Download PDF as one large file
Download PDF as one large file
Download PDF as one large file

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
JOURNAL OF CRUSTACEAN BIOLOGY, 2(1): 84-119, 1982<br />
REVISION OF THE GENERA DYNAMENELLA,<br />
ISCHYROMENE, DYNAMENOPSIS, AND CYMODOCELLA<br />
(CRUSTACEA: ISOPODA), INCLUDING A NEW GENUS<br />
AND FIVE NEW SPECIES OF EUBRANCHIATE<br />
SPHAEROMATIDS FROM QUEENSLAND WATERS<br />
K. Harrison and D. M. Holdich<br />
ABSTRACT<br />
Collections of isopod crustaceans from a wide variety of intertidal marine habitats in<br />
Queensland, Australia, contained five new species and a new genus of eubranchiate sphaeromatids.<br />
Descriptions are given for the different stages of their growth and some ecological<br />
considerations are discussed.<br />
The new genus is Paradella and the new species are: Paradella octaphymata, new genus,<br />
new species; Dynamenella liochroea, new species; D. ptychura, new species; D. trachydermata,<br />
new species; and Ischyromene polytyla, new species.<br />
Ischyromene Racovitza, Dynamenella Hansen, Cymodocella Pfeffer, and Dynamenopsis<br />
Baker are redefined and rediagnosed. The importance of founding species on descriptions<br />
of adult males is stressed, but this should be supplemented with descriptions of all other<br />
stages of the life cycle if possible. The form of the brood pouch of the ovigerous female and<br />
the form of the penes and appendix m<strong>as</strong>culina of the mature male should be included <strong>as</strong><br />
these characters are relatively consistent at the generic level.<br />
Until recently the marine isopods of Australia were poorly known, but information<br />
gained by surveys in the l<strong>as</strong>t decade demonstrates that isopods are an<br />
important comp<strong>one</strong>nt of the fauna of littoral and shallow sublittoral benthic habitats<br />
of the tropical and subtropical regions of Queensland at le<strong>as</strong>t. The re<strong>as</strong>ons<br />
for the paucity of isopod records are that many ecologists have tended to deal in<br />
detail only with the <strong>large</strong>r elements of the macrofauna (e.g., Endean et al., 1956a,<br />
b; Stephenson et al., 1958) and many species of isopods are small and live in<br />
cryptic habitats. Also until recently there w<strong>as</strong> a dearth of aids to the identification<br />
of isopods.<br />
A wide variety of marine habitats on the co<strong>as</strong>t and offshore islands of Queensland<br />
were examined and a <strong>large</strong> number of isopod species were found. Many new<br />
records of distribution and new species have been recorded—Serolidae (Holdich<br />
and Harrison, 1980a), Gnathiidae (Holdich and Harrison, 1980b), Cirolanidae<br />
(Holdich et al., 1981), and certain Sphaeromatidae (Holdich and Harrison,<br />
1980c, 1981a, b). The Infra-orders Asellota and Valvifera do not seem to be<br />
well represented, but the Anthuridea are common in the shallow sublittoral and<br />
are currently being studied.<br />
Many genera of Sphaeromatidae are not well defined. This is because workers<br />
often put a new species into a genus which is not appropriate for the characters<br />
found (e.g.. Hurley and Jansen, 1977; Bruce, 1980), and others describe new<br />
species and define new genera on subadult specimens or females which belong<br />
to already described species b<strong>as</strong>ed on males (e.g., Menzies and Glynn, 1968;<br />
Pires, 1980a). This is particularly so with the eubranchiate sphaeromatids included<br />
here.<br />
The only record of an eubranchiate sphaeromatid isopod from Queensland<br />
waters is that of Dynamene curalii Holdich and Harrison described from coral<br />
reefs (Holdich and Harrison, 1980c). However, other species belonging to genera<br />
of eubranchiate sphaeromatids have now been collected and identified. In the<br />
84
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 85<br />
current paper, six of these eubranchiates are considered together because they<br />
occupy similar habitats and resemble the 'Dynamenella' form of sphaeromatid.<br />
They do in fact, <strong>as</strong> will be shown, belong to three distinct genera. A detailed<br />
discussion of these genera, and two others with which they have been confused,<br />
is given below with the hope of clarifying their systematic positions. In future<br />
papers the rest of the eubranchiates and the hemibranchiate sphaeromatids collected<br />
from Queensland will be examined.<br />
METHODS<br />
A wide variety of littoral habitats were examined on the mainland co<strong>as</strong>t of Queensland, on two<br />
continental islands offshore (Magnetic Is. and Lizard Is.), and on a coral cay (Heron Is.). Detailed<br />
examination w<strong>as</strong> made of beach rock; rock crevices; under st<strong>one</strong>s on sand; in sand and mud; amongst<br />
algae, barnacles, rock oysters, mangrove roots, coral, sponges, bryozoan and <strong>as</strong>cidian colonies,<br />
serpulid tubes, and wood jetsam. When isopods were not obvious in the field, part of the substratum<br />
w<strong>as</strong> collected and examined under a dissecting microscope, whilst being gently fragmented in 2%<br />
formalin to agitate the infauna.<br />
Isopods from sublittoral collections made by the James Cook University Three Bays Survey, and<br />
collections from the museums listed below, were also examined for 'Dynamenella-like" sphaeromatids.<br />
In addition, collections from Hinchinbrook Is., North Stadbroke Is., and Goat Is. were kindly<br />
donated by N. L. Bruce.<br />
The following abbreviations are used here to indicate the institutions where the specimens examined<br />
are to be found: Queensland Museum (Q.M.); Australian Museum (A.M.); South Australian Museum<br />
(S. Aust. M.); Western Australian Museum (W.A.M.); South African Museum (S. Afr. M.); Canterbury<br />
Museum, New Zealand (CM.); New Zealand Oceanographic Institute (N.Z.O.I); British Museum<br />
(Natural History) (B.M.N.H.); United States National Museum (U.S.N.M.); Museum National<br />
d'Histoire Naturelle, Paris (M. d'H.N.); Naturhistoriska Riksmuseet, Stockholm (N.R.).<br />
Any material menti<strong>one</strong>d below but not designated a museum reference number h<strong>as</strong> been placed in<br />
a reference collection in the Department of Zoology, Nottingham University.<br />
SYSTEMATICS<br />
For the correct placement of the sphaeromatids from Australia, a thorough<br />
review of the literature on Ischyromene Racovitza and Dynamenella Hansen w<strong>as</strong><br />
necessary. Much taxonomic confusion w<strong>as</strong> uncovered and the review w<strong>as</strong> extended<br />
to include Dynamenopsis Baker and Cymodocella Pfeffer. Type and other<br />
identified material w<strong>as</strong> obtained from several museum collections so that the<br />
review w<strong>as</strong> b<strong>as</strong>ed on more than the literature and its many ill-defined species.<br />
Because the genus Ischyromene h<strong>as</strong> virtually been ignored in the p<strong>as</strong>t, many<br />
species belonging to this genus have been placed in the genera Dynamenella and<br />
Cymodocella (e.g., Hurley and Jansen, 1977; Sivertsen and Holthuis, 1980;<br />
Simes, 1981). These three genera and Dynamenopsis (which h<strong>as</strong> been confused<br />
with Dynamenella) have been redefined and the species redistributed on the b<strong>as</strong>is<br />
of the generic characters given. The results of this are outlined below.<br />
OrderIsopoda<br />
Family Sphaeromatidae<br />
Subfamily Dynameninae<br />
Ischyromene Racovitza, 1908<br />
Diagnosis.—Eubranchiate Sphaeromatidae with antennular peduncle article 1 not<br />
extended anteriorly <strong>as</strong> a plate. Both sexes with pereon and pleon lacking dorsal<br />
processes. Pleon with posterior margin bearing two sutures at each side. Both<br />
sexes with both rami of uropod lamellar; endopod greater than half length of<br />
exopod. Apex of pleotelson with either a simple, ventral groove or an enclosed,<br />
dorsally directed foramen. Pereopods stout; accessory unguis often markedly
86<br />
JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
bifid. Endopod of pleopod 1 triangular with internal half indurate. Exopod of<br />
pleopod 3 without an articulation. Sexual dimorphism not pronounced.<br />
Adult Male.—Pereonite 7 often longer than that of females, with posterior margin<br />
markedly bilobed. Penes short, separate to b<strong>as</strong>e with rounded apices. Appendix<br />
m<strong>as</strong>culina narrow, arising from internoproximal angle of endopod of pleopod 2<br />
and extending to, or beyond, apex of endopod.<br />
Ovigerous Female.—Mouthparts not metamorphosed. Brood pouch formed posteriorly<br />
<strong>as</strong> a pocket, opening anteriorly at the level of pereopod 4; anteriorly<br />
formed <strong>as</strong> three pairs of <strong>large</strong> oostegites arising from pereopods 2 to 4 and overlapping<br />
well in the midline. Brood not incubated in the brood pouch thus formed,<br />
but in invaginations of the ventral body wall.<br />
Type-species.—Ischyromene lacazei Racovitza, •-'VU.W T ^ t . b U , 1908.<br />
i ^ VJ^-J.<br />
Additional species:<br />
/. australis (Richardson, 1906),<br />
new combination<br />
/. australoides (Barnard, 1940),<br />
new combination<br />
/. barnardi (Menzies and Glynn,<br />
1968), new combination<br />
/. bicarinata Harrison, 1981<br />
/. brunnea (Vanhoffen, 1914),<br />
new combination<br />
/. codii (Nobili, 1906),<br />
new combination<br />
/. condita (Hurley and Jansen,<br />
1977), new combination<br />
/. cordiforaminalis (Chilton,<br />
1883), new combination<br />
/. eatoni (Miers, 1875),<br />
new combination<br />
/. hirsuta (Hurley and Jansen,<br />
1971), new combination<br />
/. huttoni (Thomson, 1879),<br />
new combination<br />
/. insulsa (Hurley and Jansen,<br />
1977), new combination<br />
/. macrocephala (Krauss, 1843),<br />
new combination<br />
/. menziesi (Sivertsen and Holthuis,<br />
1980), new combination<br />
/. mortenseni (Hurley and Jansen,<br />
1977), new combination<br />
/. ovalis (Barnard, 1914),<br />
new combination<br />
/. rubida (Baker, 1926),<br />
new combination<br />
/. sapmeri (Kensley, 1976),<br />
new combination<br />
Current name:<br />
Dynamenella australis<br />
D. australoides<br />
D. barnardi<br />
I. bicarinata<br />
D. brunnea<br />
D. codii<br />
D. condita<br />
D. cordiforaminalis<br />
Cymodocella eatoni<br />
D. hirsuta<br />
Cymodocella huttoni<br />
D. insulsa<br />
D. macrocephala<br />
D. menziesi<br />
D. mortenseni<br />
Cymodocella ovalis<br />
D. rubida<br />
Cymodocella sapmeri<br />
Recommendation<br />
b<strong>as</strong>ed on<br />
examination of:<br />
Specimens<br />
Types<br />
Literature<br />
Types<br />
Type<br />
Types<br />
Type<br />
Specimens<br />
Types<br />
Type<br />
Specimens<br />
Type<br />
Specimens<br />
Literature<br />
Literature<br />
Literature<br />
Type<br />
Literature
Additional species:<br />
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 87<br />
/. scabricula (Heller, 1865),<br />
new combination<br />
/. tuberculata (Menzies, 1962b)<br />
new combination<br />
Current name:<br />
D. scabricula<br />
D. tuberculata<br />
Recommendation<br />
b<strong>as</strong>ed on<br />
examination of:<br />
Specimens<br />
Types<br />
Remarks.—Species of the genus Ischyromene show a number of variable characters.<br />
Epistome shape varies between species and may be a useful distinguishing<br />
characteristic; species may be more or less flattened with laterally extended coxal<br />
plates; the pleotelsonic apex may have a simple groove or an enclosed foramen;<br />
the pereopodal accessory unguis may be bifid or simple (this character is perhaps<br />
size related, with the smaller species showing the bifid form more clearly than<br />
the <strong>large</strong>r species); and the uropodal rami are sometimes apically rounded (e.g.,<br />
/. lacazei) or they may have sinuous margins with the exopodal apex acute (e.g.,<br />
/. austrails).<br />
With the transfer of many species to this genus it can be seen that, far from<br />
being restricted to the Mediterranean <strong>as</strong> previous records suggest, Ischyromene<br />
is a predominantly temperate genus centred in the southern hemisphere. The<br />
western Mediterranean may well be the Old World northern distributional limit<br />
of this genus (Fig. 7A).<br />
Synonyms.—N<strong>one</strong>.<br />
Ischyromene polytyla, new species<br />
Material Examined.—Holotype adult male, 4.2 mm (Q.M.: W.8055) (+1 microslide).<br />
Type-locality.—Burleigh Heads, South E<strong>as</strong>t Queensland (28°S). From dead wood.<br />
Coll. N. Svennivig, 03.iv. 1979.<br />
Etymology.—The adjectival specific name polytyla is derived from the Greek<br />
poly + tylos, i.e., many + lump.<br />
Description of Adult Male (Fig. la-n).—Ischyromene with tergum of pereonite<br />
7 rugose (especially obvious in lateral view). Pleon with scattered, prominent<br />
tubercles. Pleotelson with main dome bearing small, round, prominent tubercles<br />
with an additional tubercle anterior to the b<strong>as</strong>e of each uropod. Apical ventral<br />
groove of pleotelson with lip thickened and smoothly rounded in dorsal view; lip<br />
margins parallel in posterior view and groove open ventrally. Ventral margins of<br />
pleotelson narrow, lacking outcurved ridges.<br />
Appendages: Antennule with peduncular article 1 longer than articles 2 and 3<br />
together; flagellum of 12 articles. Antenna with 20-articled flagellum extending to<br />
level of pereonite 5. Mouthparts not examined in detail; maxillipedal palp articles<br />
2 and 3 with pronounced setose lobes and article 4 with low setose lobe. Pereopods<br />
robust, incre<strong>as</strong>ing in length posteriorly; merus, carpus, and propodus with<br />
superior fringes and dense inferior pads of setae. All pereopods with bifid accessory<br />
unguis. Penes flattened, each four times <strong>as</strong> long <strong>as</strong> broad with semicircular<br />
tip. B<strong>as</strong>is of pleopod 1 bearing five internal coupling hooks; exopod subovate,<br />
slightly longer than triangular endopod; inner half of endopod (that region not<br />
overlapped by exopod) indurate; both rami with long, terminal, plumose setae.<br />
B<strong>as</strong>is of pleopod 2 bearing five internal coupling hooks; exopod subovate, half<br />
length of sub triangular endopod; both rami with terminal and external plumose
88<br />
JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Fig. 1. Ischyromene polytyla, new species. Adult male holotype (a) dorsal, (b) lateral, (c) pereopod<br />
1, (d) epistome and labrum, (e) transverse section (T.S.) through pleotelsonic margin at level of bar<br />
on 'f,' (f) ventral, pleotelson, (g) posterior, pleotelson, (h) pleopod 1, (i) pleopod 2, Q) appendix<br />
m<strong>as</strong>culina apex, (k) pereopod 7, (1) antennular peduncle, (m) antennal peduncle, (n) penes. Ischyromene<br />
lacazei Racovitza. Adult male (o) pleopod 3, (p) appendix m<strong>as</strong>culina apex, (q) pleopod 4, (r)<br />
pleopod 5, (s) maxiUiped. Scale line represents 1 mm.
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 89<br />
setae; endopod with appendix m<strong>as</strong>culina just longer than ramus, narrow, lying<br />
alongside longitudinal ridge on anterior surface of endopod, and curving away<br />
from midline of body, widening slightly distally to rounded apex bearing microtrichia.<br />
Pleopods 4 and 5 not examined in detail (Fig. lo-s are from type-species<br />
of genus). Uropod with endopod just longer and narrower than exopod, extending<br />
just beyond apex of pleotelson; both rami smoothly rounded apically.<br />
Colour of Specimen in Alcohol.—Cream, with dorsal surface, pleopods, ventral<br />
pereon, and b<strong>as</strong>es of pereopods with evenly distributed, small, black chromatophores.<br />
Female. —Unknown.<br />
Distribution and Habitat.—Known only from wood in the intertidal region of<br />
southe<strong>as</strong>tern Queensland. (Well-weathered and relatively permanent wood provides<br />
a very 'popular' settling place for peracarid crustaceans and a number of<br />
isopod species have been recorded from this habitat in Queensland.)<br />
Remarks.—As only <strong>one</strong> specimen of /. polytyla is known, the maxiUiped and<br />
pleopods 3 to 5 were not removed for illustration. These appendages do not,<br />
however, appear to differ in form from those of the type-species of the genus. As<br />
these appendages have never been illustrated for /. lacazei, they are shown here<br />
in Fig. lo-s. For comparative purposes the apex of the appendix m<strong>as</strong>culina of/.<br />
lacazei, with its three external, subapical teeth, is also shown. /. polytyla, new<br />
species, resembles /. lacazei in dorsal view, but /. polytyla can be distinguished<br />
by the smoothly rounded margin of the pleotelsonic notch and the form of the<br />
appendix m<strong>as</strong>culina.<br />
Synonym.—Clianella Bo<strong>one</strong>, 1923.<br />
Dynamenella Hansen, 1905<br />
Diagnosis.—Eubranchiate Sphaeromatidae with antennular peduncle article 1 not<br />
extended anteriorly <strong>as</strong> a plate. Both sexes with pereon and pleon lacking dorsal<br />
processes, and with both rami of uropod lamellar, the endopod being greater than<br />
half length of exopod. Pereopod 1 markedly more robust than other pereopods.<br />
All pereopods with simple, not bifid, accessory unguis. Exopod of pleopod 3<br />
with or without articulation. Sexual dimorphism obvious.<br />
Adult Male.—Penes long, tapering, fused at b<strong>as</strong>e. Appendix m<strong>as</strong>culina arising<br />
from internoproximal angle of endopod of pleopod 2 and extending to or beyond<br />
apex of endopod. Appendix m<strong>as</strong>culina usually broad proximally, tapering to an<br />
acute tip. Apex of pleotelson with dorsally directed foramen connected to apex<br />
by narrow slit. Ventral margins of pleotelson various; flattening posteriorly, or<br />
directed medially, or with narrow outcurved ridges which meet in posterior midline.<br />
Uropods broader than those of females and immature specimens.<br />
Ovigerous Female.—Mouthparts not metamorphosed. Apex of pleotelson various;<br />
bearing slight notch, simple groove, or foramen connected to apex by narrow<br />
slit. Uropods narrower than those of adult male. Brood pouch lacking oostegites,<br />
formed from two opposing ventral pockets covering entire ventral<br />
pereon and opening in midline between fourth pereopods.<br />
Type-species.—Dynamenella perforata (Moore, 1901).
90<br />
Additional species:<br />
D. elegans (Bo<strong>one</strong>, 1923)<br />
D. josephi Glynn, 1968b<br />
D. marginata (Bruce, 1980),<br />
new combination<br />
D. nipponica (Nishimura, 1969),<br />
new combination<br />
D. platura NobiH, 1906<br />
D. quil<strong>one</strong>nsis Filial, 1954<br />
D. savignyi (Edwards, 1840),<br />
new combination<br />
D. tropica Silva, 1960<br />
JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Current name:<br />
Dynamenella elegans<br />
Dynamenella josephi<br />
Cymodocella marginata<br />
Cymodocella nipponica<br />
Dynamenopsis platura<br />
Dynamenella<br />
quil<strong>one</strong>nsis<br />
Dynamenopsis dumerilii<br />
(Audouin, 1826)<br />
Dynamenella tropica<br />
and Dynamenella antonii<br />
Silva, 1960<br />
Recommendation<br />
b<strong>as</strong>ed on<br />
examination of:<br />
Literature<br />
Literature<br />
Literature<br />
Literature<br />
Types<br />
Literature<br />
Specimen<br />
Literature<br />
Literature<br />
Remarks.^Dynamenella tropica appears to be b<strong>as</strong>ed on subadult males and females<br />
of D. antonii (b<strong>as</strong>ed on adult male), but by page priority D. tropica is<br />
retained <strong>as</strong> the senior subjective synonym.<br />
Dynamenella elegans appears to be a species very close to, and possibly conspecific<br />
with, D. tropica (fide Menzies and Glynn, 1968: 59).<br />
Sphaeroma dumerilii Audouin, 1826, w<strong>as</strong> found to be a junior homonym of<br />
Sphaeroma dumerilii Leach, 1818, by Edwards (1840), who proposed the replacement<br />
name Sphaeroma savignyi for Audouin's species. When Monod (1933)<br />
transferred this species to the genus Dynamenopsis he used Audouin's name,<br />
calling the species Dynamenopsis dumerilii (Audouin). The International Code of<br />
Zoological Nomenclature (1961) is not clear on this matter, but following Blackwelder's<br />
interpretation (1967: 551, Fig. B), even though Audouin's name h<strong>as</strong> been<br />
transferred to a different genus, it still competes in homonymy with that of<br />
Leach. To avoid any potential nomenclatural confusion, the present authors are<br />
using Edwards' replacement name in transferring the species to the genus Dynamenella<br />
<strong>as</strong> Dynamenella savignyi (Edwards).<br />
The type locality of Dynamenella platura w<strong>as</strong> originally unknown but Nobili<br />
(1907: 423) stated that, <strong>as</strong> the specimens came from the collections of Dr. Seurat,<br />
they certainly came from the Tuamotu Islands. The label in the type bottle now<br />
reads "I. Gamlier" [sic'\ (part of the Tuamotu Archipelago in the mid South<br />
Pacific). Ischyromene codii, another species from Seurat's collections, w<strong>as</strong> collected<br />
from the Gambler Islands two years after D. platura. Unless separate<br />
collections were made in the same area, it seems possible that the types of D.<br />
platura were not collected in the Gambler Islands, but were from some other<br />
location. Monod (1934) h<strong>as</strong> reported D. platura from Indo-China.<br />
Of the other species currently placed in the genus Dynamenella: D. benedicti<br />
(Richardson, 1899) and D.fraudatrix Kussakin, 1962, are both in fact hemibranchiate<br />
sphaeromatids and their correct generic placement is unknown; D. sheareri<br />
(Hatch, 1947), D. conica Bo<strong>one</strong>, 1923, and D. glabra (Richardson, 1899) have<br />
been inadequately described and no opinion can be given <strong>as</strong> to their status; D.<br />
dioxus Barnard, 1914; D. navicula Barnard, 1914; D. parva Baker, 1929; and D.<br />
taurus Barnard, 1940, are all eubranchiates which cannot be placed in any of the<br />
five genera considered here and their correct placement is unknown at the present
HARMSON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 91<br />
time. D. angulata (Richardson, 1901) is b<strong>as</strong>ed on subadult males of a species of<br />
Paracerceis Hansen, since by the use of transmitted light the adult male pleotelsonic<br />
form of a Paracerceis can be seen within the pleotelsons of type specimens.<br />
The remaining eight species will be considered below (see Paradella, new<br />
genus, and Cymodocella).<br />
In his generic diagnosis of Dynamenella, Hansen (1905: 107) stated "Both<br />
sexes . . . without real processes." Barnard (1914: 410) modified this to "7th<br />
peraeon segment with or without processes in male." He did this in order to<br />
include his species Dynamenella dioxus in this genus and justified the emendation<br />
by saying that in Dynamenella perforata, D. australis (i.e., Ischyromene australis),<br />
and D. scabricula (i.e., /. scabricula) the seventh pereonite is extended<br />
backwards in two lobes. This lobing of the seventh pereonite cannot be equated<br />
with Hansen's "processes" (nor can the mediolateral ridging shown by species<br />
such <strong>as</strong> Dynamenella dianae (Menzies) (see below), which Pires (1980b) also referred<br />
to <strong>as</strong> "processes." Hansen w<strong>as</strong> referring to the unmistakable extensions<br />
shown by species of such genera <strong>as</strong> Dynamene Leach, Isocladus Miers, Zuzara<br />
Leach, Cilicaea Leach, and others. N<strong>one</strong> of the species in the five genera considered<br />
in this paper show similar extensions. Examination of a type specimen of D.<br />
dioxus shows that this species should not be placed in the genus Dynamenella.<br />
Hansen stated (1905: 126) that he examined an adult male and an immature<br />
male oi Dynamenella perforata. He did not examine an ovigerous female of this<br />
species (the type of the genus) but included specimens oi Dynamene eatoni Miers<br />
(actually an Ischyromene) in Dynamenella. If Hansen w<strong>as</strong> examining specimens<br />
of Ischyromene and including them in Dynamenella, this would explain why he<br />
stated "marsupial lamellae overlap each other somewhat" for Dynamenella,<br />
when in fact Dynamenella lacks oostegites. It would also explain why Hansen<br />
grouped Dynamenella with those genera lacking an articulation on the exopod of<br />
pleopod 3, when D. perforata possesses such an articulation.<br />
Species of Dynamenella vary in the presence or absence of an articulation on<br />
the exopod of pleopod 3, and in the shape of the pleotelsonic foramen. With the<br />
exception of Monod's record of D. platura, the remaining Old World species of<br />
Dynamenella lack an exopodal articulation, while the New World species possess<br />
<strong>one</strong>. With pleotelsonic foramina being variable in other genera (e.g., Ischyromene)<br />
it is not felt by the authors that the pleopodal articulation al<strong>one</strong> can be<br />
considered <strong>as</strong> a b<strong>as</strong>is for systematic separation above species level in this group.<br />
The form of the uropodal rami in species of Dynamenella varies in a manner<br />
similar to that shown by Ischyromene.<br />
Geographically, the species of Dynamenella appear to be found in tropical and<br />
subtropical locations, but at present are only represented on the African continent<br />
by records oi D. savignyi from the co<strong>as</strong>t of the Red Sea in Egypt (Fig. 7B).<br />
Synonyms.—N<strong>one</strong>.<br />
Dynamenella liochroea, new species<br />
Material Examined.—Holotype adult male, 4.14 mm (Q.M.: W.7932). Type-locality: C<strong>as</strong>uarina Beach,<br />
Lizard Island, Queensland (14°40'S, 145°30'E). Crevice in beach rock. Upper shore. Coll. D. M.<br />
Holdich, 12.vi. 1976. Paratypes.—From type-locality. Collection details <strong>as</strong> above. 4 adult males, 6<br />
subadult males (<strong>one</strong> <strong>as</strong> Q.M.: W.7933), 10 ovigerous females, 12 nonovigerous females, 3 juveniles;—<br />
C<strong>as</strong>uarina Beach, Lizard Island. In fine sand and around b<strong>as</strong>e of beach rock. Intertidal. Coll. D. M.<br />
Holdich, 07.vi. 1976. 4 adult males, 6 ovigerous females, 11 nonovigerous females, 5 juveniles;—<br />
C<strong>as</strong>uarina Beach, Lizard Island. In crevices in beach rock. Midshore. Coll. D. M. Holdich, 09.vi. 1976.<br />
8 adult males, 1 subadult male, 13 ovigerous females, 6 nonovigerous females (<strong>one</strong> <strong>as</strong> Q.M.: W.7934),<br />
6 juveniles;—South C<strong>as</strong>uarina Beach, Lizard Island. In fine sand. Intertidal. CoU. D. M. Holdich,
92 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
09.vi. 1976. 2 adult males, 3 nonovigerous females;—C<strong>as</strong>uarina Beach, Lizard Island. On beach rock<br />
on sand. Intertidal. Coll. D. M. Holdich, 10.vi. 1976. 1 adult male, 1 subadult male (supplying 1<br />
microslide, Q.M.: W.7935), 2 nonovigerous females, 2 juveniles;—C<strong>as</strong>uarina Beach, Lizard Island.<br />
Beach rock. Upper shore. CoU. D. M. Holdich, 12.vi. 1976. 3 adult males, 1 ovigerous female (<strong>as</strong><br />
Q.M.: W.7936);—West Point, Lizard Island. In rock crevice. Upper shore. Coll. D. M. Holdich,<br />
13.vi. 1976. 1 juvenile;—Ramsay Bay, Hinchinbrook Island, Queensland (18°22'S, 146°14'E). In sand.<br />
Intertidal. Coll. N. L. Bruce, 22.viii.1978. 1 adult male;—Picnic Bay, Magnetic Island, Townsville,<br />
Queensland (19°10'S, 146°50'E). From semipermanent logs and pieces of wood at top of dead coral<br />
z<strong>one</strong>. Midshore. Coll. D. M. Holdich, 09.vii.l976. 3 adult males (<strong>one</strong> supplying three microslides,<br />
Q.M.: W.7937), 1 ovigerous female, 1 nonovigerous female;—PaUarenda, Townsville. Among pink<br />
barnacles on wooden pier pile. Midshore. Coll. D. M. Holdich, 14.v. 1976. 1 subadult male, 2 ovigerous<br />
females, 2 nonovigerous females, 3 juveniles;—PaUarenda, Townsville. From pink striped barnacles<br />
and small rock oysters on rocks below spl<strong>as</strong>h z<strong>one</strong>. Upper shore. Coll. D. M. Holdich, 14.v. 1976. 1<br />
adult male, 1 nonovigerous female;—Kissing Point, Townsville. In sand by rocks at edge of pool.<br />
Intertidal. Coll. D. M. Holdich, 11.v. 1976. 14 juveniles;—Kissing Point, Townsville. From sand and<br />
rocks around edge of swimming pool. Intertidal. Coll. D. M. Holdich, 10.vii.l976. 1 adult male, 1<br />
subadult male, 3 nonovigerous females, 2 juveniles;—Kenny (South of Mission Beach), Kurrimine,<br />
Queensland (17°54'S, 146°05'E). Under st<strong>one</strong>s on sand near jetty. Midshore. Coll. D. M. Holdich,<br />
19.V. 1976. 14 adult males, 11 subadult males, 28 nonovigerous females, 6 juveniles;—Kenny, Kurrimine.<br />
In fine sand. Midshore. CoU. D. M. Holdich, 19.v. 1976. 2 subadult males, 1 immature male,<br />
1 nonovigerous female;—Kurrimine. In red filamentous alga on red sandst<strong>one</strong> rocks. Midshore. CoU.<br />
D. M. Holdich, 19.v. 1976. 3 adult males, 1 nonovigerous female;—Yorkey's Knob, Cairns, Queensland<br />
(16°51'S, 145°43'E). From tree trunk (bored by Teredo sp.) on sand by jetty. Intertidal. CoU. D.<br />
M. Holdich, 28.V. 1976. 2 adult males;—Yorkey's Knob, Cairns. In fine sand by rocks. Midshore.<br />
Coll. D. M. Holdich, 28.v. 1976. 6 adult males, 4 subadult males, 3 nonovigerous females, 9 juveniles.<br />
Etymology.—The specific name liochroea is derived from the Greek<br />
leios + chroia, i.e., smooth -I- skin.<br />
Description of Adult Male (Fig. 2a-k, m; Fig. 3a-h).—Dynamenella with pleon<br />
bearing low, transverse tubercle either side of midline. Pleotelson smooth in<br />
dorsal view, but appearing slightly rugose in lateral view. Subapical foramen<br />
circular, directed anterodorsally, separated from apex by slit which is closed<br />
anteriorly and widens posteriorly. Ventral margins of pleotelson flattening posteriorly,<br />
not extending to meet in midline at apex.<br />
Appendages: Antennule with peduncular article 1 twice length and twice<br />
breadth of article 2; article 3 half length of articles 1 and 2 together; flagellum<br />
9-articled. Antennal peduncular articles 2, 4, and 5 incre<strong>as</strong>ing in length; articles<br />
1 and 3 short; all articles subequal in breadth; I2-articled flagellum extending to<br />
level of pereonite 3. Mouthparts of usual sphaeromatid form but distal molar<br />
region of left mandible with a conical tooth which fits into an indentation in<br />
corresponding region of right mandible. Endite of maxilliped with numerous surface<br />
microtrichia; palp articles 2 to 4 with setose lobes. Pereopod 1 robust. Pereopods<br />
2 and 3 very slender. Pereopod 4 robust. Pereopods 5, 6, and 7 becoming<br />
slender and incre<strong>as</strong>ing in length. Merus, carpus, and propodus of all pereopods<br />
with dense, inferior mats of setae; b<strong>as</strong>is and ischium with superior setae; all<br />
dactyls with simple accessory unguis. Penes fused at b<strong>as</strong>e, tapering distally to<br />
acute tips with thickened regions midway along external margins. Pleopod 1 with<br />
distal margin of endopod broadly rounded, internal margin thickened, convex.<br />
Endopod of pleopod 2 with appendix m<strong>as</strong>culina flattened, broad proximaUy, tapering<br />
to narrowly rounded tip just beyond ramal apex. Exopod of pleopod 3<br />
lacking an articulation. B<strong>as</strong>es of pleopods 1 to 3 each bearing three internal<br />
coupling hooks; all rami with long, terminal, plumose setae. Endopod of pleopod<br />
4 apically truncate. Exopod of pleopod 5 with a subterminal articulation and<br />
three, finely toothed, internodistal bosses. Uropodal rami subequal, broad, rounded,<br />
extending well beyond apex of pleotelson.
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 93<br />
Fig. 2. Dynamenella liochroea, new species. Adult male (a) dorsal, (b) lateral, (c) right mandible,<br />
(d) left mandible, (e) maxilliped, (f) penes, (g) ventral, pleotelson, (h) posterior, pleotelson, (i) antenna,<br />
(j) antennule, (k) T.S. through pleotelsonic margin at level of bar on 'g,' (m) pleopod 2.—Subadult<br />
male (1) penes, (n) pleopod 2, (p) posterior, pleotelson.—Ovigerous female (o) T.S. through pleotelsonic<br />
margin at level of bar on 's,' (q) lateral, (r) dorsal, (s) ventral, pleotelson, (t) posterior, pleotelson.<br />
Each scale line represents 1 mm.
94 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Description of Ovigerous Female (3.00 mm) (Fig. 2o, q-t).—Dynamenella with<br />
pleon and pleotelson smooth. Posteromedial margin of pleotelson with shallow<br />
notch. Ventral margins of pleotelson lacking outcurved ridges; flattening terminally.<br />
Endopod of uropod narrow with acutely rounded apex extending to level<br />
of apex of pleotelson; exopod % length of endopod with serrate external and<br />
distal margins and rounded apex.<br />
Description of Nonovigerous Female.—As above but lacking brood pouch.<br />
Description of Sub adult Male (Fig. 21, n, p).—Resembling nonovigerous female,<br />
but dorsal to apical notch of pleotelson having narrow, vertical band of transparent<br />
cuticle. Penes obvious but half length of adult penes and separate to b<strong>as</strong>e,<br />
lacking thickened regions. Endopod of pleopod 2 with appendix m<strong>as</strong>culina visible<br />
through cuticle along internal margin, but not free; internal margin extended only<br />
slightly at apex with short terminal seta.<br />
Description of Immature Male (i.e., stage before subadult male).—Resembling<br />
small nonovigerous female but bearing penes. Penes short, separate to b<strong>as</strong>e, each<br />
Wi times <strong>as</strong> long <strong>as</strong> broad. No indication of appendix m<strong>as</strong>culina.<br />
Colour of Specimens in Alcohol.—Variable. From uniform cream with only few<br />
chromatophores, to dark, mottled purple-brown. Many individuals cream, with<br />
patches of dark colour. All individuals lacking chromatophores on ventral pereon<br />
and pereopods.<br />
Distribution and Habitat.—Found only on the mainland co<strong>as</strong>t of Queensland<br />
from Townsville north to Cairns, and on the continental islands. Lizard Island<br />
and Magnetic Island. Found in wood, amongst filamentous red algae, in beach<br />
rock and rock crevices on upper and middle shore, under st<strong>one</strong>s on sand and in<br />
fine sand around rocks on the midshore. Present on a variety of shores, usually<br />
among fine sand and rocks.<br />
Remarks.—Compared to other species of Dynamenella, D. liochroea, new<br />
species, resembles D. perforata but differs in having the pleotelson smooth dorsally,<br />
not finely tuberculate; in lacking an articulation on the exopod of pleopod<br />
3; and in having a circular foramen on the pleotelson.<br />
Synonyms.—N<strong>one</strong>.<br />
Dynamenella ptychura, new species<br />
Material Examined.—Holotype adult male, 2.5 mm (Q.M.: W.7939). Type-locality: Heron Island,<br />
Capricorn Group, Great Barrier Reef (23°25'S, 15r55'E). In beach rock crevice on upper shore. Coll.<br />
D. M. Holdich, 13.iv. 1976. Paratypes.—From type-locality. Collection details <strong>as</strong> above. 52 adult<br />
males (<strong>one</strong> supplying three microslides, Q.M.: W.7940), 39 subadult males (<strong>one</strong> <strong>as</strong> Q.M.: W.7941,<br />
another supplying microslide, Q.M.: W.7942), 2 immature males (<strong>one</strong> supplying microslide, Q.M.:<br />
W.7943), 281 ovigerous females (<strong>one</strong> <strong>as</strong> Q.M.: W.7944), 54 nonovigerous females (<strong>one</strong> <strong>as</strong> Q.M.:<br />
W.7945), 336 juveniles;—Heron Island. On Turbinaria sp. on inner reef flat. Intertidal. Coll. D. M.<br />
Holdich, 08.iv. 1976. 1 ovigerous female;—Heron Island. Under slab of beach rock. At level of high<br />
water neap tide. Coll. N. L. Bruce, 08.xii.l979. 2 adult males, 1 ovigerous female (Q.M.: W.8582);—<br />
Ramsay Bay, Hinchinbrook Island, Queensland (18°22'S, 146°14'E). In sand. Intertidal. CoU. N. L.<br />
Bruce, 22.viii.1978. 3 adult males, 3 subadult males, 3 ovigerous females, 2 nonovigerous females;—<br />
C<strong>as</strong>uarina Beach, Lizard Island. From crevices in beach rock. Midshore. Coll. D. M. Holdich,<br />
09.vi. 1976. 1 adult male, 2 ovigerous females;—C<strong>as</strong>uarina Beach, Lizard Island. From crevices in<br />
beach rock. Upper shore. Coll. D. M. Holdich, 12.vi. 1976. 3 adult males, 2 ovigerous females, 1<br />
nonovigerous female;—West Point, Lizard Island. In crevice. Upper shore. Coll. D. M. Holdich,<br />
13.vi. 1976. 19 adult males, 7 subadult males, 21 ovigerous females, 14 nonovigerous females;—Between<br />
Research Point and Freshwater Beach, Lizard Island. On clumps of oysters. Intertidal (Station<br />
78 Liz-PBW-3). Coll. Weate, Oldfield, and Berents, 16.iv.l978. 2 adult males. (AM.: P.27017).
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 95<br />
Etymology.—The specific name ptychura is formed from the Greek ptyche + ura,<br />
i.e., fold + tail.<br />
Description of Adult Male (Fig. 3i-n, r, s).—Dynamenella with pleon bearing<br />
low, transverse tubercle either side of midline. Pleotelson granulose, especially<br />
obvious in lateral view, with irregular, small tubercles over main dome; anterior<br />
region of dome, either side of midline, with short, granular, longitudinal ridge,<br />
with several raised tubercles posterior to this. Subapical foramen circular, directed<br />
anterodorsally, separated from apex by slit which is closed along entire<br />
length. Ventral margins of pleotelson folded <strong>as</strong> outcurved ridges extending posteromedially<br />
to meet in midline ventral to foramen.<br />
Appendages: Antennule <strong>as</strong> in D. liochroea. Antennal peduncle articles incre<strong>as</strong>ing<br />
in length 1 to 5; 13 flagellar articles extending to level of pereonite 3. Mouthparts<br />
similar to those of D. liochroea. Pereopods following general pattern of<br />
those of D. liochroea but less setose superiorly. Penes fused at b<strong>as</strong>e, tapering to<br />
external thickenings at midpoint, then margins subparallel in distal half, with<br />
apices rounded. Pleopod 1 <strong>as</strong> in D. liochroea but endopod thickened internoproximally,<br />
with internal margin slightly concave. Appendix m<strong>as</strong>culina of pleopod<br />
2 approximately \Vi times length of endopod, flattened, broad proximaUy, tapering<br />
only slightly to rounded apex. Pleopods 3 to 5 similar to those of Z). liochroea.<br />
Rami of uropod broad, extending well beyond apex of pleotelson; endopod with<br />
apex slightly obtusely angled and upcurved; exopod externally and distally serrate<br />
with apex rounded.<br />
Description of Ovigerous Female (2.72 mm) (Fig. 3q, t-w).—Dynamenella with<br />
pleon bearing low tubercle either side of midline. Pleotelson weakly rugose with<br />
main dome, either side of midline, bearing two short, low, longitudinal ridges,<br />
<strong>one</strong> posterior to other. Apex of pleotelson slightly extended, bearing small, subovate,<br />
posterodorsally directed foramen connected to the posteromedial margin<br />
by vertical, closed slit. Ventral margins of pleotelson lacking outcurved ridges,<br />
and flattened terminally. Endopod of uropod with acute apex not extending <strong>as</strong><br />
far <strong>as</strong> apex of pleotelson; exopod % length of endopod with serrate apex.<br />
Description of Nonovigerous Female.—As above but lacking brood pouch.<br />
Description of Subadult Male (Fig. 3o, p).—Resembling nonovigerous female,<br />
but with penes and endopod of pleopod 2 resembling those of subadult male of<br />
D. liochroea, except penes slightly broader terminally.<br />
Description of Immature Male (i.e., stage before subadult male).—Resembling<br />
small nonovigerous female but bearing penes. Penes short, separate to b<strong>as</strong>e, each<br />
twice <strong>as</strong> long <strong>as</strong> broad. No indication of appendix m<strong>as</strong>culina.<br />
Colour of Specimens in Alcohol.—Pale with fine grey chromatophores. Chromatophores<br />
absent from ventral pereon and pereopods.<br />
Distribution and Habitat.—Found on offshore islands. Common on Lizard Island<br />
and Heron Island.<br />
Found mainly on the upper shore in beach rock crevices, amongst rock oysters<br />
and in sand. All stages of the life history have been found in crevices.<br />
Remarks.—D. ptychura, new species, resembles D. platura but differs in having<br />
a circular foramen on the pleotelson; in lacking an articulation on the exopod of<br />
pleopod 3; and in the form of the appendix m<strong>as</strong>culina. The female of D. platura<br />
shows a simple notch on the pleotelson, not a foramen <strong>as</strong> in D. ptychura.
96<br />
JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Fig. 3. Dynamenella liochroea, new species. Adult male (a) pleopod 1, (b) pleopod 3, (c) pleopod<br />
4, (d) pleopod 5, (e) pereopod 1, (f) pereopod 4, (g) pereopod 7, (h) pereopod 2. Dynamenellaptychura,<br />
new species. Adult male (i) lateral, (j) dorsal, (k) pleopod 2, (1) T.S. through pleotelsonic margin at<br />
level of bar on 'n,' (m) posterior, pleotelson, (n) ventral, pleotelson, (r) pleopod 1, (s) penes.—<br />
Subadult male (o) pleopod 2, (p) penes.—Ovigerous female (q) T.S. through pleotelsonic margin at<br />
level of bar on 'u,' (t) posterior, pleotelson, (u) ventral, pleotelson, (v) lateral, (w) dorsal. Each scale<br />
line represents 1 mm.
Synonyms.—N<strong>one</strong>.<br />
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 97<br />
Dynamenella trachydermata, new species<br />
Material Examined.—Holotype adult male, 2.68 mm (Q.M.: W.7946). Type-locality: Kissing Point,<br />
Townsville, Queensland (19°14'S, 146°48'E). From rock crevice above red alga z<strong>one</strong>. Upper shore.<br />
Coll. D. M. Holdich, 11.v. 1976. Paratypes.—From type-locality. Collection details <strong>as</strong> above. 2 adult<br />
males (<strong>one</strong> supplying three microslides, Q.M.: W.7947), 144 ovigerous females (<strong>one</strong> <strong>as</strong> Q.M.: W.7948),<br />
6 nonovigerous females, 1 juvenile;—Kissing Point, Townsville. In sand by rocks at edge of pool.<br />
Intertidal. Coll. D. M. Holdich, 11.v. 1976. 1 ovigerous female;—Kissing Point, Townsville. From<br />
sand and rocks around edge of swimming pool. Intertidal. Coll. D. M. Holdich, 10.vii.l976. 5 adult<br />
males, 1 subadult male;—Kissing Point, Townsville. From rock crevices by swimming pool. Upper<br />
shore. Coll. D. M. Holdich, 10.vii.l976. 1 adult male, 57 ovigerous females, 13 juveniles;—Ramsay<br />
Bay, Hinchinbrook Island, Queensland. In sand. Intertidal. Coll. N. L. Bruce, 22.viii.1978. 1 ovigerous<br />
female;—Pallarenda, Townsville. Among pink barnacles on wooden pier pile. Midshore level.<br />
Coll. D. M. Holdich, 14.v. 1976. 4 adult males, 1 ovigerous female, 3 nonovigerous females;—Pallarenda,<br />
Townsville. From pink striped barnacles and small rock oysters on rocks below spl<strong>as</strong>h z<strong>one</strong>.<br />
Upper shore. Coll. D. M. Holdich, 14.v. 1976. 8 adult males, 2 subadult males, 74 ovigerous females,<br />
2 nonovigerous females, 8 juveniles;—Kurrimine, Queensland. On soft red sandst<strong>one</strong> rock. Upper<br />
shore. Coll. D. M. Holdich, 18.v. 1976. 6 adult males, 7 ovigerous females, 1 ovigerous female with<br />
penes of subadult male form, 2 nonovigerous females;—Kurrimine. In red weed on red rocks. Midshore.<br />
Coll. D. M. Holdich, 19.V.1976. 1 adult male, 1 ovigerous female, 2 juveniles;—Palm Beach,<br />
Cairns, Queensland. On rock oysters and red algae on vertical face. Intertidal. CoU. D. M. Holdich,<br />
25.V. 1976. 9 subadult males, 3 ovigerous females, 26 nonovigerous females, 58 juveniles;—Palm<br />
Beach, Cairns. In rock crevice in oyster z<strong>one</strong>. Intertidal. Coll. D. M. Holdich, 27.v. 1976. 4 adult<br />
males, 1 subadult male, 25 ovigerous females, 5 nonovigerous females;—Clifton Beach (south end).<br />
Cairns. In log bored by Teredo sp. Intertidal. Coll. D. M. Holdich, 30.v. 1976. 1 adult male, 2 subadult<br />
males;—Yorkey's Knob, Cairns. From log, bored by Teredo sp., on sand by jetty. Intertidal. Coll.<br />
D. M. Holdich, 28.v. 1976. 5 adult males, 3 subadult males (<strong>one</strong> <strong>as</strong> Q.M.: W.7949, another supplying<br />
1 microslide, Q.M.: W.7950), 2 ovigerous females, 3 nonovigerous females (<strong>one</strong> <strong>as</strong> Q.M.: W.7951),<br />
4 juveniles;—Yorkey's Knob, Cairns, In crevice below rock oyster z<strong>one</strong>. Intertidal. Coll. D. M.<br />
Holdich, 28.v. 1976. 3 adult males, 4 ovigerous females, 2 nonovigerous females.<br />
Etymology.—The specific name trachydermata is derived from the Greek trachydermon,<br />
i.e., rough skin.<br />
Description of Adult Male (Fig. 4a-j).—Dynamenella with dorsal body surface<br />
granular. Pleon bearing pronounced, transverse tubercle either side of midline.<br />
Pleotelson obviously tuberculate over main dome. Subapical foramen elliptical;<br />
directed anterodorsally; separated from apex by slit which is closed anteriorly<br />
and widens posteriorly. Ventral margins of pleotelson flattening posteriorly, not<br />
extending to meet in midline at apex.<br />
Appendages: Antennule, antenna, and mouthparts <strong>as</strong> for those of D. liochroea.<br />
Pereopods following general pattern of those of D. liochroea, superior setae longest<br />
on b<strong>as</strong>is and ischium of pereopod 7. Penes <strong>as</strong> in D. liochroea. Pleopod 1 <strong>as</strong><br />
in D. liochroea but endopod thickened internoproximally with internal margin<br />
straight. Appendix m<strong>as</strong>culina flattened, broad proximally, tapering to narrowly<br />
rounded apex just beyond apex of endopod of pleopod 2. Pleopods 3 to 5 similar<br />
to those of D. liochroea, but terminal article of exopod of pleopod 5 relatively<br />
shorter. Rami of uropod simlar to those of D. ptychura.<br />
Description of Ovigerous Female (2.45 mm) (Fig. 4n-q).—Dynamenella with<br />
pleon very slightly raised in midline. Pleotelson weakly rugose on main dome;<br />
apex not extended, bearing pronounced vertical incision with subparallel, lateral<br />
margins. Ventral margins of pleotelson lacking outcurved ridges and flattened<br />
terminally. Endopod of uropod with narrowly rounded apex extending to level<br />
of apex of pleotelson; exopod % length of endopod with rounded apex.<br />
Description of Nonovigerous Female.—As above but lacking brood pouch.
98<br />
JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Fig. 4. Dynamenella trachydermata, new species. Adult male (a) dorsal, (b) lateral, (c) antenna, (d)<br />
antennule, (e) posterior, pleotelson, (f) T. S. through pleotelsonic margin at level of bar on 'g,' (g)<br />
ventral, pleotelson, (h) pleopod 1, (i) pleopod 2, (j) penes.—Subadult male (k) pleopod 2, (1) penes<br />
(not to scale with 'j').—Ovigerous female (m) T.S. through pleotelsonic margin at level of bar on 'n,'<br />
(n) ventral, pleotelson, (o) posterior, pleotelson, (p) lateral, (q) dorsal. Each scale line represents 1
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 99<br />
Description ofSubadult Male (Fig. 4k, 1).—Resembling nonovigerous female, but<br />
with penes resembhng those of subadult male of D. liochroea. Endopod of pleopod<br />
2 with future appendix m<strong>as</strong>culina visible through cuticle, but not free; apex<br />
bearing short seta but not extended.<br />
Colour of Specimens in Alcohol.—Variable. From mottled dark grey over entire<br />
dorsal surface to sandy yellow with fine black chromatophores. Scattered chromatophores<br />
over ventral pereon but pereopods lacking chromatophores.<br />
Distribution and Habitat.—Found at various mainland localities from Townsville<br />
north to Cairns, and at Hinchinbrook Island.<br />
Found amongst filamentous red algae, wood and rock crevices from upper to<br />
midshore. Also amongst rock oysters and barnacles on upper and middle shore,<br />
and in sand around midshore rocks. All stages of life history found in cryptic<br />
habitats. Present on moderately sheltered and exposed rocky shores, usually<br />
where fine sand is present.<br />
Remarks.—D. trachydermata, new species, most closely resembles D. liochroea,<br />
new species, but differs in having a tuberculate pleotelson; in having a relatively<br />
longer endopod to pleopod 1; and in the form of the female pleotelson.<br />
Paradella, new genus<br />
Diagnosis.—Eubranchiate Sphaeromatidae with antennular peduncle article 1 not<br />
extended anteriorly <strong>as</strong> a plate. Both sexes with dorsal pereon and pleon lacking<br />
processes. Both sexes with both rami of uropod lamellar; endopod greater than<br />
half length of exopod. All pereopods with accessory unguis simple, not bifid.<br />
Exopod of pleopod 3 with an articulation. Sexual dimorphism obvious.<br />
Adult Male.—Penes long, tapering, fused at b<strong>as</strong>e. Appendix m<strong>as</strong>culina narrow,<br />
lateral margins subparallel; arising from internoproximal angle of endopod of<br />
pleopod 2 and extending well beyond apex of endopod. Apex of pleotelson with<br />
dorsally directed foramen connected to apex by short, narrow slit. Ventral margins<br />
of pleotelson with broad, low, straight, outfolded ridges not meeting in posterior<br />
midline. Uropods broader than those of females and immature specimens.<br />
Ovigerous Female.—Mouthparts not metamorphosed. Pleotelson apically extended,<br />
not broadly rounded. Uropods narrower than those of adult male. Brood<br />
pouch formed from two opposing pockets covering entire ventral pereon, and<br />
opening in midline between fourth pereopods. Each pereopod 4 bearing short<br />
oostegite, not reaching animal's midline.<br />
Type-species.—Paradella octaphymata, new species<br />
Etymology.—The generic name Paradella is a combination of the Greek<br />
para -\- d'ella (feminine), i.e., beside -I- contraction of 'Dynamenella\<br />
Recommendation<br />
b<strong>as</strong>ed on<br />
Additional species: Current name: examination of:<br />
P. acutitelson (Menzies and Glynn,<br />
1968), new combination Dynamenella acutitelson Literature<br />
P. bakeri (Menzies, 1962b),<br />
new combination D. bakeri Types<br />
P. dianae (Menzies, 1962a),<br />
new combination D. dianae Specimens
100 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
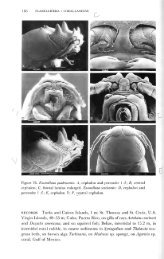
Fig. 5. Paradella octaphymata, new genus, new species. Adult male (a) dorsal, (b) lateral, (c)<br />
antennule, (d) ventral, pleotelson, (e) T.S. through pleotelsonic margin at level of bar on 'd,' (f)<br />
antenna, (g) posterior, pleotelson, (h) pleopod 1, (i) pleopod 2, (k) maxilliped, (m) penes.—Subadult<br />
male (j) pleopod 2 (not to scale with 'h' and 'i'), (1) penes.—Ovigerous female (n) posterior, pleotelson,<br />
(o) ventral, pleotelson, (p) T.S. through pleotelsonic margin at level of bar on 'o,' (q) lateral, (r)<br />
dorsal. Each scale line represents 1 mm.
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 101<br />
Recommendation<br />
b<strong>as</strong>ed on<br />
Additional species: Current name: examination of:<br />
P. pllcatura (Glynn, 1970),<br />
new combination D. pllcatura Literature<br />
P. quadripunctata (Menzies and<br />
Glynn, 1968), new combination D. quadripunctata Literature<br />
P. setosa (Glynn, 1968b),<br />
new combination D. setosa Literature<br />
P. tumicauda (Glynn, 1970),<br />
new combination D. tumicauda Literature<br />
Remarks.—Dynamenella acutitelson appears to be b<strong>as</strong>ed on females and subadult<br />
males of <strong>one</strong> or more species oiParadella, new genus. Adult males of D. setosa<br />
have never been illustrated, but figured specimens of this species appear to belong<br />
in Paradella, new genus. Most species referred above to Paradella, new genus,<br />
show greater similarity in dorsal view to <strong>one</strong> another than to species of Dynamenella.<br />
These species are: Paradella octaphymata, new species (see below), P.<br />
bakeri, P. quadripunctata, and P. tumicauda. The species P. dianae and P.<br />
pllcatura are, in dorsal view, more suggestive oi Dynamenella species.<br />
Species of Paradella are distinguished from species of Dynamenella most obviously<br />
by the presence in the former of small oostegites in ovigerous females<br />
and by the form of the ventral margins of the pleotelson in adult males. The<br />
appendix m<strong>as</strong>culina in species of Paradella is long and narrow with the margins<br />
subparallel, and in subadult males the fused appendix m<strong>as</strong>culina extends beyond<br />
the endopodal apex much further than in species oi Dynamenella.<br />
Paradella is known only from the Americ<strong>as</strong>, where it occurs in both tropical<br />
and temperate regions, and from Australia and the Marshall Islands (Fig. 7c).<br />
Synonyms. —N<strong>one</strong>.<br />
Paradella octaphymata, new species<br />
Material Examined.—Holotype adult male, 4.14 mm (Q.M.: W.7925). Type-locality: West Point,<br />
Lizard Island, Queensland (14°14'S, 145°30'E). Among barnacles in rock crevice. Lower shore. Coll.<br />
D. M. Holdich, 13.vi. 1976. Paratypes.—From type-locality. Collection details <strong>as</strong> above. 7 adult males<br />
(<strong>one</strong> supplying three microslides, Q.M.: W.7926), 2 subadult males (<strong>one</strong> <strong>as</strong> Q.M.: W.7927, the other<br />
supplying microslide, Q.M.: W.7928), 15 ovigerous females (<strong>one</strong> <strong>as</strong> Q.M.: W.7929), 7 nonovigerous<br />
females (<strong>one</strong> <strong>as</strong> Q.M.: W.7930), 6 juveniles;—Kapock Cove, Lizard Island. From rock scrapings<br />
below rock oyster z<strong>one</strong>. Intertidal. Coll. D. M. Holdich, 08.vi. 1976. 1 adult male, 5 juveniles;—West<br />
Point, Lizard Island. In rock crevice on rock platform. Lower shore. Coll. D. M. Holdich,<br />
09.vi.l976.11 ovigerous females, 1 nonovigerous female;—South C<strong>as</strong>uarina Beach, Lizard Island. In<br />
crevice. Midshore. Coll. D. M. Holdich, 12.vi. 1976. 1 ovigerous female;—West Point, Lizard Island.<br />
Among rock oysters. Intertidal. Coll. D. M. Holdich, 13.vi. 1976. 1 immature male (supplying microslide,<br />
Q.M.: W.7931), 3 ovigerous females, 1 juvenile;—Between Research Point and Freshwater<br />
Beach, Lizard Island. On clumps of oysters. Intertidal (station 78 Liz-PBW-3). Coll. Weate, Oldfield,<br />
and Berents, 16.iv.1978. 1 adult male, 4 ovigerous females, 1 nonovigerous female (A.M.: P.27018);—<br />
Alma Bay, Magnetic Island, Townsville. In rock oyster z<strong>one</strong> and below. Intertidal. Coll. D. M.<br />
Holdich, 28.iv. 1976 . 2 adult males, 10 ovigerous females, 4 nonovigerous females.<br />
Etymology.—The specific name octaphymata is derived from the Greek<br />
octo + phyma, i.e., eight + tubercle.<br />
Description of Adult Male (Fig. 5a-i, k, m).—Paradella with posterior margins<br />
of pereonites 6 and 7 bearing transverse ridge with medial indentation. Pleon<br />
bearing prominent, transverse tubercle either side of midline. Each side of pos-
102 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
terior pleonal margin, lateral to point of articulation with pleotelson, bearing two<br />
suture lines merging in their posterior halves to produce a Y-shape with only <strong>one</strong><br />
line reaching posterior margin. Pleotelson granulose with eight prominent tubercles<br />
arranged in two transverse rows of four. Subapical foramen subelliptical,<br />
connected to apex by slit which is open dorsally and widens posteriorly. Slit<br />
closed posteroventrally. Ventral margins of pleotelson on each side with broad,<br />
low, outcurved ridge extending to apex; ridges not touching in midline.<br />
Appendages: Antennular peduncle articles 1 and 2 twice breadth of article 3;<br />
flagellum 10-articled. Antennal peduncle articles 1 to 5 incre<strong>as</strong>ing in length; 16articled<br />
flagellum extending to level of pereonite 3. Mouthparts of usual sphaeromatid<br />
form; distal molar regions of mandibles lacking obvious projections/<br />
depressions; mandibular palps present; maxillipedal palp articles 2 to 4 with setose<br />
lobes. Pereopods 2 and 3 not <strong>as</strong> markedly slender <strong>as</strong> those of preceding<br />
species. Penes fused at b<strong>as</strong>e, proximal margins subparallel, distal margins tapering<br />
evenly to acute apex. Pleopod 1 with distal margin of endopod extended<br />
beyond exopod, narrowly rounded; internal margin bearing short, nonmarginal<br />
setae. Endopod of pleopod 2 with appendix m<strong>as</strong>culina almost twice length of<br />
endopod, narrow, with margins subparallel and rounded apex. B<strong>as</strong>es of pleopods<br />
1 to 3 each bearing three internal coupling hooks. Endopod of pleopod 4 with<br />
apex acutely tapering and internally deflected, with single apical spine. Exopod<br />
of pleopod 5 with subterminal articulation and three, finely toothed, internodistal<br />
bosses. Endopod of uropod just longer than exopod, extending just beyond apex<br />
of pleotelson, upcurved terminaUy; both rami with outer margins serrate, apices<br />
obtusely angled.<br />
Description of Ovigerous Female (3.4 mm) (Fig. 5n-r).—Paradella with pleon<br />
bearing low tubercle either side of midline. Pleotelson granular with eight low<br />
tubercles arranged in two transverse rows of four; apex slightly extended with<br />
narrow notch reaching half way up overhanging posterior face. Ventral margins<br />
of pleotelson lacking outcurved ridges, not flattening terminally. Endopod of<br />
uropod reaching level of apex of pleotelson; exopod just shorter than endopod,<br />
with serrate external and distal margins; both rami acutely rounded apically.<br />
Description of Nonovigerous Female.—As above but lacking brood pouch.<br />
Description of Subadult Male (Fig. 5j, 1).—ResembHng nonovigerous female but<br />
with penes half length of adult penes, fused at b<strong>as</strong>e, and tapering evenly to<br />
narrowly rounded apices. Endopod of pleopod 2 bearing appendix m<strong>as</strong>culina<br />
slightly longer than ramus but appendix free only for short distance proximal and<br />
distal to apex of endopod, otherwise fused completely with ramus. Tip of appendix<br />
m<strong>as</strong>culina with short seta.<br />
Description of Immature Male (i.e., stage before subadult male).—Resembling<br />
small nonovigerous female but bearing penes. Penes short, separate to b<strong>as</strong>e, each<br />
twice <strong>as</strong> long <strong>as</strong> wide. No indication of appendix m<strong>as</strong>culina present.<br />
Colour of Specimens in Alcohol.—Mottled dark reddish brown over dorsal surface.<br />
Ventral pereon and pereopods lacking chromatophores. Chromatophores<br />
absent dorsally from regions of muscle attachment.<br />
Distribution and Habitat.—Found on the continental islands. Lizard Island and<br />
Magnetic Island.<br />
Found on the mid to lower shore amongst barnacles and rock oysters, and in<br />
rock crevices. Associated with stable, moderately exposed, rocky shores. All<br />
stages of the life history found in cryptic habitats.
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 103<br />
Remarks.—Paradella octaphymata, new species, most closely resembles P. bakeri<br />
but differs in having a circular, not a transverse, pleotelsonic foramen; in<br />
having Y-shaped, not V-shaped, sutures on the posterior margin of the pleon;<br />
and in having eight <strong>large</strong> tubercles on the pleotelson. (Specimens of P. bakeri<br />
have a small tubercle between the lateral pair of <strong>large</strong> tubercles.)<br />
Paradella dianae (Menzies, 1962), new combination<br />
Dynamenopsis dianae Menzies, 1962a: 341. Glynn, 1968a: 573, 575, 583, 584, 591, 622. Schultz, 1969:<br />
123.<br />
Dynamenella dianae: Menzies and Glynn, 1968: 63, 113. Glynn, 1970: 18, 20, 22, 22-26. Iverson,<br />
1974: 166. Pires, 1980b: 133-140.<br />
Material Examined.—PaUarenda, Townsville, Queensland. Among pink barnacles on wooden pier<br />
pile. Midshore. Coll. D. M. Holdich, 14.V.1976. 2 adult males (<strong>one</strong> <strong>as</strong> Q.M.: W.7938), 1 subadult male<br />
(Q.M.: W.7938), 1 ovigerous female (Q.M.: W.7938), 1 ovigerous female with penes of subadult male<br />
form;—PaUarenda, Townsville. From pink striped barnacles and small rock oysters on rocks below spl<strong>as</strong>h<br />
z<strong>one</strong>. Upper shore. Coll. D. M. Holdich, 14.v.1976. 1 subadult male;—^Jetty of "James Kirby," Townsville<br />
Harbour, Townsville. In barnacle tests from scrapings from steps, and concrete and copper sheathing<br />
on piles. Intertidal. Coll. D. M. Holdich, 01.vii.l976. 1 nonovigerous female (Q.M.: W.7938), 1<br />
juvenile;—Jetty of "James Kirby," Townsville Harbour. In scrapings of barnacles from steps and<br />
piles. Lower tide level. Coll. D. M. Holdich, ll.viii.l976. 1 subadult male, 3 ovigerous females, 2<br />
ovigerous females with penes of subadult male form, 14 nonovigerous females, 6 juveniles;—Dunwich,<br />
North Stradbroke Island, Brisbane. Under rock. Coll. N. L. Bruce, 19.vii.l978. 1 adult male, 2<br />
subadult males;—Dunwich, North Stradbroke Island, Brisbane. Coll. J. Cowger, ?.x.l971. 1 ovigerous<br />
female (no brood) (Q.M.: W.3746);—Goat Island. From rocks. Coll. N. L. Bruce, 21.vii.l978.<br />
1 juvenile;—Fremantle Bridge, Swan River, Western Australia (32°06'S, 115°50'E). Under st<strong>one</strong>s.<br />
Coll. N. L. Bruce, 10.vi. 1980. 19 adult males, 44 ovigerous females, 11 nonovigerous females,<br />
1 subadult male, 3 immature males, 101 juveniles;—Mayaguez, Puerto Rico. Sea wall at north end of<br />
bay. Intertidal. Coll. P. Glynn, 18.1.1966. 1 adult male, 1 ovigerous female, 1 half moulted female<br />
(U.S.N.M. 269486).<br />
Description of Adult Male (4.88 mm) (Fig. 6a-h, j, k).—Paradella with pleon<br />
bearing prominent tubercle either side of midline. Each side of posterior pleonal<br />
margin, lateral to point of articulation with pleotelson, bearing two suture lines<br />
merging in their posterior halves to produce a Y-shape with only <strong>one</strong> line reaching<br />
posterior margin. Pleotelson granulose, especially obvious in lateral view, with<br />
dome bearing anterior, longitudinal tubercle and posterior, elongate, oblique tubercle<br />
either side of midline. Subapical foramen V-shaped, directed dorsally, with<br />
anteromedial tooth bearing longitudinal ridge. Foramen closed posteriorly. Ventral<br />
margins of pleotelson on each side with low outcurved ridge extending to<br />
apex; ridges not touching in midline.<br />
Appendages: Antennule <strong>as</strong> in P. octaphymata but flagellum with 12 articles.<br />
Antenna <strong>as</strong> in P. octaphymata but flagellum with 13 articles, extending to level<br />
of pereonite 4. Mouthparts of usual sphaeromatid form; distal molar regions of<br />
mandibles lacking obvious projections/depressions; maxillipedal palp articles 2-4<br />
with setose lobes. Pereopod 1 robust; 2 and 3 slender; 4 robust; 5-7 becoming<br />
slender and incre<strong>as</strong>ing in length. Merus, carpus, and propodus of pereopods with<br />
inferior setae. Penes fused at b<strong>as</strong>e, with proximal margins subparallel; distally<br />
tapering to acute apices. Pleopod 1 with distal margin of endopod extended beyond<br />
exopod, acute; internal margin bearing short nonmarginal setae. Endopod<br />
of pleopod 2 with appendix m<strong>as</strong>culina twice length of endopod, narrow, with<br />
margins subparallel, tapering abruptly to narrowly rounded, externally deflected<br />
apex. B<strong>as</strong>es of pleopods 1 to 3 each bearing three internal coupling hooks. Endopod<br />
of pleopod 4 with apex extended, acute, internally deflected, with single<br />
apical spine. Exopod of pleopod 5 with subterminal articulation and three, finely
104 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Fig. 6. Paradella dianae (Menzies), new combination. Adult male (a) dorsal, (b) lateral, (c) ventral,<br />
pleotelson, (d) T.S. through pleotelsonic margin at level of bar on 'c,' (e) penes, (f) posterior, pleotelson,<br />
(g) pereopod 1, (h) pereopod 2, (j) pleopod 1, (k) pleopod 2.—Subadult male (i) penes, (o)<br />
pleopod 2.—Ovigerous female (1) T.S. through pleotelsonic margin at level of bar on 'm,' (m) ventral,<br />
pleotelson, (n) posterior, pleotelson, (p) lateral, (q) dorsal. Each scale line represents 1 mm.
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 105<br />
toothed internodistal bosses. Endopod of uropod extending beyond apex of pleotelson,<br />
dorsally concave with several small tubercles; exopod just shorter than<br />
endopod; both rami serrate in distal half.<br />
Description of Ovigerous Female (3.68 mm) (Fig. 61-n, p, q).—Paradella with<br />
pleon bearing low tubercle either side of midline. Pleotelson granulose with two<br />
elongate tubercles fused <strong>as</strong> oblique ridge either side of midline; apex slightly<br />
extended, opening <strong>as</strong> wide channel not visible in dorsal view. Ventral margins of<br />
pleotelson lacking outcurved ridges, not flattening terminally. Endopod of uropod<br />
extending to level of apex of pleotelson; exopod just shorter than endopod; both<br />
rami acutely rounded terminally with obvious marginal setae. Oostegites relatively<br />
smaller than those of P. octaphymata.<br />
Description of Nonovigerous Female.—As above but lacking oostegites.<br />
Description of Sub adult Male (Fig. 6i, o).—ResembHng nonovigerous female, but<br />
with penes and endopod of pleopod 2 resembling those of/*, octaphymata.<br />
Colour of Specimens in Alcohol.—Pale, with fine grey chromatophores. Chromatophores<br />
absent from ventral pereon and pereopods.<br />
Distribution and Habitat.—^Found on North Stradbroke Island, near Brisbane;<br />
in Townsville Harbour; at Pallarenda, just north of Townsville; and in Western<br />
Australia. This species h<strong>as</strong> also been recorded from Baja California (Menzies,<br />
1962a); Puerto Rico (Menzies and Glynn, 1968; Glynn, 1968a), Marshall Islands<br />
(Glynn, 1970), California (Iverson, 1974), and Brazil (Pires, 1980b). It is noteworthy<br />
that in Australia it h<strong>as</strong> been found only near international ports, often<br />
amongst sessile organisms belonging to groups commonly found attached to ships'<br />
hulls.<br />
This isopod is found amongst barnacles, bryozoans, and rock oysters on rocks<br />
and man-made structures from upper to lower shore, and tends to be <strong>as</strong>sociated<br />
with relatively stable rocky shores in moderately sheltered localities. All stages<br />
of the life history were found in cryptic habitats.<br />
This species h<strong>as</strong> a wide geographical distribution and elsewhere h<strong>as</strong> been found<br />
amongst intertidal green algae, empty polychaete tubes, and barnacle tests on<br />
both exposed and sheltered shores in Brazil (Pires, 1980b); under st<strong>one</strong>s, in empty<br />
barnacle tests, on rocks with algae, and under Chiton tuberculatus Linnaeus in<br />
Puerto Rico (Glynn, 1968a; Menzies and Glynn, 1968); w<strong>as</strong>hings from rocks on<br />
lagoon side of Fred Island, Marshall Islands (Glynn, 1970). Pires mentions that<br />
this species can live in polluted water, b<strong>as</strong>ing her <strong>as</strong>sumption on the fact that <strong>one</strong><br />
site of Menzies and Glynn w<strong>as</strong> near a polluted bay. It would appear that this<br />
species tolerates a wide variety of conditions, <strong>as</strong> Townsville Harbour is mildly<br />
polluted and the water in PaUarenda Bay can be of relatively low salinity.<br />
Dynamenopsis Baker, 1908<br />
Diagnosis.—Eubranchiate Sphaeromatidae with antennular article 1 not extended<br />
anteriorly <strong>as</strong> plate. Both sexes with pereon and pleon lacking dorsal processes.<br />
Coxal plates prominent, obvious; those of pereonite 6 with rounded posterior<br />
margins always overlapping those of pereonite 7. Apex of pleotelson with lateral<br />
margins curved ventrally and medially to meet, or almost meet, in midline producing<br />
either enclosed, posterodorsally directed foramen, or posterodorsally directed<br />
groove. Mandibles aberrant, robust; incisor processes stout, smoothly<br />
rounded; lacinia mobilis and setal rows absent; molar processes expanded <strong>as</strong>
106 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. I, 1982<br />
broad, ridged crushing plates. Pereopods subequal, stout, with accessory unguis<br />
short, smoothly rounded. Exopod of pleopod 3 with articulation.<br />
Adult Male.—Penes short, stout, separate to b<strong>as</strong>e. Appendix m<strong>as</strong>culina arising<br />
from internoproximal angle of endopod of pleopod 2 and extending beyond apex<br />
of endopod. Posterior margin of pereonite 7 markedly bilobed in dorsal view.<br />
Ovigerous Female.—Inadequately known.<br />
Type-species.—Dynamenopsis obtusa Baker, 1908.<br />
Recommendation<br />
b<strong>as</strong>ed on<br />
Additional species: Current name: examination of:<br />
D. varicolor Hurley and Jansen,<br />
1971 Dynamenopsis varicolor Type<br />
Remarks.—Originally described from South Australia, D. obtusa is here recorded<br />
from Western Australia (see Appendix). In addition, a female Dynamenopsis sp.<br />
is illustrated (Fig. 8k-o). This female differs in dorsal view from males of both<br />
D. obtusa and D. varicolor but <strong>as</strong> little is known about the degree of sexual<br />
dimorphism shown by species of Dynamenopsis, and since the female w<strong>as</strong> not<br />
found with males, specific identification remains undetermined. At the b<strong>as</strong>e of<br />
each of pereopods 2 to 4 this female bears a short, narrow, rigid oostegite, with<br />
indications of four pairs of apertures beneath the ventral cuticle between these<br />
structures. This specimen is apparently nonovigerous.<br />
Of the other species currently placed in the genus Dynamenopsis, D. dumerilii<br />
and D. platura have been placed in Dynamenella above; D. angolensis Kensley,<br />
1971, cannot be placed in any of the five genera considered here and its correct<br />
generic placement is unknown.<br />
Species of the genus Dynamenopsis are known only from New Zealand and<br />
South and Western Australia.<br />
Cymodocella Pfeffer, 1887<br />
Diagnosis.—Eubranchiate Sphaeromatidae with antennular peduncle article 1 not<br />
extended anteriorly <strong>as</strong> plate. Both sexes with pereon and pleon lacking dorsal<br />
processes. Both sexes with rami of uropod lamellar; endopod greater than half<br />
length of exopod. Apex of pleotelson with lateral margins curving ventrally and<br />
meeting in midline producing cylindrical, posteriorly directed aperture. All pereopods<br />
with simple accessory unguis, not markedly bifid. Exopod of pleopod 3<br />
without articulation. Sexual dimorphism not pronounced.<br />
Adult Male.—Penes short, stout, separate to b<strong>as</strong>e. Appendix m<strong>as</strong>culina arising<br />
from internoproximal angle of endopod of pleopod 2 and extending beyond apex<br />
of endopod. Exopod of uropod usually relatively longer than that of female.<br />
Ovigerous Female.—Mouthparts not metamorphosed. Exopod of uropod usually<br />
shorter than that of male. Brood pouch formed posteriorly <strong>as</strong> pocket, opening<br />
anteriorly at level of pereopod 4; and anteriorly <strong>as</strong> three pairs of <strong>large</strong> oostegites<br />
arising from pereopods 2 to 4 and overlapping well in midline. Brood not incubated<br />
in pouch thus formed, but in invaginations of ventral body wall.<br />
Type-species.—Cymodocella tubicauda Pfeffer, 1887.
Additional species:<br />
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 107<br />
C. algoensis (Stebbing, 1875)<br />
C. bicolor (Barnard, 1914)<br />
new combination<br />
C. cancellata Barnard, 1920<br />
C. capra Hurley and Jansen,<br />
1977<br />
C. diateichos Barnard, 1959<br />
C. egregia (Chilton, 1892)<br />
C. eutylos Barnard, 1954<br />
C. foveolata Menzies, 1962b<br />
C guarapariensis Silva, 1965<br />
C. magna Barnard, 1954<br />
C. pustulosa Barnard, 1914<br />
C. sublaevis Barnard, 1914<br />
Current name:<br />
Cymodocella algoensis<br />
Dynamenella bicolor<br />
Cymodocella cancellata<br />
Cymodocella capra<br />
Cymodocella diateichos<br />
Cymodocella egregia<br />
Cymodocella eutylos<br />
Cymodocella foveolata<br />
Cymodocella guarapariensis<br />
Cymodocella magna<br />
Cymodocella pustulosa<br />
Cymodocella sublaevis<br />
Recommendation<br />
b<strong>as</strong>ed on<br />
examination of:<br />
Literature<br />
Type<br />
Type<br />
Type<br />
Type<br />
Specimens<br />
Types<br />
Types<br />
Literature<br />
Types<br />
Types<br />
Types<br />
Remarks.—C. bicolor, new combination, resembles C. pustulosa but the pleotelson<br />
is relatively broader and the posterior margin of pereonite 7 is not bilobed.<br />
In the subadult male examined the posterior margins of the pleotelson curved<br />
ventrally and produced a posteriorly directed foramen, but did not meet in the<br />
midline, being separated by a narrow slit.<br />
Some species of Cymodocella (e.g., C. pustulosa and C. eutylos) have the<br />
coxal plates of pereonite 6 acute and deflected posteriorly, overlapping those of<br />
pereonite 7. Such overlapping h<strong>as</strong> been held in the p<strong>as</strong>t to be a distinguishing<br />
generic character of Dynamenopsis. Since this overlapping occurs in species of<br />
Cymodocella its importance <strong>as</strong> a generic character must be weakened, but in all<br />
specimens seen the genus Dynamenopsis shows this character in an extreme<br />
form. Species of Dynamenopsis have the posterior margins of the coxal plates of<br />
pereonite 6 broadly rounded, not acute, completely covering the lateral margins<br />
of pereonite 7.<br />
Of the other species currently placed in the genus Cymodocella, C. marginata<br />
and C. nipponica have been transferred to Dynamenella; and C. sapmeri, C.<br />
eatoni, C. huttoni, and C. ovalis have been transferred to Ischyromene (see<br />
above).<br />
Species of the genus Cymodocella occur mainly in the Antarctic and southern<br />
temperate regions; only C. guarapariensis and C. foveolata, from South America,<br />
are known from the tropics. No species are known from Australia (Fig. 7D).<br />
KEY TO ADULT SPECIMENS OF THE GENERA CONSIDERED ABOVE<br />
Apex of pleotelson with a posteriorly directed, ventrally closed (or almost closed) tubular<br />
foramen Cymodocella Pfeffer<br />
Apex of pleotelson with a simple channel or indentation, or a dorsally or posterodorsally<br />
directed foramen 2<br />
Coxal plates of pereonite 6 twice length of tergite, covering coxal plates of pereonite 7. Incisor<br />
processes of mandibles smoothly rounded Dynamenopsis Baker<br />
Coxal plates of pereonite 6 subequal in length to tergite. Coxal plates of pereonite 7 exposed.<br />
Incisor process of mandibles dentate 3<br />
Endopod of pleopod 1 thickened in internal half Ischyromene Racovitza<br />
Endopod of pleopod 1 not thickened 4
108 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Fig. 7. Known distribution of genera worldwide. (A) Ischyromene, (B) Dynamenella, A = exopod of<br />
pleopod 3 lacking articulation, • = exopod of pleopod 3 with articulation, (C) Paradella, (D)<br />
Cymodocella.<br />
Ventral margins of pleotelson of adult male bearing outcurved ridges which do not meet in<br />
posterior midline. Ovigerous female with a short oostegite at the b<strong>as</strong>e of pleopod 4<br />
Paradella, new genus<br />
Ventral margins of pleotelson of adult male either bearing outcurved ridges which meet in<br />
posterior midline, or lacking outcurved ridges. Ovigerous female lacking oostegites<br />
Dynamenella Hansen<br />
Sources of Error in Sphaeromatid Systematics.—Since Hansen (1905) very few<br />
workers have menti<strong>one</strong>d the brood pouch when describing new taxa. Many<br />
species of Dynamenella, Ischyromene, and Paradella (until now considered<br />
species of Dynamenella) could have been separated long ago if greater attention<br />
had been paid to this structure. The present authors consider the morphology of<br />
the brood pouch to be <strong>one</strong> of the single most important generic characters within<br />
the family Sphaeromatidae, and it should be described in detail whenever possible.<br />
The form of the penes of the adult male is also an important generic character.<br />
With males, confusion h<strong>as</strong> arisen because workers have <strong>as</strong>sumed that the presence<br />
of penes indicated the specimen w<strong>as</strong> an adult male. This is not so. The new<br />
species from Australia described here confirm that not only can subadult males<br />
have penes (and often a very different dorsal morphology when compared to the<br />
adult male), but also that penes can be obvious in immature males. This h<strong>as</strong> led<br />
in the p<strong>as</strong>t to descriptions b<strong>as</strong>ed on immature specimens (e.g., Dynamenella<br />
acutitelson and Dynamenella tropica), a situation which could have been avoided<br />
by ensuring that the specimens to be described had the appendix m<strong>as</strong>culina free<br />
and not partially or completely fused with the endopod of the pleopod.<br />
Even less desirable than describing species b<strong>as</strong>ed on immature males, in a
HARMSON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 109<br />
Fig. 8. Dynamenopsis obtusa Baker. Adult male (W. A.M.: 56-80) (a) dorsal, (b) lateral, (c) posterior,<br />
pleotelson, (d) T.S. through pleotelsonic margin at level of bar on 'e,' (e) ventral, pleotelson, (f)<br />
penes, (g) left mandible, anterior view, (h) right mandible, anterior view.—Adult male holotype<br />
(S.Aust.M.: C.359) (i) pereopod 1, (j) maxillipedal endite. Dynamenopsis sp. Nonovigerous female<br />
(W.A.M.: 57-80) (k) posterior, pleotelson, (1) T.S. through pleotelsonic margin at level of bar on 'm,'<br />
(m) ventral, pleotelson, (n) lateral, (o) dorsal. Each scale line represents 1 mm.
110 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
family where sexual dimorphism is common, is the description of species b<strong>as</strong>ed<br />
on females (at any stage of development). In some c<strong>as</strong>es it is difficult, if not<br />
impossible, to place females in the correct genus. The genus Sergiella Pires,<br />
1980a, w<strong>as</strong> founded on females and the fact that they appear to be females of a<br />
species of Paracerceis serves to demonstrate the undesirability of such descriptions.<br />
It is to be recommended, therefore, that descriptions of new species be b<strong>as</strong>ed<br />
on adult males. They should also include not only a full description of the male,<br />
but descriptions of ovigerous females and other stages of the life cycle whenever<br />
possible. Descriptions of the adult male penes and appendices m<strong>as</strong>culinae, and<br />
the brood pouch and mouthparts of the ovigerous female, should be made to<br />
prevent species from being referred to incorrect genera. Thorough re-examination<br />
of existing species should also be undertaken to facilitate much needed revision<br />
of this family of isopods.<br />
LIFE HISTORY AND ECOLOGY<br />
Until the present study the only Australian eubranchiate sphaeromatid about<br />
which details of the ecology are known w<strong>as</strong> Dynamene curalii Holdich and<br />
Harrison (1980c). Members of this genus are more commonly found in mid to<br />
lower shore cryptic habitats and algae in tidal localities of northwestern Europe,<br />
and similar habitats in the sublittoral region of the Mediterranean Sea (Holdich,<br />
1970). In addition, Dynamene ramuscula Baker h<strong>as</strong> been recorded from sponges<br />
in South Australia (Baker, 1908). In subtropical and tropical latitudes Dynamene<br />
appears to be absent from rocky shores and is replaced by Dynamenella-like<br />
sphaeromatids (e.g., Menzies and Glynn, 1968; Glynn, 1970). However, Dynamene<br />
curalii is <strong>one</strong> of the dominant eubranchiates <strong>as</strong>sociated with the intertidal<br />
coral reef on Heron Island, Queensland. On Heron Island another, <strong>as</strong> yet undescribed,<br />
eubranchiate sphaeromatid which resembles Dynamene is also abundant<br />
on the reef. Dynamene species have a biph<strong>as</strong>ic life history; juveniles feed on and<br />
inhabit intertidal algae and adults occupy cryptic habitats (e.g., crevices and<br />
empty barnacle tests) where they reproduce (Holdich, 1976). This change of habitat<br />
is no doubt correlated with the degenerate state of the ovigerous females when<br />
they are brooding. The species of Dynamenella and Paradella studied here occupy<br />
similar habitats to species of Dynamene on the co<strong>as</strong>ts of northwestern<br />
Europe, but the Dynamenella in particular live higher up the shore and also<br />
inhabit sand. N<strong>one</strong> of the species of Dynamenella and Paradella examined appear<br />
to have a biph<strong>as</strong>ic life history, <strong>as</strong> all stages of development are present in any<br />
<strong>one</strong> habitat. However, workers in other countries have found Dynamenella-likc<br />
sphaeromatids on intertidal algae. The females of the species examined do not<br />
become degenerate at the ovigerous moult and perhaps develop more than <strong>one</strong><br />
brood. Usually the sex ratio strongly favours females (see below). The food<br />
source appears to be from the algal/bacterial film on the surface of the rocks,<br />
from filamentous green and red algae, and in the c<strong>as</strong>e of sand dwellers from dead<br />
and dying peracarid crustaceans.<br />
As there is some overlap in the habitat preferences of the species under consideration,<br />
the species are examined together under each locality. However, /*chyromene<br />
polytyla is known from only a single record in southe<strong>as</strong>tern Queensland<br />
and details of its ecology are not yet known, except that it w<strong>as</strong> from wood<br />
on the shore.
HARMSON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 111<br />
Island Sites<br />
Heron Island.—Besides the coral reef and surrounding cay, are<strong>as</strong> of beach rock<br />
(formed from lithified coral sand) occur on the upper part of the south and north<br />
shores (Plate la). As described by Endean et al. (1956b) the beach rock is inhospitable<br />
with high surface temperatures and desiccating conditions at low tide.<br />
Only periwinkles such <strong>as</strong> Nodilittorina pyramidalis Quoy and Gaimard and Melaraphe<br />
spp., and the chiton, Acanthozostera gemmata (Blainville), appear able<br />
to cope with such conditions. Few workers have apparently bothered to examine<br />
crevices within the beach rock. Although these are out of the water for most of<br />
the day they are moist and contain a surprising variety of life. Most notable is<br />
Dynamenella ptychura. Many hundreds of individuals of all stages of the life<br />
cycle were found clustered in the crevices (Plate lb). In two populations 15 and<br />
24 adult males, 15 and 14 subadult males, 177 and 70 ovigerous females, 31 and<br />
20 nonovigerous females, and 74 and 240 juveniles were found. All individuals<br />
were bright green which w<strong>as</strong> perhaps because they were feeding on the film of<br />
green algae which w<strong>as</strong> growing in the outer illuminated part of the crevice. No<br />
other food appeared to be available in the crevices at low tide except for the<br />
perithecia of marine lichens. (Specimens of D. ptychura were kept alive for a<br />
number of months in an aquarium containing beach rock and its <strong>as</strong>sociated microflora.)<br />
When the beach rock occ<strong>as</strong>ionaUy became covered in water at high tide<br />
some individuals were seen to swim from the crevices and crawl on the surface<br />
of the submerged beach rock before entering another crevice. This activity could<br />
explain why <strong>one</strong> specimen w<strong>as</strong> found on algae lower down on the reef. Besides<br />
D. ptychura only <strong>one</strong> other sphaeromatid w<strong>as</strong> found in beach rock crevices and<br />
this appears to belong to an undescribed genus. Other occupants included terrestrial<br />
isopods and Ligia sp., periwinkles, lithophage barnacles, and an actinian.<br />
[No sphaeromatids were found under the chitons <strong>as</strong> w<strong>as</strong> reported by Glynn<br />
(1968a) from Puerto Rico.]<br />
Magnetic Island.—Although a wide variety of habitats were examined on this<br />
granitic island only Dynamenella liochroea and Paradella octaphymata were<br />
recorded. D. liochroea w<strong>as</strong> found in semipermanent wood in the upper z<strong>one</strong> of<br />
a moderately flat shore containing a mixture of rock and muddy sand. The wood<br />
appears to provide a refuge for many peracarid crustaceans; amongst the isopod<br />
species, Gnathia falcipenis Holdich and Harrison, Gnathia meticola Holdich and<br />
Harrison, Cirolana cranchii Leach var. australiensis Hale, Corallana nodosa<br />
Schioedte and Meinert, Sphaeromopsis serriguberna Holdich and Harrison, were<br />
found along with species of Limnoria Leach.<br />
Paradella octaphymata w<strong>as</strong> observed at Alma Bay on the mid to lower shore<br />
of a moderately exposed rock face sloping at 45° into the water (Plate Ila). As<br />
shown by Endean et al. (1956b) corals occur in patches on the rocky shores of<br />
Magnetic Island around the low water spring tide level and below. At Alma Bay<br />
brown seaweeds, Sarg<strong>as</strong>sum spp., occur in this z<strong>one</strong> with the corals; above this,<br />
and up to the high water neap tide level, barnacles, including Tetralicta rosea<br />
(Krauss), and rock oysters, Cr<strong>as</strong>sostrea am<strong>as</strong>a (Iredale), commonly occur. Between<br />
the high water neap and high water spring levels various barnacle and<br />
periwinkle species predominate. Observations were made in April on a spring<br />
tide <strong>as</strong> it flowed over the rock oyster z<strong>one</strong>. This z<strong>one</strong> also contained a mat of<br />
filamentous green and red algae with trapped silt. As the sw<strong>as</strong>h covered the rocks<br />
and then started to drain away numerous P. octaphymata and amphipods were<br />
seen moving over the surface of the algae and rocks. As soon <strong>as</strong> the water drained
112<br />
JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Plate la. Upper shore on Heron Is. showing the z<strong>one</strong> of beach rock above the reef flat and below<br />
the sand of the cay.<br />
Plate lb. The inside of a beach rock crevice on Heron Is. showing the <strong>large</strong> number ot Dynamenella<br />
ptychura present.
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 113<br />
Plate Ila. The littoral z<strong>one</strong> at Alma Bay, Magnetic Island. The arrow marks the z<strong>one</strong> of amphipods<br />
and isopods. a, z<strong>one</strong> of corals and brown algae; b, red and green filamentous algae, and rock oysters;<br />
c, rock oysters and barnacles; d, barnacles and periwinkles.<br />
Plate lib. The lower part of a wooden pier pile at low tide at Pallarenda, TownsviUe. Note the<br />
profusion of epizoonts, especially bryozoans.<br />
4d
114 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
away they ce<strong>as</strong>ed to move until the rock surface w<strong>as</strong> moistened again. When<br />
further observations were made in August, amphipods, but no isopods, were seen<br />
performing the same activity pattern. In addition to P. octaphymata a species of<br />
Dynoidella Pillai w<strong>as</strong> found in the same z<strong>one</strong> but mainly <strong>as</strong>sociated with barnacles.<br />
Lizard Island.—On this granitic island four sphaeromatid species considered here<br />
were found. As on Heron Island, Dynamenella ptychura lived in crevices of<br />
beach rock, but D. liochroea and Zuzara sp. were also found here. D. liochroea,<br />
and more commonly, D. trachydermata were found in granitic rock crevices of<br />
the upper shore. D. liochroea w<strong>as</strong> also found in sand around the lower levels of<br />
beach rock along with Sphaeromopsis serriguberna. Paradella octaphymata occupied<br />
a similar z<strong>one</strong> to that on Magnetic Island, i.e., amongst mid to lower shore<br />
rock oysters, barnacles, and filamentous algae. As on Magnetic Island Dynoidella<br />
sp. w<strong>as</strong> also recorded from this habitat. At West Point, where slabs of granite<br />
slope into the sea, P. octaphymata occupied the lower half of the rock oyster<br />
and barnacle z<strong>one</strong> down to low water; D. trachydermata occurred in the upper<br />
half of the rock oyster z<strong>one</strong> and extended to the crevices of the upper shore<br />
where D. liochroea also occurred. Although only D. liochroea w<strong>as</strong> found inhabiting<br />
sand both D. trachydermata and D. ptychura have been recorded from that<br />
habitat on Hinchinbrook Island (situated between Townsville and Cairns).<br />
Mainland Sites (Working Northwards)<br />
Townsville Harbour.—Large vessels frequently enter and leave this international<br />
harbour. It is very sheltered in the inner parts and all the concrete, metal, and<br />
wooden man-made structures are covered by algae and sessile animals. A number<br />
of isopod species were collected from the inner harbour but, of the sphaeromatids<br />
considered here, only Paradella dianae w<strong>as</strong> found. It occurred with Gnathia<br />
biorbis Holdich and Harrison, Gnathia meticola, and Sphaeroma walkeri Stebbing<br />
among barnacles attached to pier piles near low water. S. walkeri w<strong>as</strong> a<br />
common inhabitant of the harbour especially in summer. P. dianae also occurs<br />
further south than Townsville and h<strong>as</strong> been recorded from Goat Island, and North<br />
Stradbroke Island near Brisbane, from under intertidal rocks.<br />
Kissing Point.—This promontory of red granite is exposed to much wave action<br />
and a wide variety of microhabitats are present. Crevices on the upper shore<br />
often contained both Dynamenella trachydermata and D. liochroea. Both were<br />
also found under st<strong>one</strong>s on sand, and, in more sheltered places, in sand near<br />
midshore rocks. Adults were more common in crevices than sand. Individuals in<br />
sand tended to be paler than those in crevices. Those from crevices were mostly<br />
green. The colour variation probably results from diet, <strong>as</strong> algal food can lead to<br />
a change in subcuticular pigmentation in sphaeromatids (Holdich, 1969).<br />
Pallarenda (Just North ofTownsville).—Collections were made from the piles of<br />
a decaying wooden pier and from rocky outcrops in the mid and upper shore (no<br />
rocks were present on the lower shore). Both habitats contained Paradella dianae,<br />
Dynamenella liochroea, and D. trachydermata. The lower parts of the pier<br />
piles were thickly covered by epizoonts—mainly bryozoans, sponges, barnacles,<br />
and hydroids (Plate lib). The three isopods were found mainly under this growth<br />
amongst the barnacles. The rocks of the mid and upper shore were covered with<br />
rock oysters and pink barnacles (Tetralicta rosea) and all three isopods were<br />
found amongst them. Sphaeroma walkeri w<strong>as</strong> also collected on the mid and upper<br />
shore.
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 115<br />
Kurrimine and District.—Most of the shores sampled in this area were sandy<br />
with either outcrops of soft red rock, or boulders scattered on sand, on the mid<br />
and upper shore. Dynamenella trachydermata w<strong>as</strong> found on both mid and upper<br />
shores in crevices and amongst filamentous red algae <strong>as</strong>sociated with the red<br />
rock. D. liochroea w<strong>as</strong> found amongst the red algae on the midshore only, but<br />
it w<strong>as</strong> also relatively common under st<strong>one</strong>s on fine sand or in the sand itself on<br />
the midshore, especially where there w<strong>as</strong> freshwater running across the shore or<br />
an estuary nearby. Sphaeromopsis serriguberna, however, w<strong>as</strong> much more abundant<br />
than D. liochroea under the st<strong>one</strong>s (Holdich and Harrison, 1981b).<br />
Cairns and District.—At the headland of Palm Beach only Dynamenella trachydermata<br />
w<strong>as</strong> found in the rock oyster z<strong>one</strong> and above. It occurred in crevices<br />
containing lithophage barnacles and green algae just like D. ptychura on Heron<br />
Island and Lizard Island. On the midshore at Yorkey's Knob both D. liochroea<br />
and D. trachydermata were <strong>as</strong>sociated with Teredo-bored wood. D. liochroea<br />
w<strong>as</strong> also found around midshore rocks in fine sand, and D. trachydermata w<strong>as</strong><br />
found in crevices just below the rock oyster z<strong>one</strong>.<br />
ACKNOWLEDGEMENTS<br />
Thanks are due to the Nuffield Foundation for a Fellowship in Tropical Marine Biology to D. M.<br />
Holdich, the Natural Environment Research Council for research grants, and to Professor C. Burdon-<br />
J<strong>one</strong>s (James Cook University, TownsviUe) and Professor P. N. R. Usherwood (Nottingham University),<br />
for providing laboratory facilities. The authors would also like to thank Dr. A. H. Sommerstein<br />
(Department of Cl<strong>as</strong>sics, Nottingham University) for reviewing the construction of the new<br />
generic and specific names, the members of staff in the museums menti<strong>one</strong>d in the Methods section<br />
for providing specimens, and N. L. Bruce for donating specimens from several localities.<br />
LITERATURE CITED<br />
Audouin, V. 1826. Explication des Planches: Crustacea.—In: Savigny's Description de I'Egypte.<br />
Histoire Naturelle 1: 77-98.<br />
Baker, W. H. 1908. Notes on some species of the isopod family Sphaeromidae, from the South<br />
Australian co<strong>as</strong>t.—Transactions of the Royal Society of South Australia 32: 105-119.<br />
. 1926. Species of the isopod family Sphaeromidae, from the e<strong>as</strong>tern, southern, and western<br />
co<strong>as</strong>ts of Australia.—Transactions of the Royal Society of South Australia 50: 247-279.<br />
1929. Australian species of the isopod familiy Sphaeromidae (continued).—Transactions of<br />
the Royal Society of South Australia 52: 49-61.<br />
Barnard, K. H. 1914. Contributions to the crustacean fauna of South Africa. 3—Additions to the<br />
marine Isopoda with notes on some previously incompletely known species.—Annals of the<br />
South African Museum 10: 325a-358a, 359-442.<br />
. 1920. Contributions to the crustacean fauna of South Africa. 6—Further additions to the list<br />
of marine Isopoda.—Annals of the South African Museum 17: 319-438.<br />
. 1940. Contributions to the crustacean fauna of South Africa. 12—Further additions to the<br />
Tanaidacea, Isopoda, and Amphipoda together with keys for the identification of the hitherto<br />
recorded marine and freshwater species.—Annals of the South African Museum 32: 120-543.<br />
. 1954. New records and new species of Crustacea from South Africa.—Annals du Musee<br />
Royale du Congo Beige (N.S. 4to), Zoologie 1: 120-131.<br />
. 1959. New and little known South African marine isopods.—Annals and Magazine of Natural<br />
History 13: 715-720.<br />
Blackwelder, R. E. 1967. Taxonomy, a text and reference book. John Wiley and Sons, Inc., New<br />
York.<br />
Bo<strong>one</strong>, P. L. 1923. New marine tanaid and isopod Crustacea from California.—Proceedings of the<br />
Biological Society of W<strong>as</strong>hington 36: 147-156.<br />
Bruce, N. L. 1980. The systematics of some Japanese marine isopods (Fam. Sphaeromatidae) of the<br />
genera Dynoides Barnard, 1914 and Cymodocella Pfeffer, 1887, with descriptions of two new<br />
species.—Crustaceana 38: 199-211.<br />
Chilton, C. 1883. On two new isopods.—Transactions and Proceedings of the New Zealand Institute<br />
15: 188-190.
116 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
1892. Notes on some New Zealand Amphipoda and Isopoda.—Transactions and Proceedings<br />
of the New Zealand Institute 24: 258-269.<br />
Endean, R., R. Kenny, and W. Stephenson. 1956a. The ecology and distribution of intertidal organisms<br />
on the rocky shores of the Queensland mainland.—Australian Journal of Marine and Freshwater<br />
Research 7: 88-146.<br />
, W. Stephenson, and R. Kenny. 1956b. The ecology and distribution of intertidal organisms<br />
on certain islands off the Queensland co<strong>as</strong>t.—Australian Journal of Marine and Freshwater Research<br />
7: 317-342.<br />
Glynn, P. W. 1968a. Ecological studies on the <strong>as</strong>sociations of chitons in Puerto Rico, with special<br />
reference to sphaeromatid isopods.—Bulletin of Marine Science 18: 572-626.<br />
. 1968b. A new genus and two new species of sphaeromatid isopods from the high intertidal<br />
z<strong>one</strong> at Naos Island, Panama.—Proceedings of the Biological Society of W<strong>as</strong>hington 81: 587-603.<br />
1970. A systematic study of the Sphaeromatidae (Crustacea: Isopoda) of Isla Margarita,<br />
Venezuela, with descriptions of three new species.—Memori<strong>as</strong>, Sociedad de Cienci<strong>as</strong> Naturales<br />
La Salle 30: 5^8.<br />
Hansen, H. J. 1905. On the propagation, structure and cl<strong>as</strong>sification of the Family Sphaeromidae.—<br />
Quarterly Journal of Microscopical Science 49; 69-135.<br />
Harrison, K. 1981. Preliminary description of a new species of Ischywmene (Isopoda: Sphaeromatidae)<br />
from the e<strong>as</strong>tern Mediterranean.—Crustaceana 41: 312-314.<br />
Hatch, M. H. 1947. The Chelifera and Isopoda of W<strong>as</strong>hington and adjacent regions.—University of<br />
W<strong>as</strong>hington Publications in Biology 10: 155-274.<br />
Heller, C. 1865. Crustaceen.—In: Reise der Osterreichischen Fregatte Novara um die Erde in den<br />
Jahren 1857, 1858, 1859, Zoologischer Theil II Band 3: 1-280. Vienna.<br />
Holdich, D. M. 1969. Polychromatism in the genus Dynamene (Crustacea: Isopoda).—Publicazi<strong>one</strong><br />
della Stazi<strong>one</strong> Zoologica di Napoli 37: 18-27.<br />
. 1970. The distribution and habitat preferences of the Afro-European species of Dynamene<br />
(Crustacea: Isopoda).—Journal of Natural History 4: 419-^38.<br />
. 1976. A comparison of the ecology and life cycles of two species of littoral isopod.—Journal<br />
of Experimental Marine Biology and Ecology 24: 133-149.<br />
, and K. Harrison. 1980a. Morphological variation in the Serolis minuta-group (Isopoda: Serolidae)<br />
from Australian waters.—Zoological Journal of the Linnean Society, London 68: 373-386.<br />
, and . 1980b. The crustacean isopod genus Gnathia Leach from Queensland waters<br />
with descriptions of nine new species.—Australian Journal of Marine and Freshwater Research<br />
31: 215-240.<br />
, and . 1980c. The isopod genus Dynamene from Australian waters, with the description<br />
of a new species from coral reefs.—Memoirs of the Queensland Museum 20: 163-170.<br />
, and . 1981a. Platybranch sphaeromatids (Crustacea: Isopoda) from the Australian<br />
region with description of a new genus.—Records of the Australian Museum 33: 617-643.<br />
, and . 1981b. The sphaeromatid isopod genus Sphaeromopsis Holdich & J<strong>one</strong>s<br />
in African, Australian and South American waters.—Crustaceana 41; 286-300.<br />
and N. L. Bruce. 1981. Cirolanid isopod crustaceans from the Townsville region<br />
of Queensland, Australia, with descriptions of six new species.—Journal of Natural History<br />
15; 555-605.<br />
Hurley, D. E. and K. P. Jansen. 1971. Preliminary diagnoses of four new littoral species of marine<br />
Crustacea Isopoda (family Sphaeromatidae).—New Zealand Journal of Marine and Freshwater<br />
Research 4: 472-473.<br />
, and . 1977. The marine fauna of New Zealand; Family Sphaeromatidae (Crustacea;<br />
Isopoda: Flabellifera).—Memoirs of the New Zealand Oceanographic Institute 63; 1-95.<br />
International Commission on Zoological Nomenclature. 1961. International Code of Zoological Nomenclature<br />
adopted by the XV International Congress of Zoology, London, July 1958. International<br />
Trust for Zoological Nomenclature, London.<br />
Iverson, E. W. 1974. Range extensions for some California marine isopod crustaceans.—Bulletin of<br />
Ihe Southern California Academy of Sciences 73: 164-169.<br />
Kensley, B. 1971. A new eubranchiate sphaeromatid isopod from Angola.—Cimbeb<strong>as</strong>ia (Ser. A) 2:<br />
37^2.<br />
. 1976. Isopodan and Tanaidacean Crustacea from St. Paul and Amsterdam Islands, Southern<br />
Indian Ocean.—Annals of the South African Museum 69; 261-323.<br />
Krauss, F. 1843. Die siidafrikanischen Crustaceen. E. Schweizerbart'sche Verlagsbuchhandlung,<br />
Stuttgart.<br />
Kussakin, O. G. 1962. Novyi vid literal nogo raka (Isopoda, Sphaeromidae) iz dal'nevostochnykh<br />
morei SSSR.—Issledovanie Dal'nevostochnykh morei SSSR (Akademiya Nauk. SSSR) 8:<br />
238-242 (In Russian).<br />
Leach, W. E. 1818. Cymothoadees.—In: Dictionnaire des Sciences Naturelles 12. G. F. Cuvier (Ed).<br />
Paris.<br />
Menzies, R. J. 1962a. The marine isopod fauna of Bahia de San Quintin, Baja California, Mexico.—<br />
Pacific Naturalist 3: 337-348.
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 117<br />
—. 1962b. The zoogeography, ecology and systematics of the Chilean marine isopods. Report<br />
of the University of Lund Chile Expedition 1948-49 (42).—Acta Universitatis Lundensis 2, 57:<br />
1-162.<br />
and P. W. Glynn. 1968. The common marine isopod Crustacea of Puerto Rico. A handbook<br />
for marine biologists.—Studies on the Fauna of Curajao and Other Caribbean Islands 27: 1-133.<br />
Miers, E. J. 1875. Descriptions of new species of Crustacea collected at Kerguelen's Island by the<br />
Rev. A. E. Eaton.—Annals and Magazine of Natural History 16: 73-76.<br />
Milne Edwards, H. 1840. Histoire naturelle des Crustaces .... Librairie Encyclopedique de Roret,<br />
Paris.<br />
Monod, T. 1933. Mission Robert Ph. Dollfus en Egypt: Tanaidacea et Isopoda.—Memoires, Institut<br />
d'Egypte21: 161-264.<br />
. 1934. Isopodes marins des campagnes du "de Lanessan."—Note. Institut Oceanographique<br />
de ITndochine 23: 1-22.<br />
Moore, H. F. 1901. Reports on Porto Rican Isopoda.—Bulletin of the United States Fish Commission<br />
20: 161-176.<br />
Nishimura, S. 1969. A new CymodoceWa from Japan (Isopoda: Sphaeromatidae).—Publications of the<br />
Seto Marine Biological Laboratory 16: 335-344.<br />
Nobili, G. 1906. Diagnoses preliminaires des crustaces, Decapodes et Isopodes nouveaux recueillis<br />
par M. le Dr G. Seurat aux lies Touamotou.—Bulletin du Museum d'Histoire Naturelle 12:<br />
256-270.<br />
. 1907. Ricerche sui Crostacei de la Polinesia, Decapodi, Stomatopodi, Anisopodi e Isopodi.—<br />
Memorie della (Reale) Accademia delle Scienze di Torino, Ser. 2, 57: 351^30.<br />
Pfeffer, G. 1887. Die Krebse von Siid-Georgien nach der Ausbeute der Deutschen Station<br />
1882-1883.—Jahrbuch der Hamburgischen WissenschaftUchen Anstalten 4: 43-150.<br />
Pillai, N. K. 1954. A preliminary note on the Tanaidacea and Isopoda of Travancore.—Bulletin of<br />
the Research Institute of the University of Travancore 3: 1-23.<br />
Fires, A. M. S. 1980a. Sergiella angra, a new genus and species of Sphaeromatidae from Brazil.—<br />
Crustaceana 38: 212-218.<br />
. 1980b. New record of Sphaeromatidae (Isopoda) from the Brazilian southern co<strong>as</strong>t: Dynamenella<br />
dianae (Menzies, 1962).—Crustaceana 39: 133-140.<br />
Racovitza, E. G. 1908. Ischyromene lacazei n.g., n. sp. Isopode mediterraneen de la famille des<br />
spheromides (Note preliminaire).—Archives de Zoologie Experimentale et Generale 9 (Notes et<br />
Revue): LX-LXIV.<br />
Richardson, H. 1899. Key to the isopods of the Pacific co<strong>as</strong>t of North America, with descriptions of<br />
twenty-two new species.—Proceedings of the United States National Museum 21: 815-869.<br />
. 1901. Key to the isopods of the Atlantic co<strong>as</strong>t of North America, with descriptions of new<br />
and little known species.—Proceedings of the United States National Museum 23: 493-579.<br />
1906. Descriptions of new isopod crustaceans of the family Sphaeromidae.—Proceedings of<br />
the United States National Museum 31: 1-22.<br />
Schultz, G. A. 1969. How to know the marine isopod crustaceans. W. C. Brown, Dubuque, Iowa.<br />
Silva, J. de Loyola e. 1960. Sphaeromatidae do litoral Br<strong>as</strong>ileiro (Isopoda-Crustacea).—Boletim da<br />
Universidade do Parana 4: 1-182.<br />
. 1965. Especie nova de Cymodocella Pfeffer, 1887 (Sphaeromatidae- Isopoda) do litoral Br<strong>as</strong>ileiro.—Boletim<br />
do Instituto de Defesa do Patrimonio Natural 7: 1-20.<br />
Simes, E. G. 1981. On the systematic position of Dynamenella eatoni (Miers, 1875) and redescription<br />
of the species.—Crustaceana 40: 160-172.<br />
Sivertsen, E., and L. B. Holthuis. 1980. The marine isopod Crustacea of the Tristan da Cunha<br />
Archipelago.—Gunneria 35: 1-128.<br />
Stebbing, T. R. R. 1875. On some new exotic sessile-eyed crustaceans.—Annals and Magazine of<br />
Natural History 15: 184-188.<br />
Stephenson, W., R. Endean, and I. Bennett. 1958. An ecological survey of the marine fauna of Low<br />
Isles, Queensland.—Australian Journal of Marine and Freshwater Research 9: 261-318.<br />
Thomson, G. M. 1879. New Zealand Crustacea, with descriptions of new species.—Transactions and<br />
Proceedings of the New Zealand Institute 11: 230-248.<br />
Vanhoffen, E. 1914. Die Isopoden der Deutschen Siidpolar—Expedition 1901-1903.—Deutsche Siidpolar—Expedition<br />
1901-1903, 15, Zoologie 7: 449-598.<br />
APPENDIX<br />
Additional Material Examined<br />
Ischyromene lacazei Racovitza, 1908—Banyuls-sur-mer, France. Pres. G. Racovitza. 3 specimens<br />
(Syntypes. B.M.N.H.: 1910.1.10.21-23).—Lagune du Brusc, France. On Cystoseira stricta. Coll.<br />
M. L. Roman, 13.ii. 1978. 4 adult males, 14 subadult males, 2 ovigerous females, 2 nonovigerous<br />
females, 25 juveniles. (Fig. lo-s).
118 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 2, NO. 1, 1982<br />
Ischyromene bicarinata Harrison, 1981—Palmachim, north of Ashdod, Israel. On Hypnea sp.<br />
Intertidal. Coll. L. Fishelson, 23.viii.1977. 1 adult male (Holotype. B.M.N.H.: 1980.222.1), 1<br />
subadult male, 1 nonovigerous female, 3 juveniles (Paratypes. B.M.N.H.: 1980.223.5).<br />
Dynamenopsis obtusa Baker, 1908—Denial Bay, South Australia. Coll. Verco and Torr. 1 adult<br />
male (Holotype. S.Aust.M.: C.359) (Fig. 8i, j)—North Point, Green Island, Rottnest Island,<br />
Western Australia. Amongst algae and rock on limest<strong>one</strong> reef platform. Depth 0.3 m. Coll. L.<br />
M. Joll, 08-10.vi.l972. 1 adult male (W.A.M.: 56-80) (Fig. 8a-h).<br />
Dynamenopsis dumerilii (Audouin, 1826)—Bir Suer, Red Sea. On weed. Coll. L. Fishelson,<br />
06.xi. 1976. 1 subadult male.<br />
Dynamenopsisplatura (NobiH, 1906)—I. Gambler. Coll. Dr. Seurat, 1903. 8 specimens. (Syntypes.<br />
M.d'H.N.).<br />
Dynamenopsis varicolor Hurley and Jansen, 1971—New Zealand. 1 adult male. (Holotype. CM.).<br />
Dynamenopsis sp.—Natural Jetty, Rottnest Island, Western AustraHa. Coll. R. W. George,<br />
19.ii.l959. 1 nonovigerous female. (W.A.M.: 57-80) (Fig. 8k-o).<br />
Cymodocella tubicauda Pfeffer, 1887—Hallett B., Antarctica. 7 specimens. (N.Z.O.I.: Z.1795).<br />
Cymodocella cancellata Barnard, 1920—South Africa. Pres. S.Afr.M. 1 specimen. (Holotype.<br />
B.M.N.H.: 1937.11.10.82).—34°12.9'S, 18°49.I'E. Coll. University of Cape Town. 1 male<br />
(S.Afr.M.: A. 14954).<br />
Cymodocella capra Hurley and Jansen, 1977—New Zealand. 1 male (Paratype. N.Z.O.I.: E.977).<br />
Cymodocella diateichos Barnard, 1959—Saldanha Bay, South Africa. Intertidal. Coll. University<br />
of Cape Town, 22.ix.1957. 1 specimen (Holotype. S.Afr.M.: A.13663).<br />
Cymodocella egregia (Chilton, 1892)—Makorori Beach, New Zealand. Intertidal. Coll. N.Z.O.I.,<br />
20.ii.l968. 7 specimens (N.Z.O.I.: E.982).<br />
Cymodocella eutylos Barnard, 1954—South Africa. 1 male, 1 ovigerous female. (Syntypes.<br />
S.Afr.M.: A. 10409).<br />
Cymodocella foveolata Menzies, 1962b—Estacion de Biologia Marina, Montemar, Chile<br />
(32°57'24"S, 71°33'25"W). Intertidal. Coll. Lund University Chile Expedition 1948-49, 15.x. 1948.<br />
1 specimen (Holotype. N.R.: ISOPODA 9453);—(Details <strong>as</strong> above) Coll. 15.vi.l949. 3 specimens<br />
(Paratypes. N.R.: ISOPODA 9454).<br />
Cymodocella magna Barnard, 1954—South Africa. 3 specimens (Syntypes. S.Afr.M.: A. 10408);—<br />
South Africa. Coll. R. W. Rand. 31 specimens (S.Afr.M.: A. 10415).<br />
Cymodocella pustulata Barnard, 1914—Mouille Pt., South Africa. Coll. K. H. Barnard, 15.iv.1913.<br />
125 specimens (Syntypes. S.Afr.M.: A.2607);—South Africa. Coll. K. H. Barnard. 10 specimens<br />
(Syntypes. B.M.N.H.: 1927.5.31.99-108).<br />
Cymodocella sublevis Barnard, 1914—Mouille Pt., South Africa. Coll. K. H. Barnard, 29.xi.1913.<br />
18 specimens (Syntypes. S.Afr.M.: A.2623);—Sea Point, near Cape Town, South Africa. Coll.<br />
K. H. Barnard. 12 specimens (Syntypes. B.M.N.H.: 1927.5.31.109-118).<br />
Dynamenella perforata (Moore, 1901)—Ensenada Honda, Culebra, Puerto Rico. Coll. U.S. Fish<br />
Commission (Steamer "Fish Hawk"), ll.ii.l899. 3 adult males, 1 subadult male, 1 ovigerous<br />
female (no brood), 1 nonovigerous female (Types. U.S.N.M.: 32649);—Puerto Rico. Coll. P.<br />
Glynn. 1 male, 2 ovigerous females, 9 nonovigerous females (U.S.N.M.: 259321).<br />
Dynamenella acuticauda Menzies, 1962b—Estrecho de Magellanes, south of Punta Aren<strong>as</strong><br />
(53°irS, 70°55'W). Intertidal. Coll. Lund University Chile Expedition 1948^9, 03.V.1949. 1<br />
nonovigerous female (Holotype. N.R.: ISOPODA 9450).<br />
Dynamenella angulata (Richardson, 1901)—Florida, U.S.A. Coll. H. Hemphill. 11 subadult males<br />
(Types. U.S.N.M.: 23906).<br />
Dynamenella australis Richardson, 1906—Cape Town, South Africa. Coll. K. H. Barnard. 2 adult<br />
males. (B.M.N.H.: 1927.5.31.127-128).<br />
Dynamenella australoides Barnard, 1940—St. James, Cape Peninsula, South Africa. Pres. S.Afr.M.<br />
2 ovigerous females (Syntypes. B.M.N.H.: 1938.4.27.6-7).<br />
Dynamenella bakeri (Menzies, 1962b)—Iquique, Chile. (20°12'30"S, 70''10'19"W). The harbour. Tidal<br />
belt. Coll. Lund University Chile Expedition 1948^9 (station M.133), 02.vii.l949. 3 adult<br />
males, 2 ovigerous females, 1 juvenile (Types, N.R.: ISOPODA 9468).<br />
Dynamenella benedicti Richardson, 1899—Monterey Bay, California, U.S.A. At the surface. Coll.<br />
H. Heath. 1 male, 1 ovigerous female (Types. U.S.N.M.: 22570).<br />
Dynamenella bicolor Barnard, 1914—Sea Point, Near Cape Town, South Africa. Coll. K. H. Barnard.<br />
1 subadult male (Syntype. B.M.N.H.: 1927.5.13.129).<br />
Dynamenella brunnea Vanhoffen, 1914—St. Paul. Coll. Deutsche Siidpolar Expedition 1901-03. 1<br />
ovigerous female (Syntype. B.M.N.H.: 1924.7.19.34).<br />
Dynamenella codii Nobili, 1906—Makapu. Coll. Dr. Seurat, 1905. 8 specimens. (Syntypes.<br />
M.d'H.N.).<br />
Dynamenella condita Hurley and Jansen. 1977—New Zealand. 1 adult male. (Holotype. CM.).<br />
Dynamenella cordiforaminalis (Chilton, 1883)—Bethell's Beach, New Zealand. Intertidal. Coll.<br />
N.Z.O.I., 21.x. 1968. 40 specimens (N.Z.O.I.: E.949).
HARRISON AND HOLDICH: NEW SPHAEROMATID ISOPODS FROM AUSTRALIA 119<br />
Dynamenella dioxus Barnard, 1914—Cape Town, South Africa. Coll. K. H. Barnard. 1 male (Syntype.<br />
B.M.N.H.: 1927.5.31.119).<br />
Dynamenella eatoni (Miers, 1875)—Swain's Bay, Kerguelen. Coll. A. E. Eaton. 1 female (Lectotype.<br />
B.M.N.H.: 1979.219.1), 2 specimens (Paralectotypes. B.M.N.H.: 1979.220.2) (Lectotype<br />
chosen by E. Gomez Simes);—Cumberland Bay, South Georgia. Coll. Barrett Hamilton Expedition.<br />
2 subadult males (B.M.N.H.: 1921.12.15.30-31);—Puerto Deseado, Peninsula Foca, Santa<br />
Cruz. Pres. E. Gomez Simes, 08.vi.l976. 6 specimens (B.M.N.H.: 1979.335.6).<br />
Dynamenella fraudatrix Kussakin, 1962—Petrov Island, Japanese Sea. Intertidal. Coll. ?.ix.l934.<br />
Pres. Acad. Sci. Leningrad (As Dynamene glabra. Id. G. Gurjanova). Specimens including males<br />
and ovigerous females. (B.M.N.H.: 1936.3.18.19-26).<br />
Dynamenella hirsuta Hurley and Jansen, 1971—New Zealand. 1 adult male. (Holotype. CM.).<br />
Dynamenella huttoni (Thomson, 1878)—Maunganui Bluff, New Zealand. Intertidal. Coll. N.Z.O.I.,<br />
22.x. 1968. 13 specimens (N.Z.O.I.: E.952);—Lyttelton, New Zealand. Pres. C. Chilton. 5 specimens<br />
(B.M.N.H.: 1906.2.20.1-6);—Gough Island, South Atlantic. Coll. M. W. Holdgate. 10<br />
specimens (B.M.N.H.: 1958.4.15.120-129);—Cape Town, South Africa. Pres. K. H. Barnard. 1<br />
subadult male, 2 nonovigerous females (Syntypes of Dynamenella kraussi Barnard, 1914.<br />
B.M.N.H.: 1915.1.11.5-8);—West Co<strong>as</strong>t, Cape Peninsula, South Africa. Coll. K. H. Barnard. 9<br />
specimens (<strong>as</strong> Dynamenella kraussi) (B.M.N.H.: 1927.5.13.130-138).<br />
Dynamenella insulsa Hurley and Jansen, 1977—New Zealand. 1 adult male. (Holotype. CM.).<br />
Dynamenella macrocephala (Krauss, 1843)—Cape Town, South Africa. Coll. K. H. Barnard. 5<br />
specimens (B.M.N.H.: 1927.5.31.122-126).<br />
Dynamenella navicula Barnard, 1940—Port Elizabeth, South Africa. Pres. S.Afr.M. 1 ovigerous<br />
female, I nonovigerous female (Syntypes. B.M.N.H.: 1937.11.10.246-247).<br />
Dynamenella ovalis Barnard, 1914—St. James, False Bay, South Africa. Coll. K.H. Barnard. 1<br />
adult male, 1 juvenile (B.M.N.H.: 1927.5.31.120-121);-StiU Bay, South Africa. Coll. S.Afr.M.<br />
8 specimens (B.M.N.H.: 1937.11.10.162-167).<br />
Dynamenella parva Baker, 1929—Port Willunga Reef, South Australia. Coll. H. M. Hale. 11 specimens<br />
(Syntypes. S.Aust.M.: C.3723).<br />
Dynamenella rubida Baker, 1926—Marouba, New South Wales, Australia. 1 adult male (Type.<br />
A.M.: P.9487).<br />
Dynamenella scabricula (Heller, 1865)—Cape Town, South Africa. Pres. K. H. Barnard. 10 specimens<br />
(B.M.N.H.: 1916.11.20.43-52).<br />
Dynamenella taurus Barnard, 1940—B<strong>as</strong>t London, South Africa. Pres. S.Afr.M. 1 subadult male,<br />
1 nonovigerous female (Syntypes. B.M.N.H.: 1938.4.27.8-9).<br />
Dynamenella tuberculata Menzies, 1962b—Punta Corona, Canal Chacao, Chile (41°47'S,<br />
73°53'07"W). Intertidal. Coll. Lund University Chile Expedition 1948-49, 28.ii.1949. 1 adult male<br />
(Type. N.R.: ISOPODA 9445).<br />
Dynamenella sp.—Grande Anse, Mahe, Seychelles. From algae. Coll. A. Harris, 27.ix. 1976. 2 adult<br />
males, 4 immature specimens. (In general appearance these specimens resemble Dynamenella<br />
trachydermata, new species, the only major differences being that the endopod of pleopod 1 of<br />
the adult male does not extend beyond the exopod, and the pleotelsons of the immature specimens<br />
are smoother than those of D. trachydermata. These specimens have been desiccated and their<br />
condition is not sufficiently acceptable to allow their description <strong>as</strong> a new species) (Q.M.:<br />
W.7952).<br />
RECEIVED: 9 March 1981.<br />
ACCEPTED: 21 May 1981.<br />
Address: Department of Zoology, The University, Nottingham NG7 2RD, England.