 22, 2 0<br />
Aegiochus crozetensis, 02, 03, 4, 40, 2 3, 237, 238<br />
Aegiochus cyclops. See Aega cyclops<br />
Aegiochus dentata, 2 3<br />
Aegiochus dollfusi, 2 3<br />
Aegiochus francoisae, 4, 2 3, 24<br />
Aegiochus glacialis, 219, 22<br />
Aegiochus gordoni, 97, 08, 22, 28, 40<br />
Aegiochus gracilipes, 2 3<br />
Aegiochus incisa, 2 3<br />
Aegiochus insomnis, 103, 22<br />
Aegiochus kakai, 3, 97, 109, 9, 40<br />
Aegiochus kanohi, 97, 114, 40, 5 , 238<br />
Aegiochus koltuni, 2 9<br />
Aegiochus laevis, 2, 03, 120, 23<br />
Aegiochus leptonica, 4, 2 3<br />
Aegiochus lethrina, 6, 4<br />
Aegiochus longicornis, 2 4<br />
Aegiochus maxima, 244<br />
Aegiochus nohinohi, 08, 22, 123, 28, 40, 5<br />
Aegiochus nordenskjoldii, 83<br />
Aegiochus perulis, 28, 2 4<br />
Aegiochus piihuka, 6, 128, 34, 40, 48, 238
Aegiochus plebeia, 4, 6, 84, 34, 48, 2 4, 238, 239<br />
Aegiochus pushkini, 02, 03, 134, 40, 5<br />
Aegiochus quadratisinus, 44, 2 4<br />
Aegiochus riwha, 2, 3, 88, 141, 44<br />
Aegiochus spongiophila, 6, 2 4<br />
Aegiochus symmetrica, 34, 2 4, 24<br />
Aegiochus synopthalma, 9, 2 4<br />
Aegiochus tara, 146, 48, 238,240<br />
Aegiochus tenuipes, 2 4<br />
Aegiochus tiaho, 2<br />
Aegiochus tumida, 2 4<br />
Aegiochus uschakovi, 02, 03, 4, 40, 2 4, 241<br />
Aegiochus ventrosa, 6, 3, 4, 83, 34, 48, 2 4, 240, 245<br />
Aegiochus vigilans, 2, 3, 150, 52, 23 , 246<br />
Aegiochus webberi, 2<br />
Aegiochus weberi, 233<br />
Aegiochus sp., 5 , 245, 246<br />
Alaska, 89, 2 3, 2 4, 2 5, 240, 24<br />
Alitropus, 7, 26<br />
Alitropus typus, 2 4, 245<br />
Antarctic butterfish, see Hyperoglyphe antarctica<br />
Antarctica, 2 6<br />
Antipodes Islands, 93<br />
Anuropidae, 4<br />
Arafura Sea, 97<br />
Arctic, 89<br />
Argentina, 02<br />
ascidian (sea squirts), 6, 2 3<br />
Atlantic, 7, 2 3, 2 4, 2 5<br />
North Atlantic, 6, 7, 44, 34, 49, 98, 99, 206, 2 2,<br />
2 3, 2 4, 2 5<br />
South Atlantic, 33, 40<br />
Australia, 6, 7, 26, 36, 37, 44, 52, 55, 68, 70, 83, 88, 93,<br />
97, 9, 23, 34, 50, 5 , 57, 62, 2 0, 2 , 2 2, 2 3,<br />
2 4, 233, 235. See also individual States<br />
Azores, 2 2<br />
Bahamas, 44, 2 2<br />
Banda Sea, 206, 2 3, 2 4, 245<br />
barracouta, see Thyrsites atun<br />
barramundi, see Lates calcarifer<br />
Barybrotes indus, 25, 246<br />
Barybrotidae, 25<br />
Bass Strait, 235<br />
bathypelagic, 7<br />
Bay <strong>of</strong> Islands, 220<br />
black-eared spurdog, see Squalus melanurus<br />
bluenose, see Hyperoglyphe antarctica<br />
Bounty Trough, 63, 69<br />
Bouvet Island, 40<br />
Brazil, 2 5<br />
Britain, 2 3, 2 4, 2 5<br />
Brunnaega roeperi, 4<br />
California, 2 2, 2 3, 2 4<br />
Campbell Plateau, 93<br />
Cape Horn, 237<br />
Carangidae, 68<br />
250<br />
Carcharhinidae, 83<br />
Carcharhinus, 2 2<br />
Carcharhinus galapagensis, 83<br />
Caribbean, 77, 2 2, 2 5<br />
Celebes Sea, 2 4<br />
Centrolophidae, medusafishes, 44, 55, 161<br />
Centrophoridae, gulper sharks, 44<br />
Centrophorus squamosus, 44<br />
Challenger Plateau, 206<br />
Chatham Islands, 79<br />
Chatham Rise, 44, 03, 4, 9, 28, 84, 89, 97<br />
Chile, 50, 28, 2 3, 2 5, 24<br />
China, 2 2<br />
Cirolana, 4<br />
Cirolanidae, 7, 4, 25, 2<br />
Cirolanoidea, 4<br />
Cook Strait, 70, 73, 97<br />
Coral Sea, 88, 2 2, 2 3, 2 4<br />
Corallanidae, 4, 25, 2<br />
coral-reef, 6, 52<br />
Costa Rica, 2 2, 2 5<br />
Crozet Islands, 50, 03, 2 3, 238<br />
cryptic species, 3, 67, 238<br />
Cuba, 2 3, 2 4, 2 5<br />
Cymothoida, 6, 25<br />
Cymothoidae, 4, 22<br />
Cymothooidea, 4, 22<br />
Dalatiidae, sleeper sharks, 44<br />
Denmark, 43<br />
dogfish, see Squalidae<br />
Drake Passage, 03, 2 4, 24 . See also Tierra del Fuego<br />
Ecuador, 2 5<br />
endemism, 7<br />
England, see Britain<br />
Epicaridea, 4<br />
Epulaega, 7, 0, 151, 2 0<br />
Epulaega derkoma, 2, 5 , 152, 55<br />
Epulaega fracta, 6, 5, 52, 156, 6<br />
Epulaega lethrina, 5 , 52, 2 4, 245<br />
Epulaega monilis, 5 , 6 , 69, 2 4<br />
Epulaega nodosa, 4, 5 , 2 4, 245<br />
Euplectella, 2 4<br />
Færoe Islands, 44, 2 3<br />
Falkland Islands, 33, 50, 2 2, 238<br />
Florida, 2 3, 2 4<br />
Fossil <strong>Aegidae</strong>, 4<br />
France, 2 2<br />
Gadus morrhua, cod, 44<br />
Galapagos Islands, 2 2, 2 3, 2 4, 2 5, 24<br />
Galapagos shark, 9<br />
Gempylidae, snake mackerel, 67<br />
Genypterus blacodes, cusk eel, 67<br />
Georgia, USA, 2 5<br />
German Sea, 43<br />
Great Barrier Island, <strong>New</strong> <strong>Zealand</strong> 55, 6<br />
Great Barrier Reef, Australia 52, 6 , 2 4
Greenland, 2 2, 2 3, 2 5<br />
Greenland shark, see Somniosus microcephalus<br />
Gulf <strong>of</strong> Mexico, 2 2, 2 3<br />
Gulf <strong>of</strong> Suez, 2 3<br />
gulper shark, see Centrophorus squamosus<br />
Haploniscidae, 6<br />
hapuku, see Polyprion oxygeneios<br />
Hauraki Gulf, 37<br />
Hawai’i, 44, 89, 2 4, 2 5<br />
Hawke Bay, 9<br />
Hexactinellidae, glass sponges, 34, 23<br />
Hippoglossus, halibut, 2 5<br />
Hong Kong, 23 , 245<br />
Hyperoglyphe antarctica, 44, 55, 6<br />
Iceland, 43, 2 2, 2 3, 2 4<br />
India, 5 , 2<br />
Indian Ocean, 7, 25, 33, 49, 97, 02, 40, 2 4, 23 , 238<br />
Indo–Australasia, 7<br />
Indo-Malaysia, 6, 2 4<br />
Indonesia, 64, 77, 206, 2 2, 2 3, 2 4, 23 , 239<br />
Indo-Pacific, 152, 215<br />
Japan, 37, 206, 2 2, 2 4, 2 5, 239<br />
Juan Fernandez Islands, 50<br />
Kaikoura, 6<br />
Kerguelen Islands, 44, 50, 237<br />
Kermadec Islands, 6, 83, 2<br />
Lates calcarifer, 245<br />
Lethrinidae, rudderfishes, 152, 214<br />
Loligo bleekeri, 46<br />
longnose spurdog, see Squalidae<br />
Lophiidae, monkfishes, 212<br />
Lophius setigerus, sea devil fish, 212<br />
Lord Howe Island, 88<br />
Lord Howe Rise, 65, 83, 206<br />
Macquarie Island, 50<br />
Macquarie Ridge, 40, 89<br />
Madagascar, 2 2, 236<br />
Maine, 2 4<br />
Marion Island, 33, 50<br />
matiri, see Hyperoglyphe antarctica<br />
Mediterranean, 98, 2 2, 2 3, 2 4, 2 5<br />
mesopelagic, 6, 7, 98<br />
Metacirolana, 08<br />
Metacirolana caeca, 7, 26, 44<br />
Mexico, 89, 2 5<br />
micropredators, 6<br />
Mississippi, 2 5<br />
molecular analyses, 3<br />
Natatolana, 25<br />
<strong>New</strong> Caledonia, 6, 43, 60, 68, 75, 79, 8 , 92, 97, 9, 34,<br />
43, 45, 5 , 206, 2 2, 2 3, 239, 24<br />
<strong>New</strong> South Wales, 52, 33, 209, 2 0, 2 2, 2 5<br />
Norfolk Basin, 55<br />
Norfolk Island, 83<br />
Norfolk Ridge, 65, 73, 92, 45, 49, 202, 239, 24<br />
Norway, 442, 2 2, 2 3, 2 4<br />
25<br />
Ophidiidae, 68<br />
Otago, 97<br />
Pacific Ocean, 7<br />
East Pacific, 6, 7, 26, 28, 127, 133, 212, 214, 215, 231,<br />
238, 239<br />
North Pacific, 7<br />
southwestern Pacific, 6, 7, 13, 161, 198<br />
western Pacific, 7, 97, 134<br />
Palaega, 3<br />
Panama, 2 5, 240, 24<br />
Papua <strong>New</strong> Guinea, 6, 2 4<br />
parasites, 6<br />
pelagic, 7<br />
Peru, 99, 2 4, 2 5, 23<br />
Philippines, 5 , 2 , 2 3, 245<br />
Polyprionidae, wreckfishes, 50<br />
Polyprion oxygeneios, 68, 70<br />
Polyprion prognatus, 50<br />
Poor Knights Islands, 09<br />
Portugal, 2 2, 2 3<br />
Puerto Rico, 2 2<br />
Queensland, 75, 88, 23, 2 4, 2 5, 245<br />
Raja binoculata, big skate, 2 5<br />
Raja nasuta, rough skate, 60<br />
Rajidae, skates, 60<br />
Red Sea, 2 2, 2 3<br />
Rexea solandri, silver gemfish, 68<br />
Rhamphion, 2, 83, 84<br />
Rocinela, 6, 7, 0, 3, 4, 5, 8, 68, 161, 98<br />
Rocinela affinis, 84, 2 4<br />
Rocinela americana, 2 4<br />
Rocinela angustata, 6, 2 4<br />
Rocinela australis, 2 4<br />
Rocinela belliceps, 6, 2 4<br />
Rocinela bonita, 163, 67, 89<br />
Rocinela cornuta, 69, 89, 2 4<br />
Rocinela cubensis, 2 4<br />
Rocinela danmoniensis, 6 , 68, 2 4<br />
Rocinela dumerilii, 2 4<br />
Rocinela garricki, 169, 72, 73, 97<br />
Rocinela granulosa, 2 5<br />
Rocinela hawaiiensis, 69, 89, 2 5<br />
Rocinela insularis, 2 5<br />
Rocinela japonica, 2 5<br />
Rocinela juvenalis, 68, 2 5<br />
Rocinela kapala, 84, 2 5<br />
Rocinela laticauda, 2 5<br />
Rocinela leptopus, 2, 174, 77, 92, 93, 99<br />
Rocinela lukini, 2 5<br />
Rocinela maculata, 2 5<br />
Rocinela major, 69<br />
Rocinela media, 77, 2 5<br />
Rocinela modesta, 68, 2 5<br />
Rocinela murilloi, 2 5<br />
Rocinela niponia, 84, 2 5<br />
Rocinela oculata, 84, 2 5
Rocinela orientalis, 211, 2 5, 246<br />
Rocinela pakari, 178, 84<br />
Rocinela patriciae, 2 5<br />
Rocinela propodialis, 2 5<br />
Rocinela resima, 68, 184, 89<br />
Rocinela richardsonae, 2 5<br />
Rocinela runga, 77, 189, 92<br />
Rocinela satagia, 73, 193, 96, 98<br />
Rocinela signata, 7, 77, 2 5<br />
Rocinela sila, 2 5<br />
Rocinela tridens, 2 5<br />
Rocinela tropica, 2 5<br />
Rocinela tuberculosa, 2 5<br />
Rocinela wetzeri, 2 5<br />
Rocinela sp., 98<br />
Ross Sea, 2 4, 2 5, 2 8, 2 9, 220, 22 , 23<br />
rough skate, see Raja nasuta<br />
Russia, 2 5<br />
Scotland, 44, 2 4<br />
Scymnum microcephalum, 44<br />
Sea <strong>of</strong> Okhotsk, 2 5<br />
Serranidae, 68, 52, 2 4<br />
Seychelles, 68<br />
sibling species, 3<br />
Skagerak, Norway, 44<br />
Solomon Islands, 97, 2 2<br />
Somniosus microcephalus, sleeper shark, 44, 2 2, 2 3<br />
South Africa, 6, 7, 44, 50, 55, 60, 66, 68, 98, 2 2, 2 3,<br />
2 5, 234, 245, 246<br />
South Australia, 2 0<br />
South China Sea, 2 0, 239<br />
Southern Ocean, 9, 66, 207, 23<br />
Sphaeromatidae, 6, 7<br />
sponges, 6, 4, 34, 2 2, 2 3, 23<br />
Squalidae, 37, 60<br />
Squalus blainvillei, 37<br />
Squalus melanurus, 60<br />
Straits <strong>of</strong> Magellan, 50, 2 2, 2 4, 237<br />
Sulawesi, 2 5<br />
Symbioses, 6<br />
252<br />
Syscenus, 6, 7, 2, 3, 5, 7, 26, 198<br />
Syscenus atlanticus, 99, 2 5<br />
Syscenus infelix, 98, 99, 206, 209, 2 5<br />
Syscenus intermedius, 202, 2 5, 241<br />
Syscenus kapoo, 199, 20<br />
Syscenus karu, 98, 2 5<br />
Syscenus latus, 99, 202, 204, 206<br />
Syscenus moana, 99, 202, 206<br />
Syscenus pacificus, 98, 206<br />
Syscenus peruanus, 98, 2 5<br />
Syscenus springthorpei, 99, 208, 209, 2 0<br />
Syscenus sp., 2 2<br />
Taiwan, 37, 34, 45, 2 2<br />
Taranaki Bight, 37<br />
Tasman Sea, 7, 88, 50<br />
Tasmania, 70, 6 , 2 2, 2 3, 235<br />
Three Kings Island, 28<br />
Thyristes atun, 70<br />
Tierra del Fuego, 24<br />
Timor Sea, 65<br />
Tonga, 68<br />
Trachurus novaezelandiae, horse mackerel, 68<br />
Tridentellidae, 3, 2<br />
Triglidae, 68<br />
Tristan da Cunha, 68<br />
Vancouver, 24<br />
Vanuatu, 34, 23<br />
Victoria, Australia, 55, 2 2, 2 3, 235<br />
Washington State, 2 5<br />
Weddell Sea, 2 9, 22<br />
Western Australia, 2 2<br />
Xenuraega, 7, 2, 5, 26<br />
Xenuraega ptilocera, 2 5